
In Vitro Evaluation of the Effects of BAP Concentration and Pre-Cooling Treatments on Morphological, Physiological, and Biochemical Traits of Different Olive (Olea euorpea L.) Cultivars


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Explant Disinfection, Media Preparation, and Treatments
2.2. In Vitro Propagation and Data Collection
2.3. Measurement of Growth Traits
2.4. Assessment of Physiological Parameters
2.5. Assessment ofBiochemical Contents
2.5.1. Quantification of Primary Metabolites
2.5.2. Quantification of Secondary Metabolites
2.6. Statistical Analysis
3. Results
3.1. Growth Traits
3.2. Physiological Parameters
3.3. Biochemical Contents
3.3.1. Primary Metabolites
3.3.2. Secondary Metabolites

{kind=link}
{kind=link}
{kind=link}
| Treatments | Alkaloids mg (g DW)−1 | Flavonoids mg (g DW)−1 | Tannins mg (g DW)−1 | Phenols mg (g DW)−1 |
|---|---|---|---|---|
| Pre-cooling (T) (Hours) | ||||
| T1 (0 h) | 1.52 ± 0.074 c | 1.35 ± 0.067 b | 0.63 ± 0.015 a | 0.70 ± 0.025 a |
| T2(24 h) | 1.75 ± 0.077 b | 1.44 ± 0.077 ab | 0.65 ± 0.014 a | 0.85 ± 0.026 a |
| T3 (48 h) | 2.00 ± 0.09 a | 1.60 ± 0.082 a | 0.72 ± 0.015 a | 0.93 ± 0.026 a |
| LSD | 0.18 | 0.15 | 0.17 | 0.14 |
| Varieties (V) | ||||
| V1 (Leccino) | 1.5 ± 0.093 d | 1.39 ± 0.084 c | 1.45 ± 0.084 a | 1.00 ± 0.049 d |
| V2 (Gemlik) | 1.70 ± 0.10 c | 1.42 ± 0.080 bc | 1.43 ± 0.080 a | 1.03 ± 0.055 bc |
| V3 (Morailo) | 1.84 ± 0.10 b | 1.50 ± 0.095 ab | 1.49 ± 0.095 a | 1.08 ± 0.059 ab |
| V4 (Arbosana) | 2.5 ± 0.01 a | 1.54 ± 0.098 a | 1.50 ± 0.098 a | 1.5 ± 0.064 a |
| LSD | 0.045 | 0.09 | 0.1 | 0.06 |
| BAP (LB) (mgL−1) | ||||
| L1 (0) | 1.02 ± 0.031 d | 0.84 ± 0.035 d | 0.68 ± 0.017 d | 0.73 ± 0.026 d |
| L2 (0.5) | 1.64 ± 0.054 b | 1.20 ± 0.049 c | 0.80 ± 0.027 c | 1.04 ± 0.035 c |
| L3 (1.5) | 1.48 ± 0.054 c | 1.42 ± 0.054 b | 1.01 ± 0.032 b | 1.27 ± 0.031 a |
| L4(2.5) | 1.85 ± 0.062 a | 1.62 ± 0.068 a | 1.11 ± 0.027 a | 1.16 ± 0.033 b |
| LSD | 0.023 | 0.040 | 0.055 | 0.028 |
| Significance | ||||
| T | * | ** | ns | ns |
| V | ** | ** | ns | * |
| LB | ** | ** | ** | ** |
| T × V | ns | ns | ns | ns |
| T × LB | * | * | ns | * |
| V × LB | * | * | ns | ns |
| T × V × LB | ns | ns | ns | ns |
| PCT (T) (hours) | BAP (LB) (mgL−1) | LPS (cm) | PIS | NLPS | NSPE | Chlorophyll (μg cm−2) | Carotenoids (μg cm−2) | Glucose mg g−1 DW | Fructose mg g−1 DW | Sucrose mg g−1 DW | Proline μg g−1 FW | Alkaloids mg g−1 DW | Flavanoids mg g−1 DW | Phenols mg g−1 DW |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T1 (0 h) | L1 (0) | 1.00 ± 0.08 | 36.7 ± 1.41 | 2.45 ± 0.08 | 1.06 ± 0.03 | 18.5 ± 0.37 | 2.62 ± 0.07 | 0.16 ± 0.01 | 0.005 ± 0.0007 | 0.02 ± 0.001 | 46.7 ± 0.3 | 0.85 ± 0.03 | 0.75 ± 0.03 | 0.69 ± 0.02 |
| L2 (0.5) | 1.69 ± 0.12 | 46.0 ± 2.32 | 3.15 ± 0.17 | 1.74 ± 0.10 | 23.8 ± 0.39 | 3.60 ± 0.11 | 0.47 ± 0.009 | 0.012 ± 0.001 | 0.10 ± 0.007 | 44.7 ± 0.44 | 1.22 ± 0.052 | 0.94 ± 0.05 | 0.85 ± 0.034 | |
| L3 (1.5) | 1.94 ± 0.01 | 52.4 ± 2.94 | 3.79 ± 0.14 | 2.12 ± 0.11 | 27.5 ± 0.41 | 4.22 ± 0.9 | 0.58 ± 0.010 | 0.020 ± 0.002 | 0.21 ± 0.003 | 42.0 ± 0.90 | 1.420 ± 0.058 | 1.13 ± 0.05 | 1.001 ± 0.050 | |
| L4 (2.5) | 2.19 ± 0.07 | 57.0 ± 2.77 | 4.22 ± 0.15 | 2.40 ± 0.12 | 33.5 ± 0.52 | 4.81 ± 0.14 | 0.74 ± 0.011 | 0.028 ± 0.002 | 0.25 ± 0.004 | 22.8 ± 0.37 | 1.58 ± 0.06 | 1.28 ± 0.06 | 1.11 ± 0.04 | |
| T2 (24 h) | L1 (0) | 1.04 ± 0.02 | 42.0 ± 0.73 | 2.49 ± 0.05 | 1.21 ± 0.02 | 19.5 ± 0.31 | 2.78 ± 0.06 | 0.22 ± 0.005 | 0.008 ± 0.0003 | 0.05 ± 0.008 | 48.5 ± 0.52 | 1.08 ± 0.03 | 1.001 ± 0.04 | 0.87 ± 0.034 |
| L2 (0.5) | 2.04 ± 0.06 | 55.0 ± 1.88 | 3.25 ± 0.08 | 2.23 ± 0.12 | 27.4 ± 0.30 | 4.03 ± 0.10 | 0.56 ± 0.005 | 0.020 ± 0.001 | 0.21 ± 0.011 | 45.4 ± 0.72 | 1.44 ± 0.04 | 1.20 ± 0.03 | 1.07 ± 0.04 | |
| L3 (1.5) | 2.29 ± 0.01 | 61.2 ± 1.88 | 4.25 ± 0.16 | 2.57 ± 0.12 | 33.3 ± 0.55 | 4.94 ± 0.12 | 0.68 ± 0.006 | 0.034 ± 0.001 | 0.26 ± 0.01 | 42.8 ± 0.48 | 1.58 ± 0.05 | 1.40 ± 0.05 | 1.22 ± 0.03 | |
| L4 (2.5) | 2.57 ± 0.07 | 65.3 ± 1.54 | 4.70 ± 0.16 | 2.98 ± 0.12 | 37.4 ± 0.67 | 5.40 ± 0.9 | 0.85 ± 0.001 | 0.043 ± 0.0021 | 0.31 ± 0.010 | 24.7 ± 0.31 | 1.77 ± 0.05 | 1.61 ± 0.07 | 1.31 ± 0.03 | |
| T3 (48 h) | L1 (0) | 1.13 ± 0.03 | 41.3 ± 0.59 | 2.50 ± 0.01 | 1.20 ± 0.01 | 20.8 ± 0.29 | 2.72 ± 0.10 | 0.23 ± 0.008 | 0.007 ± 0.0004 | 0.07 ± 0.005 | 50.5 ± 0.51 | 1.12 ± 0.03 | 1.04 ± 0.03 | 0.93 ± 0.025 |
| L2 (0.5) | 2.37 ± 0.11 | 62.0 ± 2.15 | 3.79 ± 0.16 | 2.33 ± 0.17 | 31.4 ± 0.72 | 4.87 ± 0.13 | 0.63 ± 0.009 | 0.025 ± 0.001 | 0.27 ± 0.007 | 49.5 ± 0.74 | 1.77 ± 0.04 | 1.46 ± 0.03 | 1.20 ± 0.02 | |
| L3 (1.5) | 2.67 ± 0.13 | 68.4 ± 2.50 | 4.53 ± 0.12 | 2.93 ± 0.20 | 36.5 ± 0.67 | 5.37 ± 0.13 | 0.75 ± 0.009 | 0.037 ± 0.002 | 0.32 ± 0.008 | 45.0 ± 0.84 | 1.94 ± 0.04 | 1.69 ± 0.02 | 1.28 ± 0.01 | |
| L4 (2.5) | 2.89 ± 0.14 | 72.8 ± 2.58 | 4.83 ± 0.11 | 3.57 ± 0.28 | 41.8 ± 0.63 | 6.12 ± 0.17 | 0.91 ± 0.011 | 0.050 ± 0.002 | 0.36 ± 0.011 | 22.5 ± 0.39 | 2.20 ± 0.03 | 1.97 ± 0.04 | 1.39 ± 0.01 | |
| LSD | 0.1388 | 2.2443 | 0.1944 | 0.1422 | 1.0253 | 0.2206 | 0.0289 | 0.0028 | 0.0102 | 2.1257 | 0.0421 | 0.0700 | 0.0500 |
| Varieties (V) | BAP (LB) (mgL−1) | LPS | PIS | NLPS | NSPE | Chlorophyll (μg cm−2) | Carotenoids (μg cm−2) | Fructose mg g−1 DW | Sucrose mg g−1 DW | Alkaloids mg g−1 DW | Flavanoids mg g−1 DW |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Leccino (V1) | L1 (0) | 0.87 ± 0.07 | 39.1 ± 2.16 | 2.27 ± 0.07 | 1.11 ± 0.04 | 11.1 ± 0.30 | 2.64 ± 0.10 | 0.006 ± 0.0008 | 0.03 ± 0.008 | 0.93 ± 0.06 | 0.81 ± 0.04 |
| L2 (0.5) | 1.67 ± 0.15 | 47.1 ± 2.95 | 2.91 ± 0.15 | 1.64 ± 0.12 | 18.1 ± 1.18 | 3.87 ± 0.23 | 0.016 ± 0.002 | 0.17 ± 0.02 | 1.29 ± 0.10 | 1.06 ± 0.07 | |
| L3 (1.5) | 1.92 ± 0.12 | 53.1 ± 3.08 | 3.66 ± 0.14 | 1.94 ± 0.01 | 22.4 ± 1.37 | 4.62 ± 0.1 | 0.026 ± 0.003 | 0.24 ± 0.01 | 1.48 ± 0.10 | 1.24 ± 0.07 | |
| L4 (2.5) | 2.21 ± 0.10 | 57.2 ± 3.38 | 4.07 ± 0.15 | 2.26 ± 0.11 | 25.6 ± 1.44 | 5.10 ± 0.1 | 0.035 ± 0.005 | 0.28 ± 0.01 | 1.65 ± 0.13 | 1.41 ± 0.07 | |
| Gemlik (V2) | L1 (0) | 1.9 ± 0.03 | 39.4 ± 1.27 | 2.49 ± 0.04 | 1.17 ± 0.04 | 11.6 ± 0.20 | 2.87 ± 0.04 | 0.006 ± 0.0007 | 0.05 ± 0.001 | 0.97 ± 0.05 | 0.94 ± 0.05 |
| L2 (0.5) | 1.97 ± 0.13 | 54.6 ± 2.41 | 3.22 ± 0.08 | 2.01 ± 0.14 | 18.7 ± 1.39 | 3.97 ± 0.25 | 0.015 ± 0.002 | 0.19 ± 0.02 | 1.43 ± 0.08 | 1.17 ± 0.06 | |
| L3 (1.5) | 2.22 ± 0.11 | 60.2 ± 2.49 | 4.11 ± 0.18 | 2.46 ± 0.1 | 23.1 ± 1.57 | 4.62 ± 0.24 | 0.029 ± 0.003 | 0.26 ± 0.01 | 1.54 ± 0.09 | 1.34 ± 0.05 | |
| L4 (2.5) | 2.51 ± 0.10 | 65.0 ± 2.40 | 4.49 ± 0.14 | 3.01 ± 0.2 | 27.0 ± 1.52 | 5.27 ± 0.23 | 0.038 ± 0.004 | 0.29 ± 0.01 | 1.79 ± 0.13 | 1.53 ± 0.05 | |
| Morailo (V3) | L1 (0) | 1.12 ± 0.04 | 42.4 ± 1.42 | 2.59 ± 0.03 | 1.16 ± 0.02 | 11.2 ± 0.32 | 2.69 ± 0.07 | 0.007 ± 0.002 | 0.05 ± 0.001 | 1.02 ± 0.03 | 0.94 ± 0.05 |
| L2 (0.5) | 2.19 ± 0.12 | 56.1 ± 3.40 | 3.61 ± 0.16 | 2.24 ± 0.11 | 19.6 ± 1.53 | 4.34 ± 0.20 | 0.020 ± 0.003 | 0.20 ± 0.02 | 1.58 ± 0.09 | 1.23 ± 0.04 | |
| L3 (1.5) | 2.42 ± 0.16 | 62.4 ± 3.61 | 4.32 ± 0.11 | 2.76 ± 0.16 | 24.1 ± 1.91 | 4.80 ± 0.17 | 0.017 ± 0.004 | 0.27 ± 0.01 | 1.73 ± 0.07 | 1.47 ± 0.02 | |
| L4 (2.5) | 2.62 ± 0.16 | 67.0 ± 2.71 | 4.81 ± 0.14 | 3.23 ± 0.27 | 28.2 ± 1.73 | 5.70 ± 0.25 | 0.040 ± 0.003 | 0.32 ± 0.02 | 1.92 ± 009 | 1.71 ± 0.03 | |
| Arbosana (V4) | L1 (0) | 1.12 ± 0.04 | 41.2 ± 0.94 | 2.57 ± 0.11 | 1.19 ± 0.03 | 11.6 ± 0.20 | 2.62 ± 0.13 | 0.007 ± 0.0006 | 0.06 ± 0.001 | 1.13 ± 0.05 | 1.03 ± 0.03 |
| L2 (0.5) | 2.29 ± 0.14 | 59.7 ± 4.06 | 3.86 ± 0.15 | 2.51 ± 0.15 | 20.6 ± 1.52 | 4.50 ± 0.2 | 0.023 ± 0.002 | 0.21 ± 0.02 | 1.60 ± 0.09 | 1.35 ± 0.07 | |
| L3 (1.5) | 2.62 ± 0.16 | 67.0 ± 4.48 | 4.67 ± 0.13 | 3.01 ± 0.17 | 25.2 ± 1.95 | 5.24 ± 0.2 | 0.037 ± 0.002 | 0.28 ± 0.01 | 1.82 ± 0.09 | 1.57 ± 0.06 | |
| L4 (2.5) | 2.86 ± 0.17 | 71.1 ± 4.45 | 4.96 ± 0.14 | 3.44 ± 0.26 | 28.7 ± 2.11 | 5.61 ± 0.34 | 0.048 ± 0.004 | 0.33 ± 0.02 | 2.04 ± 0.08 | 1.84 ± 0.05 | |
| LSD | 0.1610 | 2.588 | 0.2234 | 0.1644 | 1.2170 | 0.2659 | 0.0045 | 0.0118 | 0.0482 | 0.0809 |
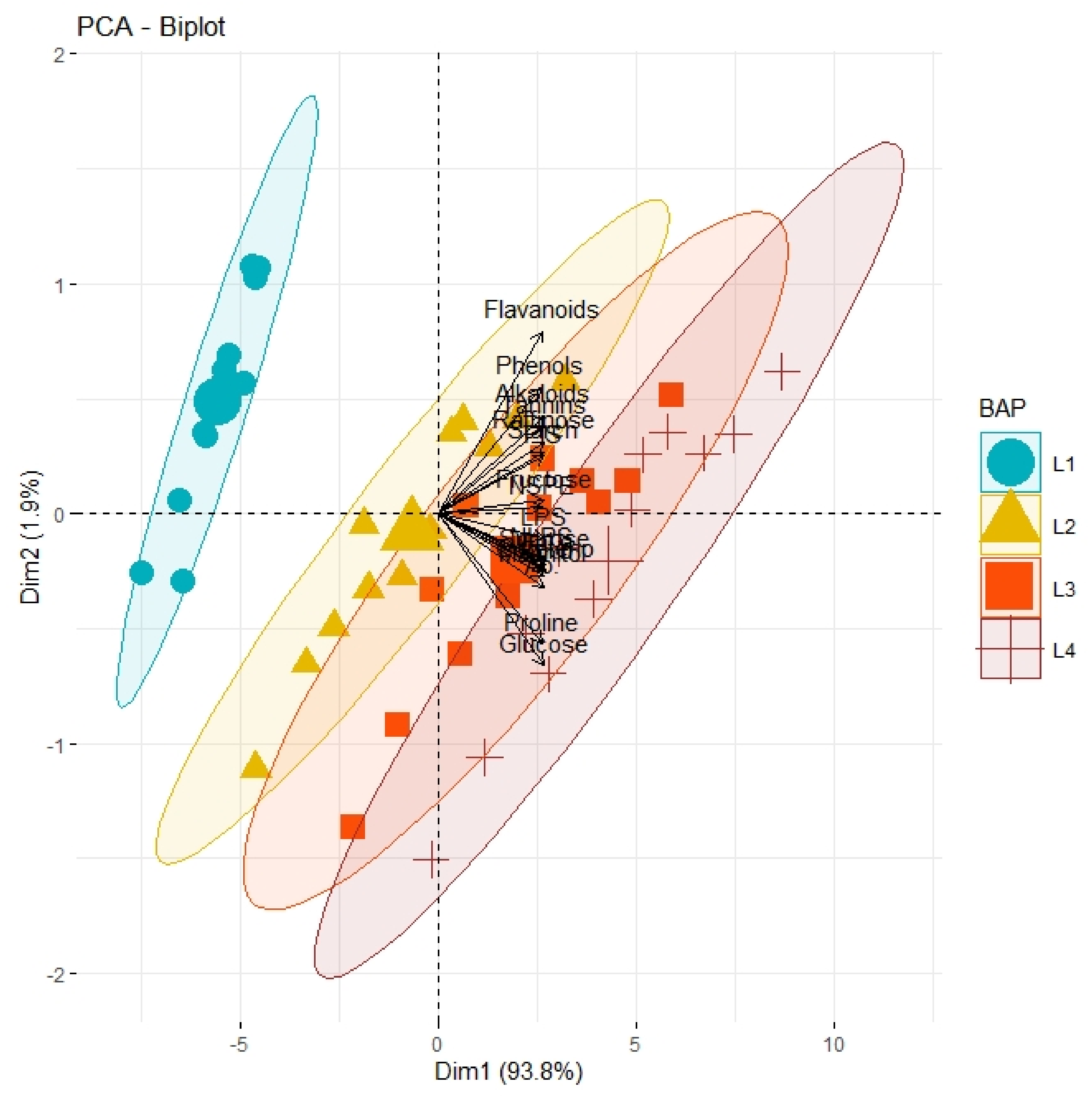
3.4. Correlation and Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peixe, A.; Raposo, A.; Lourenço, R.; Cardoso, H.; Macedo, E. Coconut water and BAP successfully replaced zeatin in olive (Olea europaea L.) micropropagation. Sci. Hortic. 2007, 113, 1–7. [Google Scholar] [CrossRef]
- Espinosa-Leal, C.A.; Puente-Garza, C.A.; García-Lara, S. In vitro plant tissue culture: Means for production of biological active compounds. Planta 2018, 248, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Lambardi, M.; Ozudogru, E.A.; Roncasaglia, R. In vitro propagation of olive (Olea europaea L.) by nodal segmentation of elongated shoots. In Protocols for Micropropagation of Selected Economically-Important Horticultural Plants; Humana Press: Totowa, NJ, USA, 2012; pp. 33–44. [Google Scholar]
- Benelli, C.; De Carlo, A. In vitro multiplication and growth improvement of Olea europaea L. cv Canino with temporary immersion system (Plantform™). 3 Biotech 2018, 8, 371. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Ahmad, T.; Abbasi, N.A.; Hafiz, I.A. Effect of different media and growth regulators on in vitro shoot proliferation of olive cultivar ‘Moraiolo’. Pak. J. Bot. 2009, 41, 783–795. [Google Scholar]
- Chaari-Rkhis, A.; Maalej, M.; Drira, N.; Standardi, A. Micropropagation of olive tree Olea europaea L. ‘Oueslati’. Turk. J. Agric. For. 2011, 35, 403–412. [Google Scholar]
- Revilla, M.A.; Pacheco, J.; Casares, A.; Rodriguez, R. In vitro reinvigoration of mature olive trees (Olea europaea L.) through micrografting. In Vitro Cell. Dev. Biol.-Plant 1996, 32, 257. [Google Scholar] [CrossRef]
- Rugini, E. In vitro propagation of some olive (Olea europaea L.) cultivars with different root ability, and medium development using analytical data from developing shoots and embryos. Sci. Hortic. 1984, 24, 123–134. [Google Scholar] [CrossRef]
- Hamooh, B.T.; Shah, Z.H. In vitro evaluation of shoot induction and proliferation protocol for olive cultivars by assessing morpho-physiologic effects of pre-cooling and growth regulators. Int. J. Biol. 2017, 11, 126–139. [Google Scholar]
- Ahmadi-Lahijani, M.J.; Kafi, M.; Nezami, A.; Nabati, J.; Erwin, J.E. ABA and BAP improve the accumulation of carbohydrates and alter carbon allocation in potato plants at elevated CO2. Physiol. Mol. Biol. Plants 2021, 27, 313–325. [Google Scholar] [CrossRef]
- Tahiri, A.; Destain, J.; Thonart, P.; Druart, P. In vitro model to study the biological properties of humic fractions from landfill leachate and leonardite during root elongation of Alnus glutinosa L. Gaertn and Betula pendula Roth. Plant Cell Tiss. Organ Cult. 2015, 122, 739–749. [Google Scholar] [CrossRef]
- Ferrara, g.; Loffredo, E.; Simeone, R.; Senesi, N. Evaluation of antimutagenic and desmutagenic effects of humic and fulvic acids on root tips of Vicia faba. Environ. Toxicol. 2000, 15, 513–517. [Google Scholar] [CrossRef]
- Ferrara, G.; Loffredo, E.; Senesi, N.; Marcos, R. Humic acids reduce the genotoxicity of mitomycin C in human lymphoblastoid cell line TK6. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2006, 603, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, G.; Loffredo, E.; Senesi, N. Anticlastogenic, antitoxic and sorption effects of humic substances on the mutagen maleic hydrazide tested in leguminous plants. Eur. J. Soil Sci. 2004, 55, 449–458. [Google Scholar] [CrossRef]
- Salem, J.; Hassanein, A.; El-Wakil, D.A.; Loutfy, N. Interaction between Growth Regulators Controls In Vitro Shoot Multiplication in Paulownia and Selection of NaCl-Tolerant Variants. Plants 2022, 11, 498. [Google Scholar] [CrossRef]
- Lebedev, V.; Arkaev, M.; Dremova, M.; Pozdniakov, I.; Shestibratov, K. Effects of Growth Regulators and Gelling Agents on Ex Vitro Rooting of Raspberry. Plants 2018, 8, 3. [Google Scholar] [CrossRef]
- Leva, A.; Petruccelli, R.; Rinaldi, L. Somaclonal Variation in Tissue Culture: A Case Study with Olive. In Recent Advances in Plant in vitro Culture; Leva, A., Rinaldi, L.M.R., Eds.; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Antonopoulou, C.; Dimassi, K.D.; Therios, I.T.; Hatzissavvidis, C. Does dikegulac affect in vitro shoot proliferation and hyperhydricity incidence in olive explants? Hortic. Sci. 2018, 45, 125–130. [Google Scholar] [CrossRef]
- Brunner, A.M.; Evans, L.M.; Hsu, C.Y.; Sheng, X. Vernalization and the chilling requirement to exit bud dormancy: Shared or separate regulation? Front. Plant Sci. 2014, 5, 732. [Google Scholar] [CrossRef]
- Pan, W.; Liang, J.; Sui, J.; Li, J.; Liu, C.; Xin, Y.; Zhang, Y.; Wang, S.; Zhao, Y.; Zhang, J.; et al. ABA and Bud Dormancy in Perennials: Current Knowledge and Future Perspective. Genes 2021, 12, 1635. [Google Scholar] [CrossRef]
- Liu, J.; Sherif, S.M. Hormonal orchestration of bud dormancy cycle in deciduous woody perennials. Front. Plant Sci. 2019, 10, 1136. [Google Scholar] [CrossRef]
- Alexandre, C.; Hennig, L. FLC-independent vernalization responses. Int. J. Plant Dev. Biol. 2007, 1, 202–211. [Google Scholar]
- Leida, C.; Conejero, A.; Arbona, V.; Gómez-Cadenas, A.; Llácer, G.; Badenes, M.L.; Ríos, G. Chilling-dependent release of seed and bud dormancy in peach associates to common changes in gene expression. PLoS ONE 2012, 10, 35777. [Google Scholar] [CrossRef] [PubMed]
- Dhami, N.; Cazzonelli, C.I. Prolonged cold exposure to Arabidopsis juvenile seedlings extends vegetative growth and increases the number of shoot branches. Plant Signal. Behav. 2020, 15, 1789320. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Saxena, P.K. Inhibition of phenylpropanoid biosynthesis in Artemisia annua L.: A novel approach to reduce oxidative browning in plant tissue culture. PLoS ONE 2013, 8, 76802. [Google Scholar] [CrossRef]
- Martini, A.N.; Papafotiou, M.; Vemmos, S.N. Season and explant origin affect phenolic content, browning of explants, and micropropagation of× Malosorbus florentina (Zucc.) Browicz. HortScience 2013, 48, 102–107. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, P.; Xue, J.; Xue, Y.; Wang, S.; Zhang, X. Advances in phenolic substances and their effects on browning in woody plant tissue culture. Acta Hortic. Sin. 2019, 46, 1645–1654. [Google Scholar]
- Mahmood, S.; Daur, I.; Al-Solaimani, S.G.; Ahmad, S.; Madkour, M.H.; Yasir, M.; Hirt, H.; Ali, S.; Ali, Z. Plant Growth Promoting Rhizobacteria and Silicon Synergistically Enhance Salinity Tolerance of Mung Bean. Front. Plant Sci. 2016, 7, 876. [Google Scholar] [CrossRef] [PubMed]
- Boussadia, O.; Steppe, K.; Zgallai, H.S.; Ben El-Hadj, S.; Braham, M.; Lemeur, R.; Van-Labeke, M.C. Effects of nitrogen deficiency on leaf photosynthesis, carbohydrate status and biomass production in two olive cultivars ‘Meski’ and ‘Koroneiki’. Sci. Hortic. 2010, 123, 336–342. [Google Scholar] [CrossRef]
- Chow, P.S.; Landhäusser, S.M. A method for routine measurements of total sugar and starch content in woody plant tissues. Tree Physiol. 2004, 24, 1129–1136. [Google Scholar] [CrossRef]
- Siddiqui, N.; Rauf, A.; Latif, A.; Mahmood, Z. Spectrophotometric determination of the total phenolic content, spectral and fluorescence study of the herbal Unani drug Gul-e-Zoofa (Nepeta bracteate Benth). J. Taibah Univ. Med. Sci. 2017, 12, 360–363. [Google Scholar] [CrossRef]
- Zhao, L.J.; Liu, W.; Xiong, S.H.; Tang, J.; Lou, Z.H.; Xie, M.X.; Xia, B.H.; Lin, L.M.; Liao, D.F. Determination of total flavonoids contents and antioxidant activity of Ginkgo biloba leaf by near-infrared reflectance method. Int. J. Anal. Chem. 2018, 2018, 8195784. [Google Scholar] [CrossRef]
- Fadda, A.; Mulas, M. Chemical changes during myrtle (Myrtus communis L.) fruit development and ripening. Sci. Hortic. 2010, 125, 477–485. [Google Scholar] [CrossRef]
- Li, L.; Long, W.; Wan, X.; Ding, Q.; Zhang, F.; Wan, D. Studies on quantitative determination of total alkaloids and berberine in five origins of crude medicine “Sankezhen”. J. Chromatogr. Sci. 2015, 53, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Zuccherelli, G.; Zuccherelli, S. In vitro propagation of fifty olive cultivars. Acta Hortic. 2002, 586, 931–934. [Google Scholar] [CrossRef]
- Guo, R.; Yuan, G.; Wang, Q. Effect of sucrose and mannitol on the accumulation of health-promoting compounds and the activity of metabolic enzymes in broccoli sprouts. Sci. Hortic. 2011, 128, 159–165. [Google Scholar] [CrossRef]
- Ejaz, B.; Sajid, Z.A.; Aftab, F. Effect of exogenous application of ascorbic acid on antioxidant enzyme activities, proline contents, and growth parameters of Saccharum spp., hybrid cv. HSF-under salt stress. Turk. J. Biol. 2012, 36, 630–640. [Google Scholar] [CrossRef]
- Aslam, M.; Sultana, B.; Anwar, F.; Munir, H. Foliar spray of selected plant growth regulators affected the biochemical and antioxidant attributes of spinach in a field experiment. Turk. J. Agric. For. 2016, 40, 136–145. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Santos-Sánchez, N.F.; Salas-Coronado, R.; Hernández-Carlos, B.; Villanueva-Cañongo, C. Shikimic Acid Pathway in Biosynthesis of Phenolic Compounds. In Plant Physiological Aspects of Phenolic Compounds; Soto-Hernández, M., García-Mateos, R., Palma-Tenango, M., Eds.; IntechOpen: London, UK, 2019. [Google Scholar]
- Ghasemzadeh, A.; Jaafar, H.Z.E.; Rahmat, A. Elevated carbon dioxide increases contents of flavonoids and phenolic compounds, and antioxidant activities in Malaysian young ginger (Zingiber officinale Roscoe.) varieties. Molecules 2010, 15, 7907–7922. [Google Scholar] [CrossRef]
- El Sabagh, A.; Islam, M.S.; Hossain, A.; Mubeen, M.; Iqbal, M.A.; Waleed, M.; Reginato, M.; Battaglia, M.; Ahmed, S.; Rehman, A.; et al. Phytohormones as growth regulators during abiotic stress tolerance in plants. Front. Agron. 2022, 8, 68. [Google Scholar] [CrossRef]



| Treatments | LPS (cm) | PIS | NLPS | NSPE |
|---|---|---|---|---|
| Pre-cooling (T) (Hours) | ||||
| T1 (0 h) | 1.97 ± 0.11 b | 70.02 ± 2.00 c | 3.71 ± 0.15 c | 2.045 ± 0.10 c |
| T2 (24 h) | 2.27 ± 0.13 ab | 78.05 ± 1.87 b | 3.93 ± 0.17 b | 2.56 ± 0.13 b |
| T3 (48 h) | 2.55 ± 0.15 a | 83.06 ± 1.62 a | 4.28 ± 0.20 a | 4.00 ± 0.19 a |
| LSD | 0.35 | 1.96 | 0.13 | 0.14 |
| Varieties (V) | ||||
| V1 (Leccino) | 1.98 ± 0.15 c | 65 ± 2.23 d | 3.58 ± 0.18 d | 2.05 ± 0.13 d |
| V2 (Gemlik) | 2.26 ± 0.14 b | 70 ± 2.49 c | 3.88 ± 0.19 c | 2.43 ± 0.18 c |
| V3 (Morailo) | 2.34 ± 0.15 b | 75 ± 2.74 b | 4.09 ± 0.20 b | 3.60 ± 0.20 b |
| V4 (Arbosana) | 3.00 ± 0.17 a | 85 ± 3.13 a | 5.00 ± 0.23 a | 4.00 ± 0.21 a |
| LSD | 0.10 | 2.70 | 0.08 | 0.08 |
| BAP (LB) (mgL−1) | ||||
| L1 (0) | 1.05 ± 0.034 d | 68 ± 0.073 d | 2.48 ± 0.045 d | 2.5 ± 0.021 d |
| L2(0.5) | 2.03 ± 0.083 c | 75 ± 0.80 c | 3.40 ± 0.09 c | 3 ± 0.093 c |
| L3 (1.5) | 2.32 ± 0.088 b | 79 ± 1.93 b | 4.18 ± 0.10 b | 3.25 ± 0.10 b |
| L4(2.5) | 2.55 ± 0.083 a | 84 ± 1.87 a | 4.58 ± 0.98 a | 4.01 ± 0.14 a |
| LSD | 0.08 | 1.30 | 0.11 | 0.08 |
| Significance | ||||
| T | * | * | ** | * |
| V | ** | ** | ** | ** |
| LB | ** | ** | ** | ** |
| T × V | ns | ns | ns | ns |
| T × LB | * | * | * | * |
| V × LB | * | * | * | * |
| T × V × LB | ns | ns | ns | ns |
| Treatments | Total Chlorophyll (μg cm−2) | Carotenoids (μg cm−2) | ACO2 (μmol m−2s−1) | Proline (μg g−1 FW) |
|---|---|---|---|---|
| Pre-cooling (T) (Hours) | ||||
| T1 (0 h) | 41.83 ± 2.52 b | 4.53 ± 0.20 c | 6.93 ± 0.37 c | 42.00 ± 2.10 a |
| T2 (24 h) | 47.96 ± 3.07 a | 5.05 ± 0.25 b | 7.47 ± 0.39 b | 39.70 ± 1.76 b |
| T3 (48 h) | 51.55 ± 3.45 a | 5.52 ± 0.32 a | 8.09 ± 0.42 a | 38.87 ± 1.76 b |
| LSD | 5.12 | 0.38 | 0.32 | 1.91 |
| Varieties (V) | ||||
| V1 (Leccino) | 42 ± 2.00 c | 4.81 ± 0.28 c | 6.59 ± 0.40 c | 41.32 ± 2.35 a |
| V2 (Gemlik) | 46.57 ± 3.37 b | 4.93 ± 0.35 a | 7.41 ± 0.44 b | 39.74 ± 2.29 b |
| V3 (Morailo) | 48.53 ± 3.76 ab | 5.09 ± 0.30 b | 7.76 ± 0.48 ab | 41.00 ± 2.14 a |
| V4 (Arbosana) | 50.37 ± 4.11 a | 5.29 ± 0.28 c | 7.93 ± 0.50 a | 38.70 ± 2.08 c |
| LSD | 2.74 | 0.11 | 0.36 | 0.59 |
| BAP (LB) (mgL−1) | ||||
| L1 (0) | 19.70 ± 0.26 d | 2.71 ± 0.050 d | 3.82 ± 0.10 d | 48.66 ± 0.63 a |
| L2(0.5) | 27.62 ± 0.70 c | 4.17 ± 0.12 c | 5.82 ± 0.12 c | 46.62 ± 0.62 b |
| L3 (1.5) | 32.53 ± 0.83 b | 4.84 ± 0.11 b | 6.40 ± 0.12 b | 43.37 ± 0.61 c |
| L4(2.5) | 37.68 ± 0.77 a | 5.44 ± 0.13 a | 6.95 ± 0.12 a | 23.41 ± 0.20 d |
| LSD | 0.60 | 0.14 | 0.17 | 1.24 |
| Significance | ||||
| T | * | * | ** | * |
| V | ** | ** | * | ** |
| L | ** | ** | ** | ** |
| T × V | ns | ns | ns | ns |
| T × LB | * | * | ns | * |
| V × LB | * | * | ns | ns |
| T × V × LB | ns | ns | ns | ns |
| Treatments | Starch mg g−1 DW | Glucose mg g−1 DW | Fructose (mg g−1 DW) | Sucrose (mg g−1 DW) |
|---|---|---|---|---|
| Pre-cooling (T) (Hours) | ||||
| T1 (0 h) | 0.33 ± 0.05 b | 1.04 ± 0.10 c | 0.029 ± 0.002 c | 0.29 ± 0.03 c |
| T2 (24 h) | 0.53 ± 0.06 ab | 1.19 ± 0.10 b | 0.032 ± 0.003 b | 0.41 ± 0.04 b |
| T3 (48 h) | 0.62 ± 0.06 a | 1.39 ± 0.12 a | 0.035 ± 0.004 a | 0.51 ± 0.05 a |
| LSD | 0.21 | 0.08 | 0.002 | 0.07 |
| Varieties (V) | ||||
| V1 (Leccino) | 0.42 ± 0.06 b | 1.07 ± 0.11 c | 0.024 ± 0.003 b | 0.32 ± 0.04 d |
| V2 (Gemlik) | 0.41 ± 0.06 b | 1.13 ± 0.11 bc | 0.027 ± 0.004 b | 0.38 ± 0.05 c |
| V3 (Morailo) | 0.47 ± 0.07 b | 1.42 ± 0.11 a | 0.028 ± 0.004 b | 0.42 ± 0.05 b |
| V4 (Arbosana) | 0.68 ± 0.08 a | 1.20 ± 0.15 b | 0.039 ± 0.005 a | 0.48 ± 0.06 a |
| LSD | 0.11 | 0.08 | 0.08 | 0.03 |
| BAP (LB) (mgL−1) | ||||
| L1 (0 ) | 0.09 ± 0.01 d | 0.21 ± 0.02 d | 0.007 ± 0.0003 d | 0.05 ± 0.004 d |
| L2 (0.5 ) | 0.19 ± 0.01 c | 0.55 ± 0.01 c | 0.020 ± 0.0014 c | 0.19 ± 0.014 c |
| L3 (1.5) | 0.32 ± 0.02 a | 0.67 ± 0.02 b | 0.031 ± 0.0020 b | 0.26 ± 0.009 b |
| L4 (2.5 ) | 0.26 ± 0.01 b | 0.83 ± 0.02 a | 0.040 ± 0.0024 a | 0.31 ± 0.010 a |
| LSD | 0.02 | 0.02 | 0.002 | 0.006 |
| Significance | ||||
| T | ** | * | ** | * |
| V | ** | ** | ** | * |
| LB | ** | ** | ** | ** |
| T × V | ns | ns | ns | ns |
| T × LB | ns | * | * | * |
| V × LB | ns | ns | * | * |
| T × V × LB | ns | ns | ns | ns |
| Starch | 0.91 *** | 0.95 *** | 0.94 *** | 0.94 *** | 0.93 *** | 0.92 *** | 0.88 *** | 0.95 *** | 0.94 *** | 0.91 *** | 0.92 *** | 0.94 *** | 0.93 *** | 0.92 *** | 0.94 *** |
| Glucose | 0.92 *** | 0.97 *** | 0.97 *** | 0.98 *** | 0.96 *** | 0.97 *** | 0.90 *** | 0.85 *** | 0.88 *** | 0.91 *** | 0.91 *** | 0.94 *** | 0.93 *** | 0.90 *** | |
| Fructose | 0.93 *** | 0.96 *** | 0.92 *** | 0.95 *** | 0.90 *** | 0.92 *** | 0.92 *** | 0.89 *** | 0.92 *** | 0.93 *** | 0.92 *** | 0.93 *** | 0.94 *** | ||
| Sucrose | 0.98 *** | 0.98 *** | 0.97 *** | 0.97 *** | 0.95 *** | 0.91 *** | 0.92 *** | 0.95 *** | 0.95 *** | 0.96 *** | 0.93 *** | 0.92 *** | |||
| Chlorophyll | 0.96 *** | 0.98 *** | 0.96 *** | 0.94 *** | 0.92 *** | 0.92 *** | 0.96 *** | 0.95 *** | 0.95 *** | 0.95 *** | 0.93 *** | ||||
| ACO2 | 0.96 *** | 0.96 *** | 0.93 *** | 0.89 *** | 0.92 *** | 0.93 *** | 0.95 *** | 0.97 *** | 0.94 *** | 0.93 *** | |||||
| Carotenoids | 0.96 *** | 0.94 *** | 0.91 *** | 0.91 *** | 0.94 *** | 0.95 *** | 0.96 *** | 0.95 *** | 0.94 *** | ||||||
| Proline | 0.90 *** | 0.85 *** | 0.87 *** | 0.91 *** | 0.91 *** | 0.93 *** | 0.91 *** | 0.90 *** | |||||||
| Alkaloids | 0.97 *** | 0.93 *** | 0.95 *** | 0.97 *** | 0.95 *** | 0.92 *** | 0.93 *** | ||||||||
| Flavonoids | 0.94 *** | 0.96 *** | 0.94 *** | 0.90 *** | 0.89 *** | 0.91 *** | |||||||||
| Phenols | 0.96 *** | 0.93 *** | 0.90 *** | 0.89 *** | 0.87 *** | ||||||||||
| Tannins | 0.95 *** | 0.91 *** | 0.90 *** | 0.89 *** | |||||||||||
| PIS | 0.97 *** | 0.93 *** | 0.96 *** | ||||||||||||
| LPS | 0.95 *** | 0.96 *** | |||||||||||||
| NLPS | 0.94 *** | ||||||||||||||
| NSPE |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatoon, S.; Liu, W.; Ding, C.-b.; Liu, X.; Zheng, Y.; Zhang, Y.; Chen, X.; Rauf, M.; Alghabari, F.; Shah, Z.H. In Vitro Evaluation of the Effects of BAP Concentration and Pre-Cooling Treatments on Morphological, Physiological, and Biochemical Traits of Different Olive (Olea euorpea L.) Cultivars. Horticulturae 2022, 8, 1108. https://doi.org/10.3390/horticulturae8121108
Khatoon S, Liu W, Ding C-b, Liu X, Zheng Y, Zhang Y, Chen X, Rauf M, Alghabari F, Shah ZH. In Vitro Evaluation of the Effects of BAP Concentration and Pre-Cooling Treatments on Morphological, Physiological, and Biochemical Traits of Different Olive (Olea euorpea L.) Cultivars. Horticulturae. 2022; 8(12):1108. https://doi.org/10.3390/horticulturae8121108
Chicago/Turabian StyleKhatoon, Sadia, Wencong Liu, Chuan-bo Ding, Xinglong Liu, Yinan Zheng, Yue Zhang, Xueyan Chen, Muhammad Rauf, Fahad Alghabari, and Zahid Hussain Shah. 2022. "In Vitro Evaluation of the Effects of BAP Concentration and Pre-Cooling Treatments on Morphological, Physiological, and Biochemical Traits of Different Olive (Olea euorpea L.) Cultivars" Horticulturae 8, no. 12: 1108. https://doi.org/10.3390/horticulturae8121108
APA StyleKhatoon, S., Liu, W., Ding, C.-b., Liu, X., Zheng, Y., Zhang, Y., Chen, X., Rauf, M., Alghabari, F., & Shah, Z. H. (2022). In Vitro Evaluation of the Effects of BAP Concentration and Pre-Cooling Treatments on Morphological, Physiological, and Biochemical Traits of Different Olive (Olea euorpea L.) Cultivars. Horticulturae, 8(12), 1108. https://doi.org/10.3390/horticulturae8121108