
Micronutrients in Food Production: What Can We Learn from Natural Ecosystems?


{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Micronutrients and Hidden Hunger
1.2. Microbial Biofortification
2. Iron (Fe)
2.1. Fe Availability and Acquisition in Arable Soils
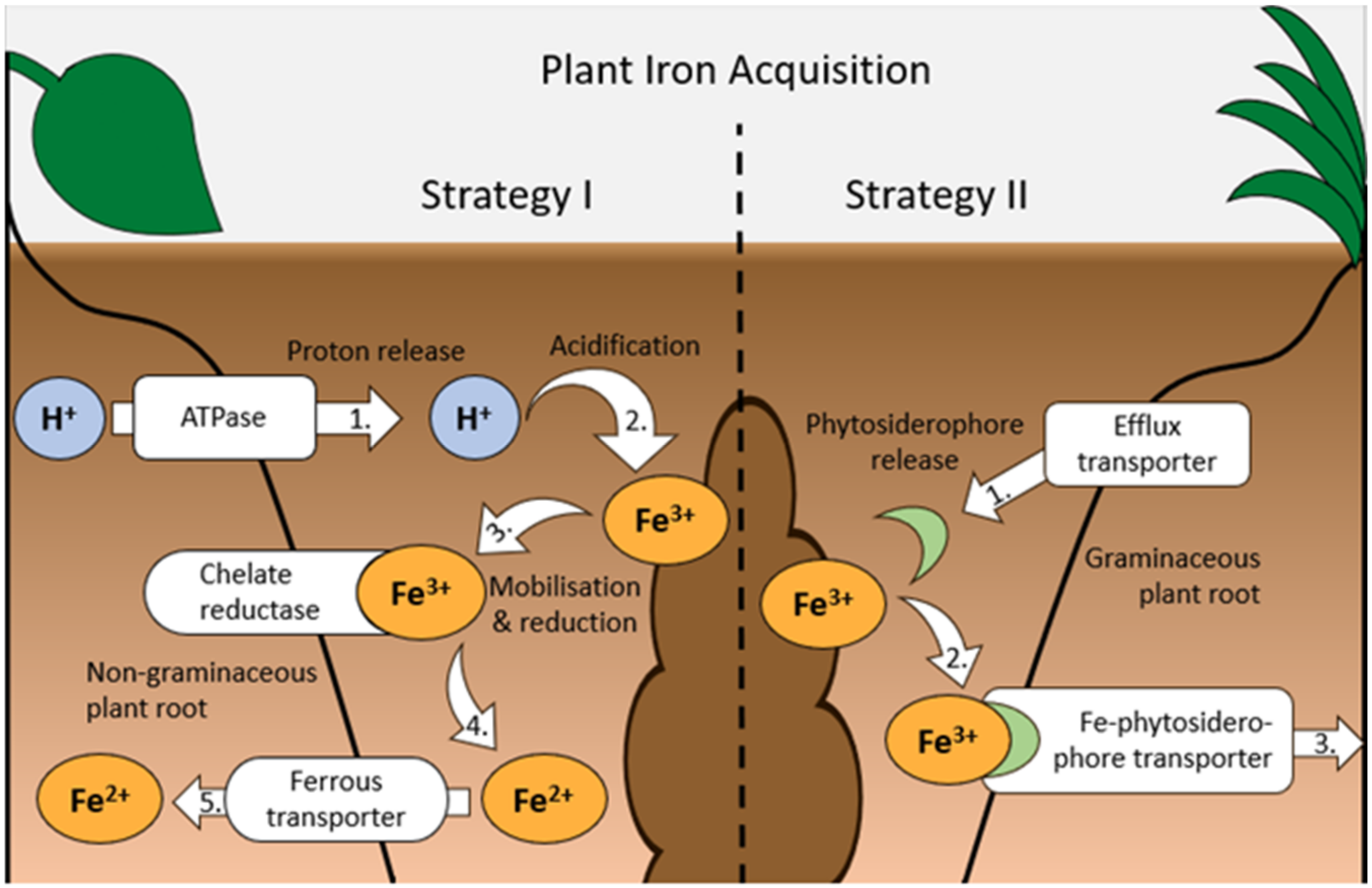
2.1.1. Fe Acquisition by Crops: Strategies and Efficiency
2.1.2. Fe Mobilization by Microbes
2.2. Fe in Natural Ecosystems
3. Zinc (Zn)
3.1. Zn Mobilization by Microbes
3.2. Lessons for Zn Biofortification
4. Boron (B) and Molybdenum (Mo)
4.1. Microbial B and Mo Requirements
4.2. Management of B and Mo in Arable Soils
4.3. B and Mo in Natural Ecosystems
5. Manganese (Mn)
5.1. Management of Mn in Arable Soils
5.2. Mn in Natural Ecosystems
6. Cobalt (Co)
Microbial Management of Co in Arable Soils
7. Copper (Cu)
Solutions for Cu Deficiency
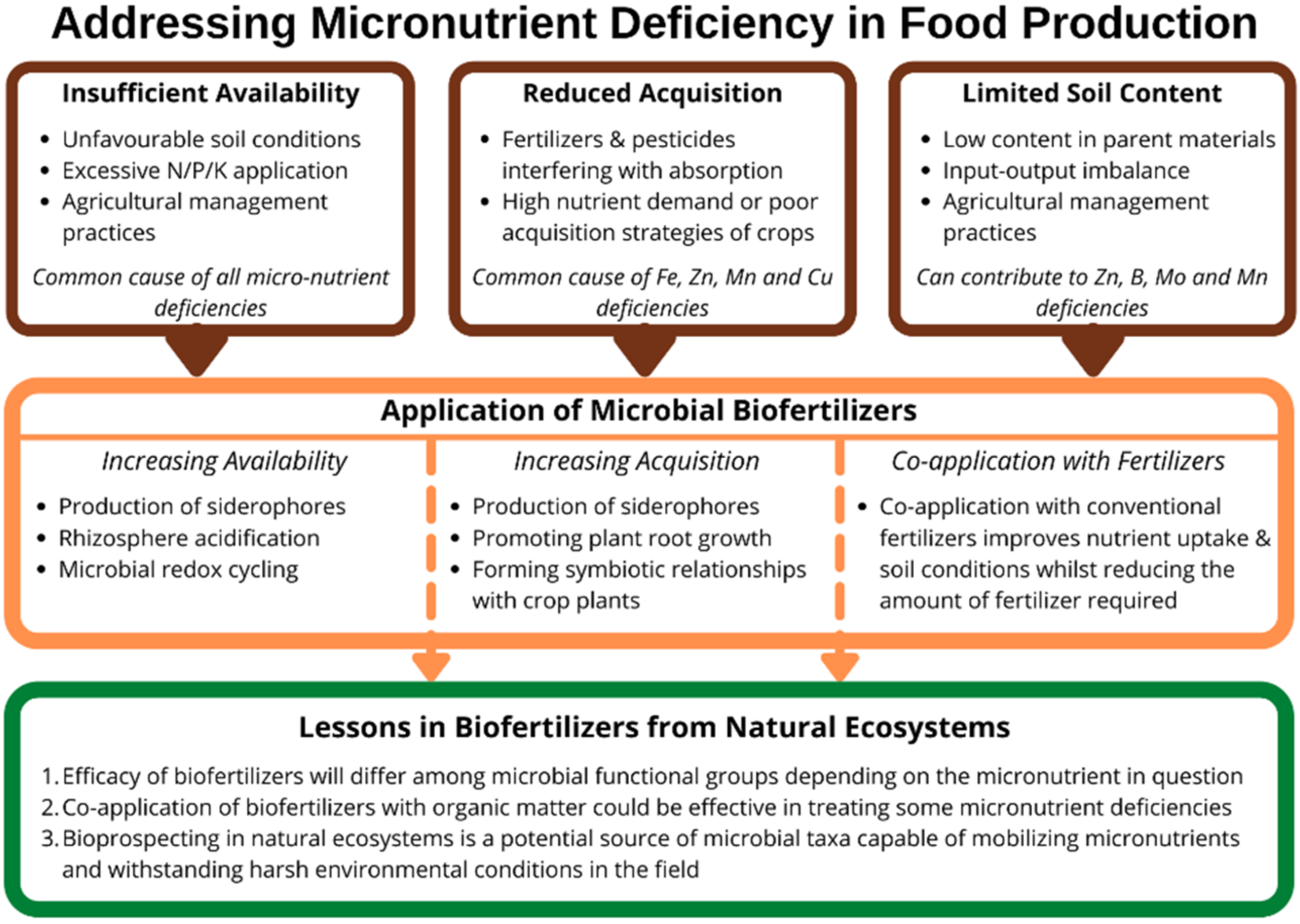
8. Conclusions
- (1)
- Microbial functional groups suitable for use as biofertilizers will differ depending on the micronutrient in question, and biotic interactions could undermine their effectiveness. Research to identify microorganisms for developing new micronutrient biofertilizers should not only characterize the mechanisms microbes employed to mobilize and acquire micronutrients, but also assess potential interactions among different microbial functional groups and ascertain the potential for plant–microbial nutrient competition.
- (2)
- The co-application of biofertilizers with inorganic fertilizers proves effective for treating deficiencies of Zn, B, and Mo, and reduces the amount of inorganic fertilizers needed. Substituting conventional inorganic fertilizers with nano-fertilizers is a growing area of interest, and research into their co-application with biofertilizers could reduce the associated risks of toxicity. Although the co-application of biofertilizers with organic fertilizers is under-researched, work on microbial interactions with easily degradable SOM in natural ecosystems suggests that the co-application of biofertilizers and organic fertilizers could boost microbial activity and micronutrient availability.
- (3)
- Finally, bioprospecting in natural ecosystems is a potential source of novel microbial taxa that are both capable of mobilizing numerous micronutrients and withstanding harsh environmental conditions. Biofertilizers, including such organisms, could mitigate the impacts of climate changes, as well as the negative impacts of conventional arable management practices on soil conditions, which can reduce microbial diversity and abundance. Therefore, the bioprospecting for microbial taxa suitable for reproduction and application as biofertilizers should focus on selecting species that are both highly effective and tolerant of unfavourable environmental conditions.
Author Contributions
Funding
Conflicts of Interest
References
- Watson, C.A.; Oborn, I.; Edwards, A.C.; Dahlin, A.S.; Eriksson, J.; Lindstrom, B.E.M.; Linse, L.; Owens, K.; Topp, C.F.E.; Walker, R.L. Using Soil and Plant Properties and Farm Management Practices to Improve the Micronutrient Composition of Food and Feed. J. Geochem. Explor. 2012, 121, 15–24. [Google Scholar] [CrossRef]
- Carvalho, S.M.P.; Vasconcelos, M.W. Producing More with Less: Strategies and Novel Technologies for Plant-Based Food Biofortification. Food Res. Int. 2013, 54, 961–971. [Google Scholar] [CrossRef]
- Ku, Y.S.; Rehman, H.M.; Lam, O.M. Possible Roles of Rhizospheric and Endophytic Microbes to Provide a Safe and Affordable Means of Crop Biofortification. Agronomy 2019, 9, 764. [Google Scholar] [CrossRef] [Green Version]
- Gomiero, T. Soil Degradation, Land Scarcity and Food Security: Reviewing a Complex Challenge. Sustainability 2016, 8, 281. [Google Scholar] [CrossRef] [Green Version]
- Power, A.G. Ecosystem Services and Agriculture: Trade-offs and Synergies. Philos. Trans. R. Soc. B 2010, 365, 2959–2971. [Google Scholar] [CrossRef]
- Scagliola, M.; Valentinuzzi, F.; Mimmo, T.; Cesco, S.; Crecchio, C.; Pii, Y. Bioinoculants as Promising Complement of Chemical Fertilizers for a More Sustainable Agricultural Practice. Front. Sustain. Food Syst. 2021, 4, 305. [Google Scholar] [CrossRef]
- Brady, N. The Nature and Properties of Soils, 8th ed.; Macmillan Publishing Co.: New York, NY, USA, 1974. [Google Scholar]
- Fageria, N.K.; Baligar, V.C.; Clark, R.B. Micronutrients in Crop Production. Adv. Agron. 2002, 77, 185–268. [Google Scholar]
- Alloway, B.J. Micronutrient Deficiencies in Global Crop Production; Springer Science & Business Media B.V.: Berlin, Germany, 2008. [Google Scholar]
- He, Z.L.; Yang, X.E.; Stoffella, P.J. Trace Elements in Agroecosystems and Impacts on the Environment. J. Trace Elem. Med. Biol. 2005, 19, 125–140. [Google Scholar] [CrossRef]
- Lowe, N.M. The global challenge of hidden hunger: Perspectives from the field. Proc. Nutr. Soc. 2021, 80, 283–289. [Google Scholar] [CrossRef]
- World Health Organisation (WHO) Guidelines on Food Fortification with Micronutrients. 2006. Available online: https://www.who.int/publications/i/item/9241594012 (accessed on 1 July 2021).
- World Health Organisation (WHO) Vitamin and Mineral Nutrition Information System (VMNIS). 2021. Available online: https://www.who.int/teams/nutrition-and-food-safety/databases/vitamin-and-mineral-nutrition-information-system (accessed on 1 July 2021).
- Ritchie, H.; Roser, M. Our World in Data: Micronutrient Deficiency. 2017. Available online: https://ourworldindata.org/micronutrient-deficiency (accessed on 1 July 2021).
- Fisher, G.E.J. Micronutrients and Animal Nutrition and the Link between the Application of Micronutrients to Crops and Animal Health. Turk. J. Agric. For. 2008, 32, 221–233. [Google Scholar]
- White, P.J.; Broadley, M.R. Biofortification of Crops with Seven Mineral Elements Often Lacking in Human Diets—Iron, Zinc, Copper, Calcium, Magnesium, Selenium and Iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- World Health Organisation (WHO) Biofortification of Staple Crops. 2019. Available online: https://www.who.int/elena/titles/biofortification/en/ (accessed on 1 July 2021).
- Abhilash, P.C.; Dubey, R.K.; Tripathi, V.; Gupta, V.K.; Singh, H.B. Plant Growth-Promoting Microorganisms for Environmental Sustainability. Trends Biotechnol. 2016, 34, 847–850. [Google Scholar] [CrossRef]
- Singh, D.; Prasanna, R. Potential of Microbes in the Biofortification of Zn and Fe in Dietary Food Grains. A Review. Agron. Sustain. Dev. 2020, 40, 15. [Google Scholar] [CrossRef]
- Behera, B.; Das, T.K.; Raj, R.; Ghosh, S.; Raza, B.; Sen, S. Microbial Consortia for Sustaining Productivity of Non-Legume Crops: Prospects and Challenges. Agric. Res. 2021, 10, 1–14. [Google Scholar] [CrossRef]
- Mitter, E.K.; Tosi, M.; Obregon, D.; Dunfield, K.E.; Germida, J.J. Rethinking Crop Nutrition in Times of Modern Microbiology: Innovative Biofertilizer Technologies. Front. Sustain. Food Syst. 2021, 5, 29. [Google Scholar] [CrossRef]
- Pii, Y.; Mimmo, T.; Tomasi, N.; Terzano, R.; Cesco, S.; Crecchio, C. Microbial Interactions in the Rhizosphere: Beneficial Influences of Plant Growth-Promoting Rhizobacteria on Nutrient Acquisition Process. A Review. Biol. Fertil. Soil. 2015, 51, 403–415. [Google Scholar] [CrossRef]
- Verma, S.; Kuila, A. Bioremediation of Heavy Metals by Microbial Process. Environ. Technol. Innov. 2019, 14, 100369. [Google Scholar] [CrossRef]
- Yadav, K.K.; Gupta, N.; Kumar, V.; Singh, J.K. Bioremediation of Heavy Metals from Contaminated Sites Using Potential Species: A Review. Indian J. Environ. Prot. 2017, 37, 65–84. [Google Scholar]
- Kabata-Pendias, A. Trace Elements in Soils and Plants; Taylor & Francis Group: Baton Rouge, LA, USA, 2010. [Google Scholar]
- Siqueira Freitas, D.; Wurr, R.; Reis dos Rodrigues, A.; De Barros Reis, F.; Soares de Carvalho, T.; Schulze, J.; Carbone Carneiro, M.A.; Guimaraes Guilherme, L.R. Hidden Nickel Deficiency? Nickel Fertilization Via Soil Improves Nitrogen Metabolism and Grain Yield in Soybean Genotypes. Front. Plant Sci. 2018, 9, 614. [Google Scholar] [CrossRef] [Green Version]
- Barman, M.; Datta, S.P.; Rattan, R.K.; Meena, M.C. Critical Limits of Deficiency of Nickel in Intensively Cultivated Alluvial Soils. J. Soil Sci. Plant Nutr. 2019, 20, 284–292. [Google Scholar] [CrossRef]
- Sayer, E.J. The Anatomy of an Excellent Review Paper. Funct. Ecol. 2018, 32, 2278–2281. [Google Scholar] [CrossRef] [Green Version]
- Lemanceau, P.; Bauer, P.; Kraemer, S.; Briat, J.-F. Iron Dynamics in the Rhizosphere as a Case Study for Analysing Interactions between Soils, Plants and Microbes. Plant Soil 2009, 321, 513–535. [Google Scholar] [CrossRef]
- Whitehead, D.C. Nutrient Elements in Grassland: Soil-Plant-Animal Relationships; CABI Publishing: Wallingford, UK, 2000. [Google Scholar]
- Meena, V.S.; Meena, S.K.; Verma, J.P.; Kumar, A.; Aeron, A.; Mishra, P.K.; Bisht, J.K.; Pattanayak, A.; Naveed, M.; Dotaniya, M.L. Plant Beneficial Rhizospheric Microorganism (PBRM) Strategies to Improve Nutrients Use Efficiency: A Review. Ecol. Eng. 2017, 107, 8–32. [Google Scholar] [CrossRef]
- Buni, A. Effects of Liming Acidic Soils on Improving Soil Properties and Yield of Haricot Bean. J. Environ. Anal. Toxicol. 2014, 4, 1–4. [Google Scholar]
- Mimmo, T.; Del Buono, D.; Terzano, R.; Tomasi, N.; Vigani, G.; Crecchio, C.; Pinton, R.; Zocchi, G.; Cesco, S. Rhizospheric Organic Compounds in the Soil-Microorganism-Plant System: Their Role in Iron Availability. Eur. J. Soil Sci. 2014, 65, 629–642. [Google Scholar] [CrossRef]
- Calabrese, S.; Barcellos, D.; Thompson, A.; Porporato, A. Theoretical Constrains on Fe Reduction Rates in Upland Soils as Function of Hydroclimatic Conditions. J. Geophys. Res. Biogeosciences 2020, 125, e2020JG005894. [Google Scholar] [CrossRef]
- Abbasi, M.K.; Zafar, M.; Sultan, T. Changes in Soil Properties and Microbial Indices across Various Management Sites in the Mountain Environments of Azad Jammu and Kashmir. Commun. Soil Sci. Plant Anal. 2010, 41, 768–782. [Google Scholar] [CrossRef]
- Deiss, L.; Kleina, G.B.; Moraes, A.; Franzluebbers, A.J.; Motta, A.C.V.; Dieckow, J.; Sandini, I.E.; Anghinoni, I.; Carvalho, P.C.F. Soil Chemical Properties Under No-Tillage as Affected by Agricultural Trophic Complexity. Eur. J. Soil Sci. 2019, 71, 1090–1105. [Google Scholar] [CrossRef]
- Bhople, B.S.; Sharma, S. Seasonal Variation of Rhizospheric Soil Properties under Different Land Use Systems at Lower Shivalik Foothills of Punjab, India. Agrofor. Syst. 2020, 94, 1959–1976. [Google Scholar]
- Schiere, J.B.; Ibrahim, M.N.M.; Van Keulen, H. The Role of Livestock for Sustainability in Mixed Farming: Criteria and Scenario Studies under Varying Resource Allocation. Agric. Ecosyst. Environ. 2002, 2, 139–153. [Google Scholar] [CrossRef]
- Sharma, S.; Dhaliwal, S.S. Effects of Sewage Sludge and Rice Straw Compost on Yield, Micronutrient Availability and Soil Quality under Rice-Wheat System. Commun. Soil Sci. Plant Anal. 2019, 50, 1943–1954. [Google Scholar] [CrossRef]
- Eker, S.; Ozturk, L.; Yazici, A.; Erenoglu, B.; Romheld, V.; Cakmak, I. Foliar-Applied Glyphosate Substantially Reduced Uptake and Transport of Iron and Manganese in Sunflower (Helianthus annus L.) Plants. J. Agric. Food Chem. 2006, 54, 10019–10025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborne, L.D.; Robson, A.D.; Bowran, D.G. The Impact of Chlorsulfuron and Diclofop-Methyl on Nutrient Uptake by Wheat. Aust. J. Agric. Res. 1993, 44, 1757–1766. [Google Scholar] [CrossRef]
- Kraemer, S.M.; Crowley, D.E.; Kretzschmar, R. Geochemical Aspects of Phytosiderophore-Promoted Iron Acquisition by Plants. Adv. Agron. 2006, 91, 1–46. [Google Scholar]
- Ferreira, M.J.; Silva, H.; Cunha, A. Siderophore-Producing Rhizobacteria as a Promising Tool for Empowering Plants to Cope with Iron Limitation in Saline Soils: A Review. Pedosphere 2019, 29, 409–420. [Google Scholar] [CrossRef]
- Curie, C.; Briat, J.F. Iron Transport and Signalling in Plants. Annu. Rev. Plant Biol. 2003, 54, 183–206. [Google Scholar] [CrossRef]
- Marschner, P.; Crowley, D.; Rengel, Z. Rhizosphere Interactions between Microorganisms and Plants Governs Iron and Phosphorus Acquisition along the Root Axis—Model and Research Methods. Soil Biol. Biochem. 2011, 43, 883–894. [Google Scholar] [CrossRef]
- Schroder, I.; Johnson, E.; De Vries, S. Microbial Ferric Iron Reductases. FEMS Microbiol. Rev. 2003, 27, 427–447. [Google Scholar] [CrossRef] [Green Version]
- Colombo, C.; Palumbo, G.; He, J.-Z.; Pinton, R.; Cesco, S. Review on Iron Availability in Soil: Interaction of Fe Minerals, Plants, and Microbes. J. Soils Sedim. 2014, 14, 538–548. [Google Scholar] [CrossRef]
- Roriz, M.; Carvalho, S.M.P.; Castro, P.M.L.; Vasconcelos, M.W. Legume Biofortification and the Role of Plant Growth-Promoting Bacteria in a Sustainable Agricultural Era. Agronomy 2020, 10, 435. [Google Scholar] [CrossRef] [Green Version]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial Siderophores and Their Potential Applications: A Review. Environ. Sci. Pollut. Res. 2016, 23, 3984–3999. [Google Scholar] [CrossRef]
- Vaid, S.K.; Kumar, A.; Sharma, A.; Srivastava, P.C.; Shukla, A.K. Role of Some Plant Growth Promotory Bacteria in Enhanced Fe Uptake of Wheat. Commun. Soil Sci. Plant Anal. 2017, 48, 756–768. [Google Scholar] [CrossRef]
- Yehuda, Z.; Shenker, M.; Romheld, V.; Marschner, H.; Hadar, Y.; Chen, Y. The Role of Ligand Exchange in the Uptake of Iron from Microbial Siderophores by Gramineous Plants. Plant Physiol. 1996, 112, 1273–1280. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Hayat, R.; Begum, F.; Bohannan, B.J.M.; Inebert, L.; Meyer, K. Variation in Soil Physical, Chemical and Microbial Parameters Under Different Land Uses in Bagrot Valley, Gilgit, Pakistan. J. Chem. Soc. Pak. 2017, 39, 97–107. [Google Scholar]
- Bhattacharyya, A.; Campbell, A.N.; Tfaily, M.M.; Lin, Y.; Kukkadapu, R.K.; Silver, W.L.; Nico, P.S.; Pett-Ridge, J. Redox Fluctuations Control the Coupled Cycling of Iron and Carbon in Tropical Forest Soils. Environ. Sci. Technol. 2018, 52, 14129–14139. [Google Scholar] [CrossRef] [Green Version]
- Chacon, N.; Silver, W.L.; Dubinsky, E.A.; Cusack, D.F. Iron Reduction and Soil Phosphorus Solubilization in Humid Tropical Forest Soils: The Roles of Labile Carbon Pools and an Electron Shuttle Compound. Biogeochemistry 2006, 78, 67–84. [Google Scholar] [CrossRef]
- Dubinsky, E.A.; Silver, W.L.; Firestone, M.K. Tropical Forest Soil Microbial Communities Couple Iron and Carbon Biogeochemistry. Ecology 2010, 91, 2604–2612. [Google Scholar] [CrossRef]
- Barcellos, D.; O’Connell, C.S.; Silver, W.; Meile, C.; Thompson, A. Hot Spots and Hot Moments of Soil Moisture explain Fluctuations in Iron and Carbon Cycling in a Humid Tropical Forest Soil. Soil Syst. 2018, 2, 59. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, P.; Pandey, A. Bioprospecting Plant Growth Promoting Endophytic Bacteria Isolated from Himalayan Yew (Taxus wallichiana Zucc.). Microbiol. Res. 2020, 239, 126536. [Google Scholar] [CrossRef]
- Dastager, S.G.; Deepa, C.K.; Pandey, A. Plant Growth Promoting Potential of Pontibacter niistensis in Cowpea (Vigna unguiculata (L.) Walp.). Appl. Soil Ecol. 2011, 49, 250–255. [Google Scholar] [CrossRef]
- Balcazar, W.; Rondon, J.; Rengifo, M.; Ball, M.M.; Melfo, A.; Gomez, W.; Yarzabal, L.A. Bioprospecting Glacial Ice for Plant growth promoting Bacteria. Microbiol. Res. 2015, 177, 1–7. [Google Scholar] [CrossRef]
- Yadav, A.N.; Sachan, S.G.; Verma, P.; Saxena, A.K. Bioprospecting of Plant Growth Promoting Psychrotrophic Bacilli from the Cold Desert of North Western Indian Himalayas. Indian J. Exp. Biol. 2016, 52, 142–150. [Google Scholar]
- Boiteau, R.M.; Shaw, J.B.; Pasa-Tolic, L.; Koppenaal, D.W.; Jansson, J.K. Micronutrient Metal Speciation is Controlled by Competitive Organic Chelation in Grassland Soils. Soil Biol. Biochem. 2018, 120, 283–291. [Google Scholar] [CrossRef]
- Kusel, K.; Wagner, C.; Trinkwalter, T.; Gossner, A.S.; Baumler, R.; Drake, H.L. Microbial Reduction of Fe(III) and Turnover of Acetate in Hawaiian Soils. FEMS Microbiol. Ecol. 2002, 40, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.; Natesan, S.K.A.; Singh, B.K.; Usha, K. Improving Zinc Efficiency of Cereals under Zinc Deficiency. Curr. Sci. 2005, 88, 36–44. [Google Scholar]
- Ullah, A.; Farooq, M.; Rehman, A.; Hussain, M.; Siddique, K.H.M. Zinc Nutrition in Chickpea (Cicer arietinum): A Review. Crop Pasture Sci. 2020, 71, 199–218. [Google Scholar] [CrossRef]
- Nepomuceno, R.A.; Brown, C.B.; Gargarino, A.M.P.; Pedro, M.S.; Brown, M.B. Growth Enhancement of Rice (Oryza sativa L.) by Zinc-Solubilising Bacteria Isolated from Vesicular-Arbuscular Mycorrhizal Root Inoculant (VAMRI). Philipp. J. Crop Sci. 2020, 45, 34–40. [Google Scholar]
- Imran, M.; Arshad, M.; Khalid, A.; Kanwal, S.; Crowley, D.E. Perspectives of Rhizosphere Microflora for Improving Zn Bioavailability and Acquisition by Higher Plants. Int. J. Agric. Biol. 2014, 16, 653–662. [Google Scholar]
- Bala, R.; Kalia, A.; Dhaliwal, S.S. Evaluation of Efficacy of ZnO Nanoparticles as Remedial Zinc Nanofertilizer for Rice. J. Soil Sci. Plant Nutr. 2019, 19, 379–389. [Google Scholar] [CrossRef]
- Ramesh, A.; Sharma, S.K.; Sharma, M.P.; Yadav, N.; Joshi, O.P. Inoculation of Zinc Solubilising Bacillus araybhattai Strains for Improved Growth, Mobilization and Biofortification of Zinc in Soybean and Wheat Cultivated in Vertisols of Central India. Appl. Soil Ecol. 2014, 73, 87–96. [Google Scholar] [CrossRef]
- Strachel, R.; Zaborowska, M.; Wyszkowska, J. Deliberations on Zinc—A Trace Mineral of a Toxic Element? J. Elem. 2016, 21, 625–639. [Google Scholar]
- Yang, X.-W.; Tian, X.-H.; Lu, X.-C.; Cao, Y.-X.; Chen, Z.-H. Impacts of Phosphorus and Zinc Levels on Phosphorus and Zinc Nutrition and phytic Acid Concentration in Wheat (Triticum aestivum L.). Sci. Food Agric. 2011, 91, 2322–2328. [Google Scholar] [CrossRef] [PubMed]
- Rengel, Z. Availability of Mn, Zn and Fe in the Rhizosphere. J. Soil Sci. Plant Nutr. 2015, 15, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Dubey, A.N.; Chattopadhyaya, N.; Mandal, N. Variation in Soil Microbial Population and Soil Enzymatic Activities Under Zincated Nanoclay Polymer Composites (ZNCPCs), Nano-ZnO and Zn Solubilizers in Rice Rhizosphere. Agric. Res. 2021, 10, 21–31. [Google Scholar] [CrossRef]
- Ash, M.; Yadav, J.; Yadav, J.S. Yield Attributes of Rice (Oryza Sativa) as Affected by Integrated Use of Zinc Oxide and Zinc Solubilizers. Indian J. Agric. Sci. 2020, 90, 2180–2184. [Google Scholar]
- Subramanian, K.S.; Tenshia, V.; Jayalakshmi, K.; Ramachandran, V. Biochemical Changes and Zinc Fractions in Arbuscular Mycorrhizal Fungi (Glomus intraradices) Inoculated and Uninoculated Soils under Differential Zinc Fertilization. Appl. Soil Ecol. 2009, 43, 32–39. [Google Scholar] [CrossRef]
- Suganya, A.; Saravanan, A.; Baskar, M.; Pandiyarajan, P.; Kavimani, R. Agronomic Biofortification of Maize (Zea mays L.) with Zinc by Using of Graded Levels of Zinc in Combination with Zinc Solubilising Bacteria and Arbuscular Mycorrhizal Fungi. J. Plant Nutr. 2020, 44, 988–994. [Google Scholar] [CrossRef]
- Vaid, S.K.; Srivastava, P.C.; Sharma, A.; Rawat, D.; Mathpal, B.; Shankhadhar, S.C.; Shukla, A.K. Residual Effect of Zinc Applied to Rice on Zinc Nutrition of Succeeding Wheat Crop Inoculated with Zinc Solubilizing Microbial Consortium. Isr. J. Plant Sci. 2019, 66, 227–237. [Google Scholar] [CrossRef]
- Liu, Y.M.; Cao, W.Q.; Chen, X.X.; Yu, B.G.; Lang, M.; Chen, X.P.; Zou, C.Q. The Responses of Soil Enzyme Activities, Microbial Biomass and Microbial Community Structure to Nine Years of Varied Zinc Application Rates. Sci. Total Environ. 2020, 737, 140245. [Google Scholar] [CrossRef]
- Qin, N.; Sun, M.T.; Zhu, L.X.; Ge, C.M.; Wang, J. Residual Effects of Soil Zn Fertilization on Soil Characteristics, Yield and Quality of Platycodon grandiflorum. Soil Sci. Plant Nutr. 2020, 66, 344–351. [Google Scholar]
- Asadishad, B.; Chahal, S.; Akbari, A.; Cianciarelli, V.; Azodi, M.; Ghoshal, S.; Tufenkji, N. Amendment of Agricultural Soil with Metal Nanoparticles: Effects of Soil Enzyme Activity and Microbial Community Composition. Environ. Sci. Technol. 2018, 52, 1908–1918. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Gomez, C.; Fernandez, M.D.; Garcia, S.; Obrador, A.F.; Leton, M.; Babin, M. Soil pH Effects the Toxicity of Zinc Oxide Nanoparticles to the Soil Microbial Community. Environ. Sci. Pollut. Res. 2018, 25, 28140–28152. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Pan, B.; Zhang, H.; Steinberg, C.E.W.; Qiu, H.; Vijver, M.G.; Peijnenburg, W.J.G.M. Application of Low Dosage of Copper Oxide and Zinc Oxide Nanoparticles Boosts Bacterial and Fungal Communities in Soil. Sci. Total Environ. 2021, 757, 143807. [Google Scholar] [CrossRef]
- Xu, J.B.; Luo, X.S.; Wang, Y.L.; Feng, Y.Z. Evaluation of Zinc Oxide Nanoparticles on Lettuce (Lactuca sativa L) Growth and Soil Bacterial Community. Environ. Sci. Pollut. Res. 2018, 25, 6026–6035. [Google Scholar] [CrossRef]
- Alloway, B.L. Zinc in Soils and Crop Nutrition, 2nd ed.; IZA: Brussels, Belgium; IFA: Paris, France, 2008. [Google Scholar]
- Boardman, R.; McGuire, D.O. The Role of Zinc in Forestry, I. Zinc in Forest Environments, Ecosystems and Tree Nutrition. For. Ecol. Manag. 1990, 37, 167–205. [Google Scholar] [CrossRef]
- Macdonald, C.A.; Clark, I.M.; Zhao, F.J.; Hirsch, P.R.; Singh, B.K.; McGrath, S.P. Long-Term Impacts of Zinc and Copper Enriched Sewage Sludge Additions on Bacterial, Archaeal and Fungal Communities in Arable and Grassland Soils. Soil Biol. Biochem. 2011, 43, 932–941. [Google Scholar] [CrossRef]
- Zaheer, A.; Malik, A.; Sher, A.; Qaisrani, M.M.; Mehmood, A.; Khan, S.U.; Ashraf, M.; Mirza, Z.; Karim, S.; Rasool, M. Isolation, Characterization, and Effect of Phosphate-Zinc-Solubilising Bacterial Strains on Chickpea (Cicer arietinum L.) Growth. Saudi J. Biol. Sci. 2019, 26, 1061–1067. [Google Scholar] [CrossRef]
- Liu, P.; Yang, Y.S.; Xu, G.D.; Fang, Y.H.; Yang, Y.A.; Kalin, R.M. The effect of molybdenum and boron in soil on the growth and photosynthesis of three soybean varieties. Plant Soil Environ. 2005, 51, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.C.; Sahu, S. Response of Chickpea to Micronutrients and Biofertilizers in Vertisol. Legume Res. 2012, 35, 248–251. [Google Scholar]
- Sun, T.; Wang, Y.P.; Wang, Z.Y.; Liu, P.; Xu, G.D. The Effects of Molybdenum and Boron on the Rhizosphere Microorganisms and Soil Enzyme Activities of Soybean. Acta Physiol. Plant. 2013, 35, 763–770. [Google Scholar] [CrossRef]
- Mandal, M.; Naik, S.K.; Das, D.K. Effect of Boron and Sulphur Interaction on Some Important Biological Indices in an Inceptisol. J. Plant Nutr. 2018, 41, 197–209. [Google Scholar]
- Abreu, I.; Cerda, M.E.; De Nanclares, M.P.; Baena, I.; Lloret, J.; Bonilla, I.; Bolanos, L.; Reguera, M. Boron Deficiency Affects Rhizobia Cell Surface Polysaccharides Important for Suppression of Plant Defence Mechanisms During Legume Recognition and for the Development of Nitrogen-Fixing Symbiosis. Plant Soil 2012, 361, 385–395. [Google Scholar] [CrossRef]
- National Institutes of Health Office of Dietary Supplements: Boron. Available online: https://ods.od.nih.gov/factsheets/Boron-HealthProfessional/ (accessed on 3 August 2021).
- Rana, M.S.; Hu, C.X.; Shaaban, M.; Imran, M.; Afzal, J.; Moussa, M.G.; Elyamine, A.M.; Bhantana, P.; Saleem, M.H.; Syaifudin, M.; et al. Soil Phosphorus Transformation Characteristics in Response to Molybdenum Supply in Leguminous Crops. J. Environ. Manag. 2020, 268, 110610. [Google Scholar] [CrossRef]
- Rana, M.S.; Sun, X.C.; Imran, M.; Ali, S.; Shaaban, M.; Moussa, M.G.; Khan, Z.; Afzal, J.; Binyamin, R.; Bhantana, P.; et al. Hu CX Molybdenum-Induced Effects on Leaf Ultra-Structure Rhizosphere Phosphorus Transformation in Triticum aestivum, L. Plant Physiol. Biochem. 2020, 153, 20–29. [Google Scholar] [CrossRef]
- Ge, X.X.; Vaccaro, B.J.; Thorgersen, M.P.; Poole, F.L.; Majumder, E.L.; Zane, G.M.; De Leon, K.B.; Lancaster, W.A.; Moon, J.W.; Paradis, C.J.; et al. Iron- and Aluminium- Induced Depletion of Molybdenum in Acidic Environments Impedes the Nitrogen Cycle. Environ. Microbiol. 2019, 21, 152–163. [Google Scholar] [CrossRef] [Green Version]
- Rajakumari, R.; Oluwafemi, O.S.; Thomas, S.; Kalarikkal, N. Dietary Supplements Containing Vitamins and Minerals: Formulation, Optimization and Evaluation. Powder Technol. 2018, 336, 481–492. [Google Scholar] [CrossRef]
- Hua, T.W.; Zhang, R.; Sun, H.W.; Liu, C.G. Alleviation of Boron Toxicity in Plants: Mechanisms and Approaches. Crit. Rev. Environ. Sci. Technol. 2020, 51, 2975–3015. [Google Scholar] [CrossRef]
- Adhikari, L.; Missaoui, A.M. Nodulation Response to Molybdenum Supplementation in Alfalfa and its Correlation with Root and Shoot Growth in Low pH Soil. J. Plant Nutr. 2017, 40, 2290–2302. [Google Scholar] [CrossRef]
- Kwiatkowski, C.A.; Harasim, E. Chemical Properties of Soil in Four-Field Crop Rotations under Organic and Conventional Farming Systems. Agronomy 2020, 10, 1045. [Google Scholar] [CrossRef]
- Nadeem, F.; Farooq, M.; Nawaz, A.; Ahmad, R. Boron Improves Productivity and Profitability of Bread Wheat under Zero and Plough Tillage on Alkaline Calcareous Soil. Field Crops Res. 2019, 239, 1–9. [Google Scholar] [CrossRef]
- Vera, A.; Moreno, J.L.; Siles, J.A.; Lopez-Mondejar, R.; Zhou, Y.; Li, Y.; Garcia, C.; Nicolas, E.; Bastida, F. Interactive Impacts of Boron and Organic Amendments in Plant-Soil Microbial Relationships. J. Hazard. Mater. 2021, 408, 124939. [Google Scholar] [CrossRef]
- Bilen, S.; Bilen, M.; Bardhan, S. The Effects of Boron Management on Soil Microbial Population and Enzyme Activities. Afr. J. Biotechnol. 2011, 10, 5311–5319. [Google Scholar]
- Mahmud, K.; Makaju, S.; Ibrahim, R.; Missaoui, A. Current Progress in Nitrogen Fixing Plants and Microbiome Research. Plants 2020, 9, 97. [Google Scholar] [CrossRef] [Green Version]
- Parniske, M. Intracellular Accommodation of Microbes by Plants: A Common Developmental Program for Symbiosis and Disease? Curr. Opin. Plant Biol. 2000, 3, 320–328. [Google Scholar] [CrossRef]
- Zehirov, G.; Georgiev, G. Relationships Between Cell Membrane Stability, Exudate Content and Infectivity of Bradyrhizobium japonicum Strain 639 to Boron Staved Soybean Plants. Acta Physiol. Plant 2006, 28, 171–179. [Google Scholar] [CrossRef]
- Zhang, Y.; Gladyshev, V.N. Molybdoproteomes and Evolution of Molybdenum Utilisation. J. Mol. Biol. 2008, 379, 881–899. [Google Scholar] [CrossRef] [Green Version]
- Kristek, S.; Resic, I.; Jovic, J.; Zmajic, K.; Lenart, L.; Kraljicak, Z.; Beslo, D.; Rasic, S. Effect of Various Rates of Boron on Yield and Quality of High-Grade Sugar Beet Varieties. Lis. Cukrov. A Repar. 2018, 134, 146–150. [Google Scholar]
- Vera, A.; Moreno, J.L.; Garcia, C.; Morais, D.; Bastida, F. Boron in Soil: The Impacts on the Biomass, Composition and Activity of the Soil Microbial Community. Sci. Total Environ. 2019, 685, 564–573. [Google Scholar] [CrossRef]
- Barron, A.R.; Wurzburger, N.; Bellenger, J.P.; Wright, S.J.; Kraepiel, A.M.L.; Hedin, L.O. Molybdenum Limitation of Asymbiotic Nitrogen Fixation in Tropical Forest Soils. Nat. Geosci. 2009, 2, 42–45. [Google Scholar] [CrossRef]
- Wen, X.; Hu, C.X.; Sun, X.C.; Zhao, X.H.; Tan, Q.L.; Liu, P.J.; Xin, J.; Qin, S.Y.; Wang, P.C. Characterisation of Vegetable Nitrogen Uptake and Soil Nitrogen Transformation in Response to Continuous Molybdenum Application. J. Plant Nutr. Soil Sci. 2018, 181, 516–527. [Google Scholar] [CrossRef]
- Franczuk, J.; Rosa, R.; Zaniewicz-Bajkowska, A.; Slonecka, D. Effects of Boron Application and Treatment with Effective Microorganisms on the Growth, Yield and some Quality Attributes of Broccoli. J. Elem. 2019, 24, 1335–1348. [Google Scholar]
- Shcherbakova, E.N.; Shcherbakov, A.V.; Andronov, E.E.; Gonchar, L.N.; Kalenskaya, S.M.; Chebotar, V.K. Combined Pre-Seed Treatment with Microbial Inoculants and Mo Nanoparticles Changes Composition of Root Exudates and Rhizosphere Microbiome Structure of Chickpea (Cicer arietinum L.) Plants. Symbiosis 2017, 73, 57–59. [Google Scholar] [CrossRef]
- Taran, N.Y.; Gonchar, O.M.; Lopatko, K.G.; Batsmanova, L.M.; Patyka, M.V.; Volkogon, M.V. The Effect of Colloidal Solution of Molybdenum Nanoparticles on the Microbial Composition in Rhizosphere of Cicer arietinum L. Nanoscale Res. Lett. 2014, 9, 289. [Google Scholar] [CrossRef] [Green Version]
- Lehto, T.; Smolander, A.; Aphalo, P.J. Decomposition and Element Concentrations of Silver Birch Leaf Litter as Affected by Boron Status of Litter and Soil. Plant Soil 2010, 329, 195–208. [Google Scholar] [CrossRef]
- Reed, S.C.; Cleveland, C.C.; Townsend, A.R. Relationships among Phosphorus, Molybdenum and Free-Living Nitrogen Fixation in Tropical Rainforests: Results from Observational and Experimental Analyses. Biogeochemistry 2013, 114, 135–147. [Google Scholar] [CrossRef]
- Wichard, T.; Mishra, B.; Myneni, S.C.B.; Bellenger, J.P.; Kraepiel, A.M.L. Storage and Bioavailability of Molybdenum in Soils Increased by Organic Matter Complexation. Nat. Geosci. 2009, 2, 625–629. [Google Scholar] [CrossRef]
- Liermann, L.J.; Guynn, R.L.; Anbar, A.; Brantley, S.L. Production of Molybdophore during Metal Targeted Dissolution of Silicates by Soil Bacteria. Chem. Geol. 2005, 220, 285–302. [Google Scholar] [CrossRef]
- Zulfiqar, U.; Hussain, S.; Ishfaq, M.; Ali, N.; Ahmad, M.; Ihsan, F.; Sheteiwy, M.S.; Rauf, A.; Hano, C.; El-Esawi, M.A. Manganese Supply Improves Bread Wheat Productivity, Economic Returns and Grain Biofortification Under Conventional and No Tillage Systems. Agriculture 2021, 11, 142. [Google Scholar] [CrossRef]
- Mukhopadhyay, M.J.; Sharma, A. Manganese in Cell Metabolism in Higher Plants. Bot. Rev. 1991, 57, 117–149. [Google Scholar] [CrossRef]
- World Health Organisation: Regional Office of Europe (WHO) Chapter 6.8: Manganese. 2001. Available online: https://www.euro.who.int/__data/assets/pdf_file/0003/123078/AQG2ndEd_6_8Manganese.pdf (accessed on 11 August 2021).
- Zulfiqar, U.; Hussain, S.; Ishfaq, M.; Ali, N.; Yasin, M.U.; Ali, M.A. Foliar Manganese Supply Enhances Crop Productivity, Net Benefits, and Grain Manganese Accumulation in Direct-Seeded and Puddled Transplanted Rice. J. Plant Growth Regul. 2021, 40, 1539–1556. [Google Scholar] [CrossRef]
- Yang, W.H.; Zhang, Z.; Chen, H.; Liu, J.; Ali, M.; Liu, F.; Li, L. Population Structure of Manganese-Oxidising Bacteria in Stratified Soils and Properties of Manganese Oxide Aggregates Under Manganese-Complex Medium Enrichment. PLoS ONE 2013, 8, e73778. [Google Scholar]
- Terzano, R.; Cuccovillo, G.; Gattullo, C.E.; Medici, L.; Tomasi, N.; Pinton, R.; Mimmo, T.; Cesco, S. Combined Effect of Soil Organic Acids and Flavonoids on the Mobilization of Major and Trace Elements from Soil. Biol. Fertil. Soils 2015, 51, 685–695. [Google Scholar] [CrossRef]
- Zhang, F.S. Mobilization of Iron and Manganese by Plant-Borne and Synthetic Metal Chelators. Plant Soil 1993, 155, 111–114. [Google Scholar] [CrossRef]
- Williams, C.H.; David, D.J. Effects of Pasture Improvement with Subterranean Clover and Superphosphate on the Availability of Trace Metals to Plants. Aust. J. Soil Res. 1976, 14, 85–93. [Google Scholar] [CrossRef]
- Bowles, T.M.; Mooshammer, M.; Socolar, Y.; Calderon, F.; Cavigelli, M.A.; Culman, S.W.; Deen, W.; Drury, C.F.; Y Garcia, A.G.; Gaudin, A.C.M.; et al. Long-Term Evidence Shows that Crop-Rotation Diversification Increases Agricultural Resilience to Adverse Growing Conditions in North America. One Earth 2020, 2, 284–293. [Google Scholar] [CrossRef]
- Ogut, M.; Er, F. Micronutrient Composition of Field-Grown Dry Bean and Wheat Inoculated with Azospirillum and Trichoderma. J. Plant Nutr. Soil Sci. 2006, 169, 699–703. [Google Scholar] [CrossRef]
- Posta, K.; Marschner, H.; Romheld, V. Manganese Reduction in the Rhizosphere of Mycorrhizal and Nonmycorrhizal Maize. Mycorrhiza 1994, 5, 119–124. [Google Scholar] [CrossRef]
- Kothari, S.K.; Marschner, H.; Romheld, V. Effect of Vesicular Arbuscular Mycorrhizal Fungus and Rhizosphere Microorganisms on Manganese Reduction in the Rhizosphere and Manganese Concentrations in Maize (Zea-mays L.). New Phytol. 1991, 117, 649–655. [Google Scholar] [CrossRef]
- Piromyou, P.; Buranabanyat, B.; Tantasawat, P.; Tittabutr, P.; Boonkerd, N.; Teaumroong, N. Effect of Plant Growth Promoting Rhizobacteria (PGPR) Inoculation on Microbial Community Structure in Rhizosphere of Forage Corn Cultivated in Thailand. Eur. J. Soil Biol. 2011, 47, 44–54. [Google Scholar] [CrossRef]
- Rana, A.; Saharan, B.; Nain, L.; Prasanna, R.; Shivay, Y.S. Enhancing Micronutrient Uptake and Yield of Wheat through Bacterial PGPR Consortia. Soil Sci. Plant Nutr. 2012, 58, 573–582. [Google Scholar] [CrossRef]
- Marschner, P.; Ascher, J.S.; Graham, R.D. Effect of Manganese-Reducing Rhizosphere Bacteria on the Growth of Gaeumannomyces graminis var. tritici and on Manganese Uptake by Wheat (Triticum aestivum L.). Biol. Fertil. Soil. 1991, 12, 33–38. [Google Scholar] [CrossRef]
- Innangi, M.; Schenk, M.K.; D’Alessandro, F.; Pinto, S.; Menta, C.; Papa, S.; Fioretto, A. Field and Microcosms Decomposition Dynamics of European Beech Leaf Litter: Influence of Climate, Plant Material and Soil with Focus on N and Mn. Appl. Soil Ecol. 2015, 93, 88–97. [Google Scholar] [CrossRef]
- Jones, M.E.; LaCroix, R.E.; Zeigler, J.; Ying, S.C.; Nico, P.S.; Keiluweit, M. Enzymes, Manganese, or Iron? Drivers of Oxidative Organic Matter Decomposition in Soils. Environ. Sci. Technol. 2020, 54, 14114–14123. [Google Scholar] [CrossRef]
- Sun, T.; Cui, Y.; Berg, B.; Zhang, Q.; Dong, L.; Wu, Z.; Zhang, L. A Test of Manganese Effects on Decomposition in Forest and Cropland Sites. Soil Biol. Biochem. 2019, 129, 178–183. [Google Scholar] [CrossRef]
- Whalen, E.D.; Smith, R.G.; Grandy, S.; Frey, S.D. Manganese Limitation as a Mechanism for Reduced Decomposition in Soils under Atmospheric Nitrogen Deposition. Soil Biol. Biochem. 2018, 127, 252–263. [Google Scholar] [CrossRef]
- Keiluweit, M.; Nico, P.; Harmon, M.E.; Mao, J.D.; Pett-Ridge, J.; Kleber, M. Long-Term Litter Decomposition Controlled by Manganese Redox Cycling. Proc. Natl. Acad. Sci. USA 2015, 112, E5253–E5260. [Google Scholar] [CrossRef] [Green Version]
- Alguacil, M.D.; Torres, M.P.; Montesinos-Navarro, A.; Roldan, A. Soil Characteristics Driving Arbuscular Mycorrhizal Fungal Communities in Semiarid Mediterranean Soils. Appl. Environ. Microbiol. 2016, 82, 3348–3356. [Google Scholar] [CrossRef] [Green Version]
- Collins, R.N.; Kinsela, A.S. Pedogenic Factors and Measurements of the Plant Uptake of Cobalt. Plant Soil 2011, 339, 499–512. [Google Scholar] [CrossRef]
- Banerjee, P.; Bhattacharya, P. Investigating Cobalt in Soil-Plant-Animal-Human System: Dynamics, Impact and Management. J. Soil Sci. Plant Nutr. 2021, 21, 2339–2354. [Google Scholar] [CrossRef]
- Ahmed, S.; Evans, H.J. Cobalt: A micronutrient Element for the Growth of Soybean Plants under Symbiotic Conditions. Soil Sci. 1960, 90, 205–210. [Google Scholar] [CrossRef]
- Akbar, F.M.; Zafar, M.; Hamid, A.; Ahmed, M.; Khaliq, A.; Khan, M.R.; Rehman, Z.U. Interactive Effect of Cobalt and Nitrogen Growth on Growth, Nodulation, Yield and Protein Content of Field Grown Pea. Hortic. Environ. Biotechnol. 2013, 54, 465–474. [Google Scholar] [CrossRef]
- Wilson, S.B.; Hallsworth, E.G. Studies on the Nutrition of Forage Legumes: IV The Effect of Cobalt on the Growth and Nodulated and Non-Nodulated Trifolium Subterraneum. Plant Soil 1965, 2, 260–279. [Google Scholar] [CrossRef]
- Warren, M.J.; Raux, E.; Schubert, H.L.; Escalante-Semerena, J.C. The Biosynthesis of Adenosylcobalamin (Vitamin B12). Nat. Prod. Rep. 2002, 19, 390–412. [Google Scholar] [CrossRef]
- Zhang, Y.; Rodionov, D.A.; Gelfand, M.S.; Gladyshev, V.N. Comparative Genomic Analyses of Nickel, Cobalt and Vitamin B12 Utilisation. BMC Genom. 2009, 10, 78. [Google Scholar] [CrossRef] [Green Version]
- Rod, N.K.; Gudadhe, N.N.; Karmakar, N.; Mehta, P.V.; Narwade, A.V. Cobalt Chloride Enhances Crop Duration, Increases Production, and Productivity of Chickpea. J. Plant Nutri. 2019, 42, 40–57. [Google Scholar] [CrossRef]
- Amir, H.; Pineau, R. Relationships between Extractable Ni, Co, and Other Metals and Some Microbiological Characteristics in Different Ultramafic Soils from New Caledonia. Aust. J. Soil Res. 2003, 41, 215–228. [Google Scholar] [CrossRef]
- Amir, H.; Pineau, R. Release of Ni and Co by Microbial Activity in New Caledonian Ultramafic Soils. Can. J. Microbiol. 2003, 49, 288–293. [Google Scholar] [CrossRef]
- Okamoto, S.; Eltis, L.D. The Biological Occurrence and Trafficking of Cobalt. Metallomics 2011, 3, 963–970. [Google Scholar] [CrossRef]
- Gikas, P. Single and Combined Effects of Nickel (Ni(II)) and Cobalt (Co(II)) Ions on Activated Sludge and on Other Aerobic Microorganisms: A Review. J. Hazard. Mater. 2008, 159, 187–203. [Google Scholar] [CrossRef]
- Quantin, C.; Becquer, T.; Rouiller, J.H.; Berthelin, J. Oxide Weathering and Trace Metal Release by bacterial Reduction in a New Caledonia Ferralsol. Biogeochemistry 2001, 53, 323–340. [Google Scholar] [CrossRef]
- Chaudhari, D.; Rangappa, K.; Das, A.; Layek, J.; Basavaraj, S.; Kandpal, B.K.; Shouche, Y.; Rahi, P. Pea (Pisum Sativuml.) Plant Shapes its Rhizosphere Microbiome for Nutrient Uptake and Stress Amelioration in Acidic Soils of the North-East Region of India. Front. Microbiol. 2020, 11, 968. [Google Scholar] [CrossRef] [PubMed]
- Inaba, S.; Takenaka, C. Changes in Chemical Species of Copper Added to Brown Forest Soil in Japan. Water Air Soil Pollut. 2005, 162, 285–293. [Google Scholar] [CrossRef]
- Schoffer, J.T.; Sauve, S.; Neaman, A.; Ginocchio, R. Role of Leaf Litter on the Incorporation of Copper-Containing Pesticides into Soils under Fruit Production: A Review. J. Soil Sci. Plant Nutr. 2020, 10, 990–1000. [Google Scholar] [CrossRef]
- Klevay, L.M. Is the Western Diet Adequate in Copper? J. Trace Elem. Med. Biol. 2011, 25, 204–212. [Google Scholar] [CrossRef]
- Jenkins, S.N.; Murphy, D.V.; Waite, I.S.; Rushton, S.P.; O’Donnell, A.G. Ancient Landscapes and the Relationship with Microbial Nitrification. Sci. Rep. 2016, 6, 30733. [Google Scholar] [CrossRef]
- Mishra, S.; Keswani, C.; Abhilash, P.C.; Fraceto, L.F.; Singh, H.B. Integrated Approach of Agri-Nanotechnology: Challenges and Future Trends. Front. Plant Sci. 2017, 8, 471. [Google Scholar] [CrossRef] [Green Version]
- Simonin, M.; Colman, B.P.; Tang, W.Y.; Judy, J.D.; Andersono, S.M.; Bergemanno, C.M.; Rocca, J.D.; Unrine, J.M.; Cassar, N.; Bernhardt, E.S. Plant and Microbial Response to Repeated Cu(OH)2 Nanopesticide Exposures Under Different Fertilization Levels in an Agro-Ecosystem. Front. Microbiol. 2018, 9, 1769. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.Y.; Su, X.X.; Wang, Y.Y.; Yang, X.Y.; Bi, M.; He, Q.; Chen, Y. Copper Oxide Nanoparticles Inhibited Denitrifying Enzymes and Electron Transport System Activities to Influence Soil Denitrification and N2O Emission. Chemosphere 2020, 245, 125394. [Google Scholar] [CrossRef]
- Adak, A.; Prasanna, R.; Babu, S.; Bidyarani, N.; Verma, S.; Pal, M.; Shivay, Y.S.; Nain, L. Micronutrient Enrichment Mediated by Plant-Microbe Interactions and Rice Cultivation Practices. J. Plant Nutr. 2016, 39, 1216–1232. [Google Scholar] [CrossRef]
- Keiblinger, K.M.; Schneider, M.; Gorfer, M.; Paumann, M.; Deltedesco, E.; Berger, H.; Jochlinger, L.; Mentler, A.; Zechmeister-Boltenstern, S.; Soja, G.; et al. Assessment of Cu Applications in Two Contrasting Soil-Effects on Soil Microbial Activity and Fungal Community Structure. Ecotoxicology 2018, 27, 217–233. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denton-Thompson, S.M.; Sayer, E.J. Micronutrients in Food Production: What Can We Learn from Natural Ecosystems? Soil Syst. 2022, 6, 8. https://doi.org/10.3390/soilsystems6010008
Denton-Thompson SM, Sayer EJ. Micronutrients in Food Production: What Can We Learn from Natural Ecosystems? Soil Systems. 2022; 6(1):8. https://doi.org/10.3390/soilsystems6010008
Chicago/Turabian StyleDenton-Thompson, Sarah M., and Emma J. Sayer. 2022. "Micronutrients in Food Production: What Can We Learn from Natural Ecosystems?" Soil Systems 6, no. 1: 8. https://doi.org/10.3390/soilsystems6010008
APA StyleDenton-Thompson, S. M., & Sayer, E. J. (2022). Micronutrients in Food Production: What Can We Learn from Natural Ecosystems? Soil Systems, 6(1), 8. https://doi.org/10.3390/soilsystems6010008