


Growth Performance, Physiological Responses, and Histoarchitectural Changes in Juvenile Pangasianodon hypophthalmus under Different Environmental Salinities

 ,
,  ,
,  , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Calculations for Evaluating the Fish Growth and Survival
2.3. Monitoring the Water Quality
2.4. Proximate Chemical Composition of Whole-Body
2.5. Sampling Procedures
2.6. Biochemical Analyses
2.7. Histopathological Examination
2.8. Statistical Analysis
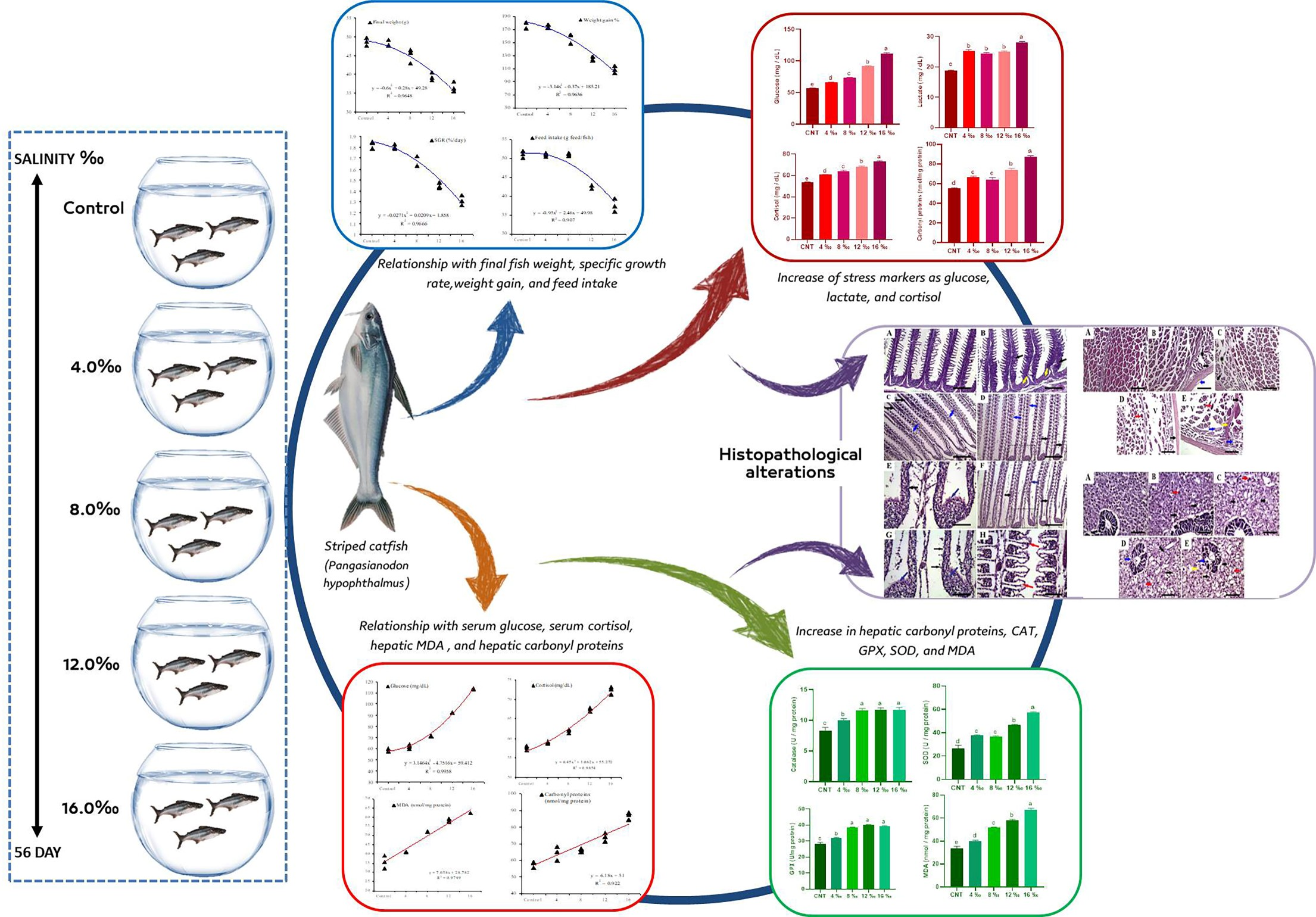
3. Results
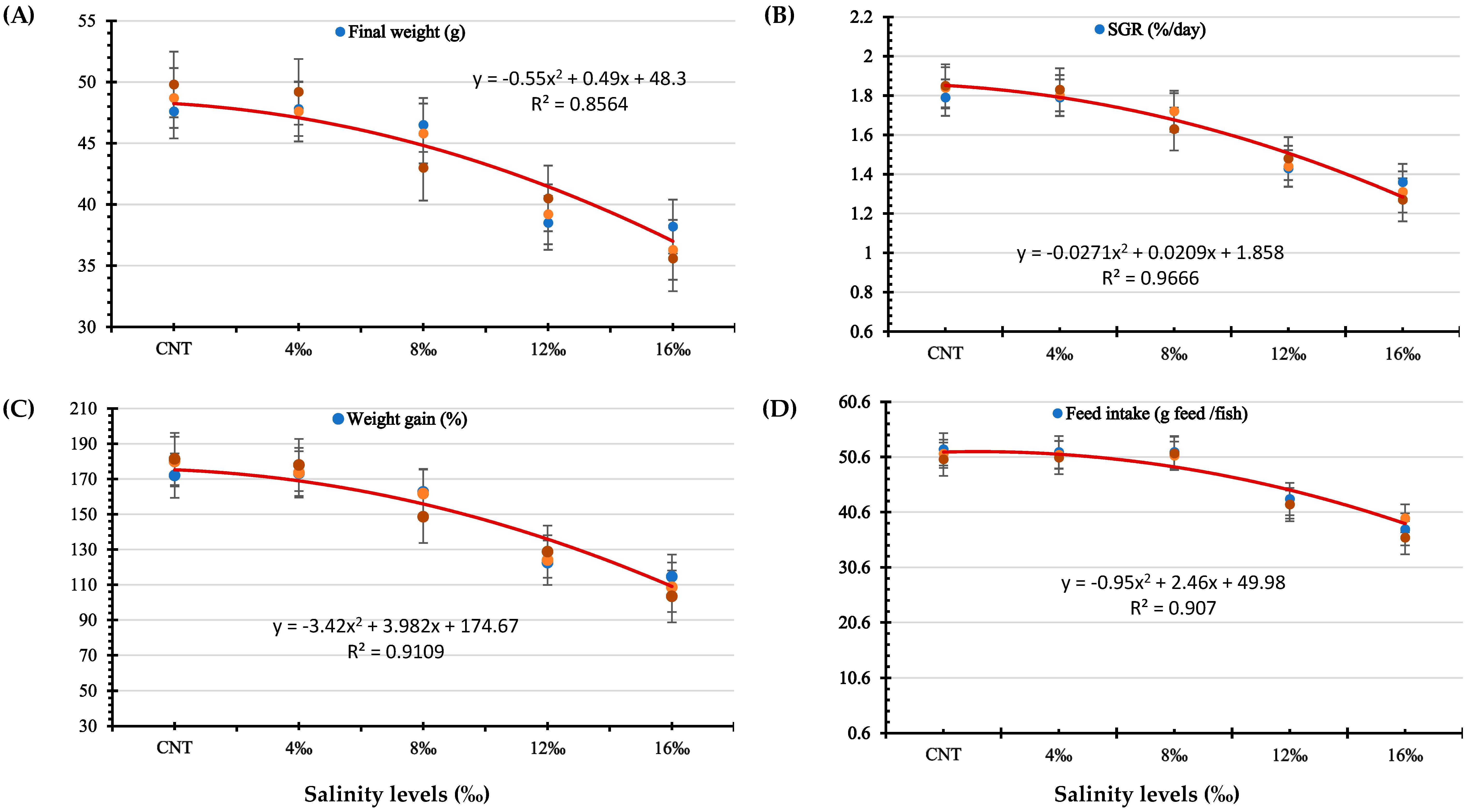
3.1. Growth Performance and Survival Rates
3.2. Whole-Body Chemical Composition
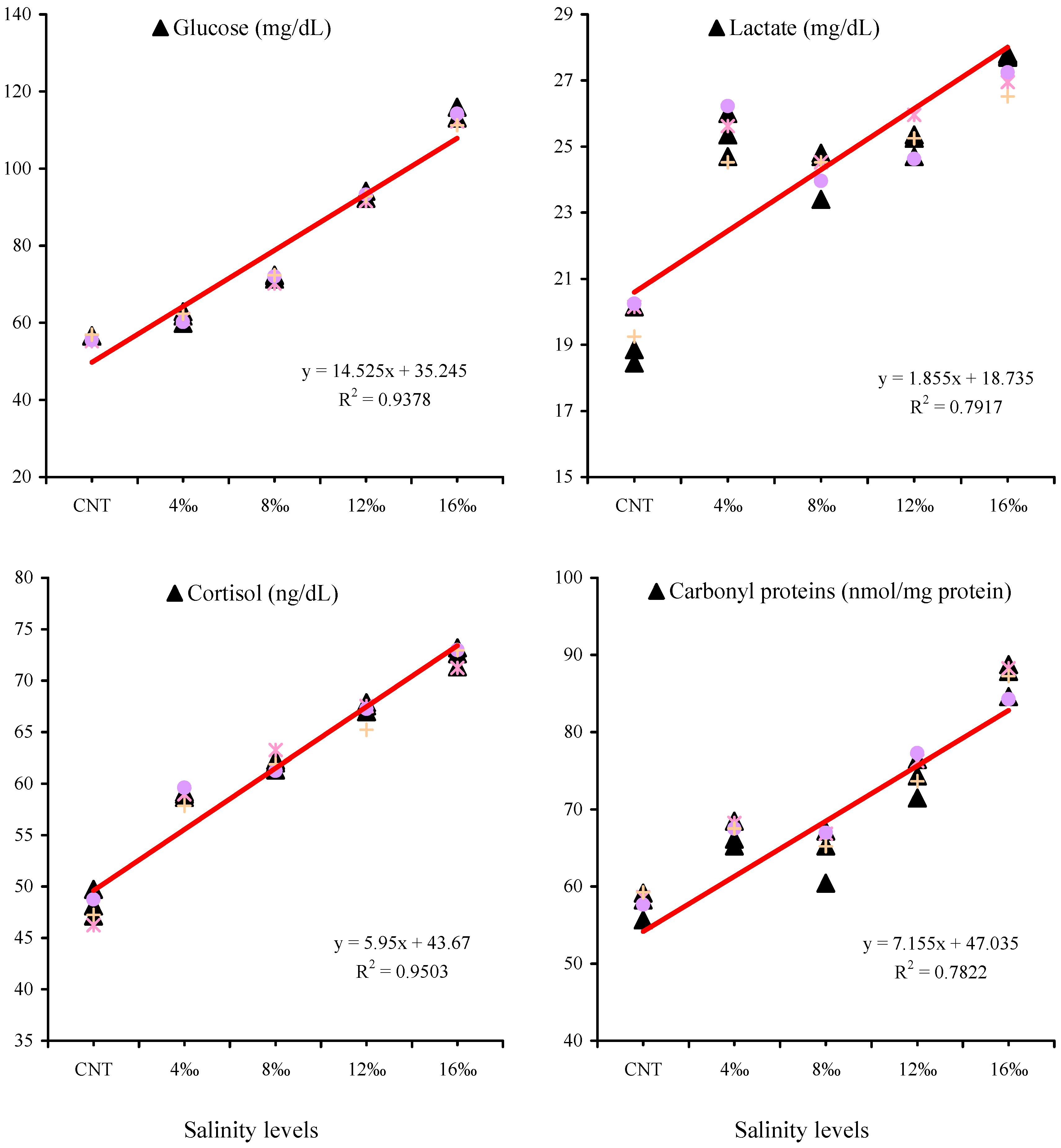
3.3. Serum Stress Biomarkers and Liver Antioxidant Parameters
3.4. Histopathological Alterations
3.4.1. Effects of Different Salinities on Fish Muscular Tissues
3.4.2. Effects of Different Salinities on the Hepatopancreatic Tissues
3.4.3. Effects of Different Salinities on the Fish Gill Tissues
3.5. Histopathological Scores of the Examined Fish Tissues
4. Discussion
5. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shukry, M.; Abd El-Kader, M.F.; Hendam, B.M.; Dawood, M.A.O.; Farrag, F.A.; Aboelenin, S.M.; Soliman, M.M.; Abdel-Latif, H.M.R. Dietary Aspergillus oryzae modulates serum biochemical indices, immune responses, oxidative stress, and transcription of HSP70 and cytokine genes in Nile tilapia exposed to salinity stress. Animals 2021, 11, 1621. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, Y.-M.; Xu, W.-B.; Chen, D.-Y.; Li, B.-W.; Cheng, Y.-X.; Guo, X.-L.; Dong, W.-R.; Shu, M.-A. The effects of salinities stress on histopathological changes, serum biochemical index, non-specific immune and transcriptome analysis in red swamp crayfish Procambarus clarkii. Sci. Total Environ. 2022, 840, 156502. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Li, E.; Suo, Y.; Su, Y.; Lu, M.; Zhao, Q.; Qin, J.G.; Chen, L. Histological and transcriptomic responses of two immune organs, the spleen and head kidney, in Nile tilapia (Oreochromis niloticus) to long-term hypersaline stress. Fish Shellfish Immunol. 2018, 76, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Chacoff, L.; Calvo, Á.; Ruiz-Jarabo, I.; Villarroel, F.; Muñoz, J.L.; Tinoco, A.B.; Cárdenas, S.; Mancera, J.M. Growth performance, osmoregulatory and metabolic modifications in red porgy fry, Pagrus pagrus, under different environmental salinities and stocking densities. Aquac. Res. 2011, 42, 1269–1278. [Google Scholar] [CrossRef]
- Herrera, M.; Aragão, C.; Hachero, I.; Ruiz-Jarabo, I.; Vargas-Chacoff, L.; Mancera, J.M.; Conceição, L.E.C. Physiological short-term response to sudden salinity change in the Senegalese sole (Solea senegalensis). Fish Physiol. Biochem. 2012, 38, 1741–1751. [Google Scholar] [CrossRef]
- Arjona, F.J.; Vargas-Chacoff, L.; Ruiz-Jarabo, I.; Martín del Río, M.P.; Mancera, J.M. Osmoregulatory response of Senegalese sole (Solea senegalensis) to changes in environmental salinity. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 148, 413–421. [Google Scholar] [CrossRef]
- Laiz-Carrión, R.; Guerreiro, P.M.; Fuentes, J.; Canario, A.V.M.; Martín Del Río, M.P.; Mancera, J.M. Branchial osmoregulatory response to salinity in the gilthead sea bream, Sparus auratus. J. Exp. Zool. Part A Comp. Exp. Biol. 2005, 303A, 563–576. [Google Scholar] [CrossRef]
- Ruiz-Jarabo, I.; Tinoco, A.B.; Vargas-Chacoff, L.; Martos-Sitcha, J.A.; Rodríguez-Rúa, A.; Cárdenas, S.; Mancera, J.M. Environmental salinity affects growth and metabolism in fingerling meagre (Argyrosomus regius). Fishes 2019, 4, 6. [Google Scholar] [CrossRef]
- Gan, L.; Xu, Z.X.; Ma, J.J.; Xu, C.; Wang, X.D.; Chen, K.; Chen, L.Q.; Li, E.C. Effects of salinity on growth, body composition, muscle fatty acid composition, and antioxidant status of juvenile Nile tilapia Oreochromis niloticus (Linnaeus, 1758). J. Appl. Ichthyol. 2016, 32, 372–374. [Google Scholar] [CrossRef]
- Bal, A.; Panda, F.; Pati, S.G.; Das, K.; Agrawal, P.K.; Paital, B. Modulation of physiological oxidative stress and antioxidant status by abiotic factors especially salinity in aquatic organisms. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 241, 108971. [Google Scholar] [CrossRef]
- Takata, R.; Mattioli, C.C.; Bazzoli, N.; Júnior, J.D.C.; Luz, R.K. The effects of salinity on growth, gill tissue and muscle cellularity in Lophiosilurus alexandri juvenile, a Neotropical freshwater catfish. Aquac. Res. 2021, 52, 4064–4075. [Google Scholar] [CrossRef]
- Ahmed, N.; Thompson, S.; Glaser, M. Global aquaculture productivity, environmental sustainability, and climate change adaptability. Environ. Manag. 2019, 63, 159–172. [Google Scholar] [CrossRef]
- Dubey, S.K.; Trivedi, R.K.; Chand, B.K.; Mandal, B.; Rout, S.K. Farmers’ perceptions of climate change, impacts on freshwater aquaculture and adaptation strategies in climatic change hotspots: A case of the Indian Sundarban delta. Environ. Dev. 2017, 21, 38–51. [Google Scholar] [CrossRef]
- McCormick, S.D.; Bradshaw, D. Hormonal control of salt and water balance in vertebrates. Gen. Comp. Endocrinol. 2006, 147, 3–8. [Google Scholar] [CrossRef]
- Ali, H.; Haque, M.M.; Belton, B. Striped catfish (Pangasianodon hypophthalmus, Sauvage, 1878) aquaculture in Bangladesh: An overview. Aquac. Res. 2013, 44, 950–965. [Google Scholar] [CrossRef]
- Phan, L.T.; Bui, T.M.; Nguyen, T.T.T.; Gooley, G.J.; Ingram, B.A.; Nguyen, H.V.; Nguyen, P.T.; De Silva, S.S. Current status of farming practices of striped catfish, Pangasianodon hypophthalmus in the Mekong Delta, Vietnam. Aquaculture 2009, 296, 227–236. [Google Scholar] [CrossRef]
- Abd-Elaziz, R.A.; Shukry, M.; Abdel-Latif, H.M.R.; Saleh, R.M. Growth-promoting and immunostimulatory effects of phytobiotics as dietary supplements for Pangasianodon hypophthalmus fingerlings. Fish Shellfish Immunol. 2023, 133, 108531. [Google Scholar] [CrossRef]
- De Silva, S.S.; Phuong, N.T. Striped catfish farming in the Mekong Delta, Vietnam: A tumultuous path to a global success. Rev. Aquac. 2011, 3, 45–73. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Ahmed, H.A.; Shukry, M.; Chaklader, M.R.; Saleh, R.M.; Khallaf, M.A. Astragalus membranaceus Extract (AME) Enhances Growth, Digestive Enzymes, Antioxidant Capacity, and Immunity of Pangasianodon hypophthalmus Juveniles. Fishes 2022, 7, 319. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Chaklader, M.R.; Shukry, M.; Ahmed, H.A.; Khallaf, M.A. A multispecies probiotic modulates growth, digestive enzymes, immunity, hepatic antioxidant activity, and disease resistance of Pangasianodon hypophthalmus fingerlings. Aquaculture 2023, 563, 738948. [Google Scholar] [CrossRef]
- Zaki, M.A.A.; Khalil, H.S.; Allam, B.W.; Khalil, R.H.; Basuini, M.F.E.; Nour, A.E.-A.M.; Labib, E.M.H.; Elkholy, I.S.E.; Verdegem, M.; Abdel-Latif, H.M.R. Assessment of zootechnical parameters, intestinal digestive enzymes, haemato-immune responses, and hepatic antioxidant status of Pangasianodon hypophthalmus fingerlings reared under different stocking densities. Aquac. Int. 2023, 1–24. [Google Scholar] [CrossRef]
- Nguyen, P.T.H.; Do, H.T.T.; Mather, P.B.; Hurwood, D.A. Experimental assessment of the effects of sublethal salinities on growth performance and stress in cultured tra catfish (Pangasianodon hypophthalmus). Fish Physiol. Biochem. 2014, 40, 1839–1848. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.C.; Kadir, S.; Hossain, A. Effects of salinity on the growth, survival and proximate composition of Pangas, Pangasius hypophthalmus. Bangladesh J. Zool. 2020, 48, 141–149. [Google Scholar] [CrossRef]
- Ha, N.T.K.; Em, N.T.; Ngoc, N.M.; Takagi, Y.; Phuong, N.T.; Huong, D.T.T. Effects of salinity on growth performance, survival rate, digestive enzyme activities and physiological parameters of striped catfish (Pangasianodon hypophthalmus) at larval stage. Can Tho Univ. J. Sci. 2021, 13, 1–9. [Google Scholar]
- Schmitz, M.; Douxfils, J.; Mandiki, S.N.M.; Morana, C.; Baekelandt, S.; Kestemont, P. Chronic hyperosmotic stress interferes with immune homeostasis in striped catfish (Pangasianodon hypophthalmus, S.) and leads to excessive inflammatory response during bacterial infection. Fish Shellfish Immunol. 2016, 55, 550–558. [Google Scholar] [CrossRef]
- Schmitz, M.; Mandiki, S.N.M.; Douxfils, J.; Ziv, T.; Admon, A.; Kestemont, P. Synergic stress in striped catfish (Pangasianodon hypophthalmus, S.) exposed to chronic salinity and bacterial infection: Effects on kidney protein expression profile. J. Proteom. 2016, 142, 91–101. [Google Scholar] [CrossRef]
- Oanh, D.T.H.; Phu, T.Q. Effect of different salinities on the susceptibility of striped catfish (Pangasianodon hypophthalmus) to Aeromonas hydrophila bacteria causing hemorrhagic disease. Can Tho Univ. J. Sci. 2021, 13, 20–25. [Google Scholar]
- Jahan, A.; Nipa, T.T.; Islam, S.M.M.; Uddin, M.H.; Islam, M.S.; Shahjahan, M. Striped catfish (Pangasianodon hypophthalmus) could be suitable for coastal aquaculture. J. Appl. Ichthyol. 2019, 35, 994–1003. [Google Scholar] [CrossRef]
- Hieu, D.Q.; Hang, B.T.B.; Huong, D.T.T.; Kertaoui, N.E.; Farnir, F.; Phuong, N.T.; Kestemont, P. Salinity affects growth performance, physiology, immune responses and temperature resistance in striped catfish (Pangasianodon hypophthalmus) during its early life stages. Fish Physiol. Biochem. 2021, 47, 1995–2013. [Google Scholar] [CrossRef]
- Hieu, D.Q.; Hang, B.T.B.; Lokesh, J.; Garigliany, M.-M.; Huong, D.T.T.; Yen, D.T.; Liem, P.T.; Tam, B.M.; Hai, D.M.; Son, V.N.; et al. Salinity significantly affects intestinal microbiota and gene expression in striped catfish juveniles. Appl. Microbiol. Biotechnol. 2022, 106, 3245–3264. [Google Scholar] [CrossRef]
- Hossain, F.; Islam, S.M.M.; Islam, M.S.; Shahjahan, M. Behavioral and histo-pathological indices of striped catfish (Pangasionodon hypophthalmus) exposed to different salinities. Aquac. Rep. 2022, 23, 101038. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Fish and Shrimp; National Academy Press: Washington, DC, USA, 2011; p. 392. [Google Scholar]
- AOAC. Official Methods of Analysis, 13th ed.; Association of Analytical Chemists: Washington, DC, USA, 2012; p. 1018. [Google Scholar]
- Reznick, A.Z.; Packer, L. Oxidative Damage to Proteins: Spectrophotometric Method for Carbonyl Assay. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1994; Volume 233, pp. 357–363. [Google Scholar]
- Bancroft, J.D.; Gamble, M. The Hematoxylin and Eosin. In Theory and Practice of Histological Techniques, 7th ed.; Suvarna, S.K., Layton, C., Bancroft, J.D., Eds.; Churchill Livingstone: Edinburgh, UK; New York, NY, USA, 2013; pp. 179–220. [Google Scholar]
- Yossa, R.; Verdegem, M. Misuse of multiple comparison tests and underuse of contrast procedures in aquaculture publications. Aquaculture 2015, 437, 344–350. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Noreldin, A.E.; Sewilam, H. Long term salinity disrupts the hepatic function, intestinal health, and gills antioxidative status in Nile tilapia stressed with hypoxia. Ecotoxicol. Environ. Saf. 2021, 220, 112412. [Google Scholar] [CrossRef]
- Mohamed, N.A.; Saad, M.F.; Shukry, M.; El-Keredy, A.M.S.; Nasif, O.; Van Doan, H.; Dawood, M.A.O. Physiological and ion changes of Nile tilapia (Oreochromis niloticus) under the effect of salinity stress. Aquac. Rep. 2021, 19, 100567. [Google Scholar] [CrossRef]
- Abass, N.Y.; Elwakil, H.E.; Hemeida, A.A.; Abdelsalam, N.R.; Ye, Z.; Su, B.; Alsaqufi, A.S.; Weng, C.-C.; Trudeau, V.L.; Dunham, R.A. Genotype–environment interactions for survival at low and sub-zero temperatures at varying salinity for channel catfish, hybrid catfish and transgenic channel catfish. Aquaculture 2016, 458, 140–148. [Google Scholar] [CrossRef]
- Laiz-Carrión, R.; Sangiao-Alvarellos, S.; Guzmán, J.M.; Martín del Río, M.P.; Míguez, J.M.; Soengas, J.L.; Mancera, J.M. Energy Metabolism in Fish Tissues Related to Osmoregulation and Cortisol Action. Fish Physiol. Biochem. 2002, 27, 179–188. [Google Scholar] [CrossRef]
- Soengas, J.L.; Sangiao-Alvarellos, S.; Laiz-Carrión, R.; Mancera, J.M. Energy Metabolism and Osmotic Acclimation in Teleost Fish. In Fish Osmoregulation; CRC Press: Boca Raton, FL, USA, 2019; pp. 277–307. [Google Scholar]
- Islam, M.J.; Kunzmann, A.; Thiele, R.; Slater, M.J. Effects of extreme ambient temperature in European seabass, Dicentrarchus labrax acclimated at different salinities: Growth performance, metabolic and molecular stress responses. Sci. Total Environ. 2020, 735, 139371. [Google Scholar] [CrossRef]
- Ruiz-Jarabo, I.; Barany, A.; Jerez-Cepa, I.; Mancera, J.M.; Fuentes, J. Intestinal response to salinity challenge in the Senegalese sole (Solea senegalensis). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2017, 204, 57–64. [Google Scholar] [CrossRef]
- Phuc, N.T.H.; Mather, P.B.; Hurwood, D.A. Effects of sublethal salinity and temperature levels and their interaction on growth performance and hematological and hormonal levels in tra catfish (Pangasianodon hypophthalmus). Aquac. Int. 2017, 25, 1057–1071. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Monier, M.N. Stimulatory effect of dietary taurine on growth performance, digestive enzymes activity, antioxidant capacity, and tolerance of common carp, Cyprinus carpio L., fry to salinity stress. Fish Physiol. Biochem. 2018, 44, 639–649. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Noreldin, A.E.; Sewilam, H. Blood biochemical variables, antioxidative status, and histological features of intestinal, gill, and liver tissues of African catfish (Clarias gariepinus) exposed to high salinity and high-temperature stress. Environ. Sci. Pollut. Res. 2022, 29, 56357–56369. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Harikrishna, V.; Reddy, A.K.; Chadha, N.; Babitha, A. Salinity tolerance of Pangasianodon hypophthalmus in inland saline water: Effect on growth, survival and haematological parameters. Ecol. Environ. Conserv. 2017, 23, 475–482. [Google Scholar]
- Kumar, A.; Krishna, V.H.; Reddy, A.; Chadha, N.; Rani, A.B. Effect of salinity on proximate composition of Pangasianodon hypophthalmus reared in inland saline water. Int. J. Zool. Stud. 2016, 3, 19–21. [Google Scholar]
- Xu, J.; Shui, C.; Shi, Y.; Yuan, X.; Liu, Y.; Xie, Y. Effect of Salinity on Survival, Growth, Body Composition, Oxygen Consumption, and Ammonia Excretion of Juvenile Spotted Scat. North Am. J. Aquac. 2020, 82, 54–62. [Google Scholar] [CrossRef]
- Barton, B.A. Stress in fishes: A diversity of responses with particular reference to changes in circulating corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Pankhurst, N.W. The endocrinology of stress in fish: An environmental perspective. Gen. Comp. Endocrinol. 2011, 170, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Bonga, S.W. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Mozanzadeh, M.T.; Safari, O.; Oosooli, R.; Mehrjooyan, S.; Najafabadi, M.Z.; Hoseini, S.J.; Saghavi, H.; Monem, J. The effect of salinity on growth performance, digestive and antioxidant enzymes, humoral immunity and stress indices in two euryhaline fish species: Yellowfin seabream (Acanthopagrus latus) and Asian seabass (Lates calcarifer). Aquaculture 2021, 534, 736329. [Google Scholar] [CrossRef]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef]
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef]
- Nordberg, J.; Arnér, E.S.J. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radic. Biol. Med. 2001, 31, 1287–1312. [Google Scholar] [CrossRef]
- Chang, C.-H.; Wang, Y.-C.; Lee, T.-H. Hypothermal stress-induced salinity-dependent oxidative stress and apoptosis in the livers of euryhaline milkfish, Chanos chanos. Aquaculture 2021, 534, 736280. [Google Scholar] [CrossRef]
- Paital, B.; Chainy, G.B.N. Antioxidant defenses and oxidative stress parameters in tissues of mud crab (Scylla serrata) with reference to changing salinity. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2010, 151, 142–151. [Google Scholar] [CrossRef]
- Rivera-Ingraham, G.A.; Barri, K.; Boël, M.; Farcy, E.; Charles, A.-L.; Geny, B.; Lignot, J.-H. Osmoregulation and salinity-induced oxidative stress: Is oxidative adaptation determined by gill function? J. Exp. Biol. 2016, 219, 80–89. [Google Scholar] [CrossRef]
- Paital, B.; Chainy, G.B.N. Effects of salinity on O2 consumption, ROS generation and oxidative stress status of gill mitochondria of the mud crab Scylla serrata. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2012, 155, 228–237. [Google Scholar] [CrossRef]
- Bernet, D.; Schmidt, H.; Meier, W.; Burkhardt-Holm, P.; Wahli, T. Histopathology in fish: Proposal for a protocol to assess aquatic pollution. J. Fish Dis. 1999, 22, 25–34. [Google Scholar] [CrossRef]
- Hughes, G.M.; Morgan, M. The structure of fish gills in relation to their respiratory function. Biol. Rev. 1973, 48, 419–475. [Google Scholar] [CrossRef]
- Wilson, J.M.; Laurent, P. Fish gill morphology: Inside out. J. Exp. Zool. 2002, 293, 192–213. [Google Scholar] [CrossRef]
- Wedemeyer, G.A. Interactions with Water Quality Conditions. In Physiology of Fish in Intensive Culture Systems; Wedemeyer, G.A., Ed.; Springer: Boston, MA, USA, 1996; pp. 60–110. [Google Scholar]
- Khafaga, A.F.; Naiel, M.A.E.; Dawood, M.A.O.; Abdel-Latif, H.M.R. Dietary Origanum vulgare essential oil attenuates cypermethrin-induced biochemical changes, oxidative stress, histopathological alterations, apoptosis, and reduces DNA damage in Common carp (Cyprinus carpio). Aquat. Toxicol. 2020, 228, 105624. [Google Scholar] [CrossRef]
- Poleksić, V.; Mitrović-Tutundžić, V. Fish Gills as a Monitor of Sublethal and Chronic Effects of Pollution. In Sublethal and Chronic Effects of Pollutants on Freshwater Fish; Muller, R., Lloyd, R., Eds.; Fishing News Books Ltd.: Farnham, UK, 1994; pp. 339–352. [Google Scholar]
- Azodi, M.; Bahabadi, M.N.; Ghasemi, A.; Morshedi, V.; Mozanzadeh, M.T.; Shahraki, R.; Khademzadeh, O.; Hamedi, S.; Avizhgan, S. Effects of salinity on gills’ chloride cells, stress indices, and gene expression of Asian seabass (Lates calcarifer, Bloch, 1790). Fish Physiol. Biochem. 2021, 47, 2027–2039. [Google Scholar] [CrossRef]
- Carmona, R.; García-Gallego, M.; Sanz, A.; Domezaín, A.; Ostos-Garrido, M.V. Chloride cells and pavement cells in gill epithelia of Acipenser naccarii: Ultrastructural modifications in seawater-acclimated specimens. J. Fish Biol. 2004, 64, 553–566. [Google Scholar] [CrossRef]
- Maetz, J.; Keynes, R.D. Fish gills: Mechanisms of salt transfer in fresh water and sea water. Philos. Trans. R. Soc. London. B Biol. Sci. 1971, 262, 209–249. [Google Scholar] [CrossRef]
- Kültz, D. Physiological mechanisms used by fish to cope with salinity stress. J. Exp. Biol. 2015, 218, 1907–1914. [Google Scholar] [CrossRef] [PubMed]
- Van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Stentiford, G.D.; Longshaw, M.; Lyons, B.P.; Jones, G.; Green, M.; Feist, S.W. Histopathological biomarkers in estuarine fish species for the assessment of biological effects of contaminants. Mar. Environ. Res. 2003, 55, 137–159. [Google Scholar] [CrossRef]
- Johnston, I.A. Muscle development and growth: Potential implications for flesh quality in fish. Aquaculture 1999, 177, 99–115. [Google Scholar] [CrossRef]
- Johnston, I.A. Environment and plasticity of myogenesis in teleost fish. J. Exp. Biol. 2006, 209, 2249–2264. [Google Scholar] [CrossRef]
- Periago, M.J.; Ayala, M.D.; López-Albors, O.; Abdel, I.; Martínez, C.; García-Alcázar, A.; Ros, G.; Gil, F. Muscle cellularity and flesh quality of wild and farmed sea bass, Dicentrarchus labrax L. Aquaculture 2005, 249, 175–188. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Salinity Levels (‰) | p Value | ||||
|---|---|---|---|---|---|---|
| 0.0 (Control) | 4.0 | 8.0 | 12.0 | 16.0 | ||
| Initial weight (Wt0; g) | 17.5 ± 0.09 | 17.3 ± 0.09 | 17.5 ± 0.12 | 17.5 ± 0.12 | 17.6 ± 0.12 | 0.990 |
| Final weight (Wt56; g) | 48.7 ± 0.64 a | 48.2 ± 0.53 a | 45.1 ± 1.07 b | 39.4 ± 0.59 c | 36.7 ± 0.78 d | < 0.001 |
| Weight gain % | 177.3 ± 2.91 a | 178.6 ± 1.55 a | 157.7 ± 4.56 b | 125.1 ± 1.89 c | 108.5 ± 3.23 d | < 0.001 |
| Specific growth rate (%/day) | 1.83 ± 0.019 a | 1.83 ± 0.015 a | 1.69 ± 0.052 b | 1.45 ± 0.022 c | 1.31 ± 0.043 d | < 0.001 |
| Feed intake (g feed/fish) | 52.0 ± 0.13 a | 51.0 ± 0.29 a | 51.2 ± 0.19 a | 42.3 ± 0.33 b | 37.7 ± 1.01 b | < 0.001 |
| Feed conversion ratio | 1.67 ± 0.032 b | 1.65 ± 0.032 b | 1.86 ± 0.065 a | 1.93 ± 0.055 a | 1.97 ± 0.073 a | 0.005 |
| Fish survival (%) | 100.0 ± 0.00 a | 100.0 ± 0.00 a | 100.0 ± 0.00 a | 91.1 ± 2.22 b | 77.8 ± 2.22 c | < 0.001 |
| Parameters (%) | Salinity Levels (‰) | p Value | ||||
|---|---|---|---|---|---|---|
| 0.0 (Control) | 4.0 | 8.0 | 12.0 | 16.0 | ||
| Moisture | 76.2 ± 0.08 a | 76.8 ± 0.36 a | 74.7 ± 0.38 b | 74.6 ± 0.58 b | 73.7 ± 0.36 b | 0.001 |
| Crude protein | 14.8 ± 0.11 b | 14.5 ± 0.26 b | 15.8 ± 0.20 a | 15.9 ± 0.42 a | 16.0 ± 0.48 a | 0.023 |
| Crude lipids | 4.2 ± 0.08 c | 4.2 ± 0.04 c | 4.7 ± 0.10 b | 5.2 ± 0.21 a | 5.2 ± 0.20 a | 0.001 |
| Ash | 3.1 ± 0.03 | 2.8 ± 0.03 | 3.0 ± 0.06 | 2.8 ± 0.15 | 2.9 ± 0.23 | 0.480 |
| Organs | Scored Lesions | Salinity Levels (‰) | p Value | ||||
|---|---|---|---|---|---|---|---|
| 0.0 | 4.0 | 8.0 | 12.0 | 16.0 | |||
| Muscular tissues | Dermis separation | 3.00 | 6.83 | 5.66 | 11.66 | 12.83 | 0.0261 |
| Atrophy of muscle bundles | 3.50 | 5.00 | 6.50 | 12.00 | 13.00 | 0.0162 | |
| Hyalinization of myotomes | 4.00 | 4.00 | 7.00 | 12.00 | 13.00 | 0.0160 | |
| Splitting and vacuolization of muscle fibers | 4.00 | 5.50 | 5.50 | 12.50 | 12.50 | 0.0147 | |
| Liver | Vascular congestion | 4.00 | 4.00 | 9.00 | 11.50 | 11.50 | 0.0215 |
| Vacuolization | 3.33 | 4.50 | 7.83 | 10.83 | 13.50 | 0.0171 | |
| Necrosis | 5.00 | 5.00 | 5.00 | 11.50 | 13.50 | 0.0087 | |
| Gills | Epithelial hyperplasia and fusion | 2.50 | 5.16 | 8.00 | 11.33 | 13.00 | 0.0156 |
| Epithelial necrosis and rupture | 4.50 | 4.50 | 6.16 | 12.33 | 12.50 | 0.0177 | |
| Lamellar epithelial lifting and edema | 3.00 | 4.00 | 11.00 | 11.00 | 11.00 | 0.0087 | |
| Chloride cells hyperplasia | 2.00 | 5.00 | 8.50 | 11.50 | 13.00 | 0.0100 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Latif, H.M.R.; Ahmed, H.A.; Shukry, M.; Khafaga, A.F.; Elkhayat, B.K.; Abdel-Tawwab, M.; Abd-elaziz, R.A. Growth Performance, Physiological Responses, and Histoarchitectural Changes in Juvenile Pangasianodon hypophthalmus under Different Environmental Salinities. Fishes 2023, 8, 282. https://doi.org/10.3390/fishes8060282
Abdel-Latif HMR, Ahmed HA, Shukry M, Khafaga AF, Elkhayat BK, Abdel-Tawwab M, Abd-elaziz RA. Growth Performance, Physiological Responses, and Histoarchitectural Changes in Juvenile Pangasianodon hypophthalmus under Different Environmental Salinities. Fishes. 2023; 8(6):282. https://doi.org/10.3390/fishes8060282
Chicago/Turabian StyleAbdel-Latif, Hany M. R., Hamada A. Ahmed, Mustafa Shukry, Asmaa F. Khafaga, Bassem Khalil Elkhayat, Mohsen Abdel-Tawwab, and Rehab A. Abd-elaziz. 2023. "Growth Performance, Physiological Responses, and Histoarchitectural Changes in Juvenile Pangasianodon hypophthalmus under Different Environmental Salinities" Fishes 8, no. 6: 282. https://doi.org/10.3390/fishes8060282
APA StyleAbdel-Latif, H. M. R., Ahmed, H. A., Shukry, M., Khafaga, A. F., Elkhayat, B. K., Abdel-Tawwab, M., & Abd-elaziz, R. A. (2023). Growth Performance, Physiological Responses, and Histoarchitectural Changes in Juvenile Pangasianodon hypophthalmus under Different Environmental Salinities. Fishes, 8(6), 282. https://doi.org/10.3390/fishes8060282