
Genetic and Cellular Basis of Resistance to Black Rot Caused by Xanthomonas campestris pv. campestris in Brassica rapa


 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Inoculation Test
2.3. Resistance Evaluation
2.4. Genetic Analysis
2.5. Scanning Electron Microscopy
2.6. Transmission Electron Microscopy
2.7. Data Analysis
3. Results
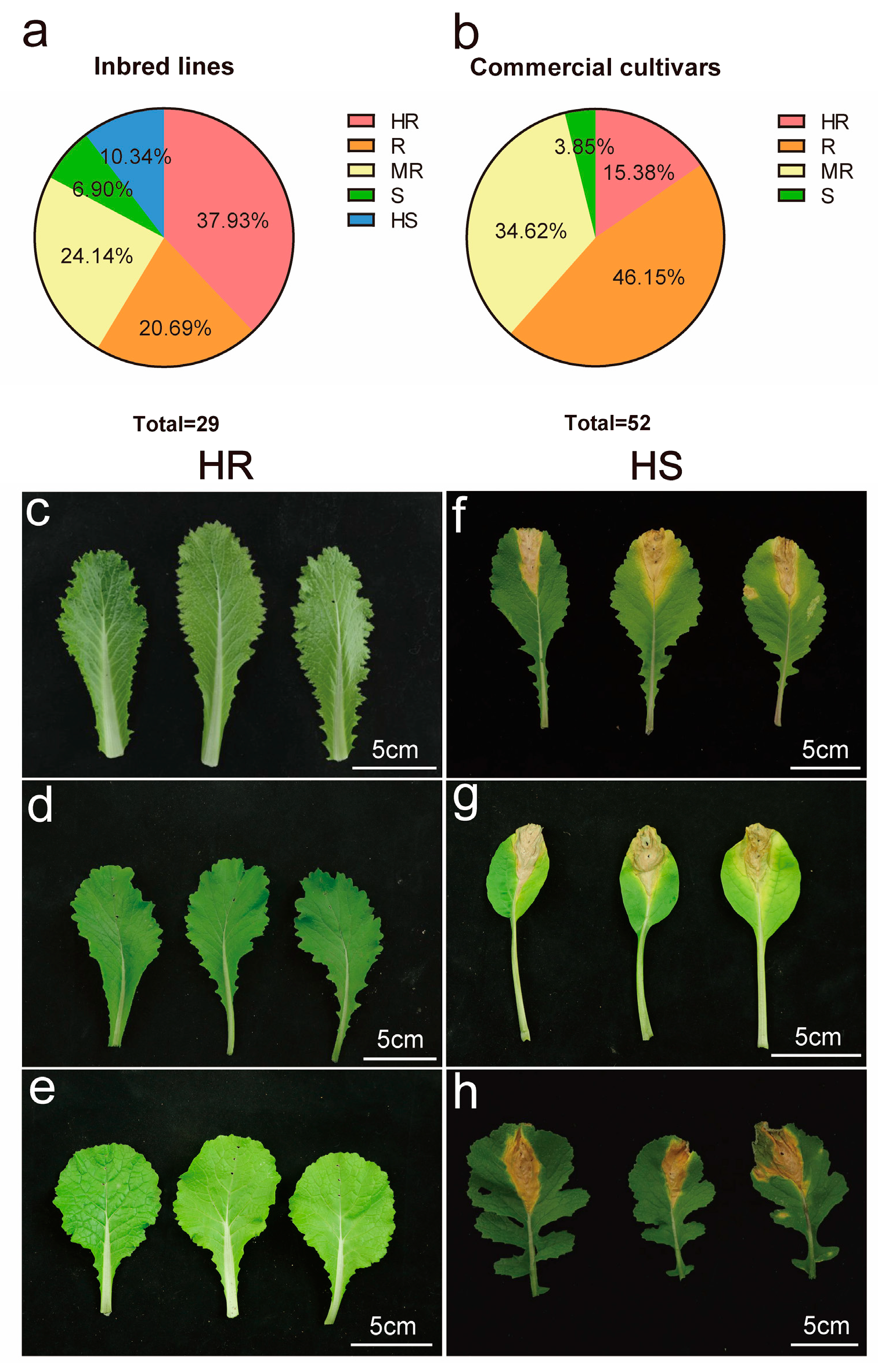
3.1. Screening of Resistant Sources to Black Rot in B. rapa
3.2. Genetic Analysis of Resistance to Black Rot in B. rapa
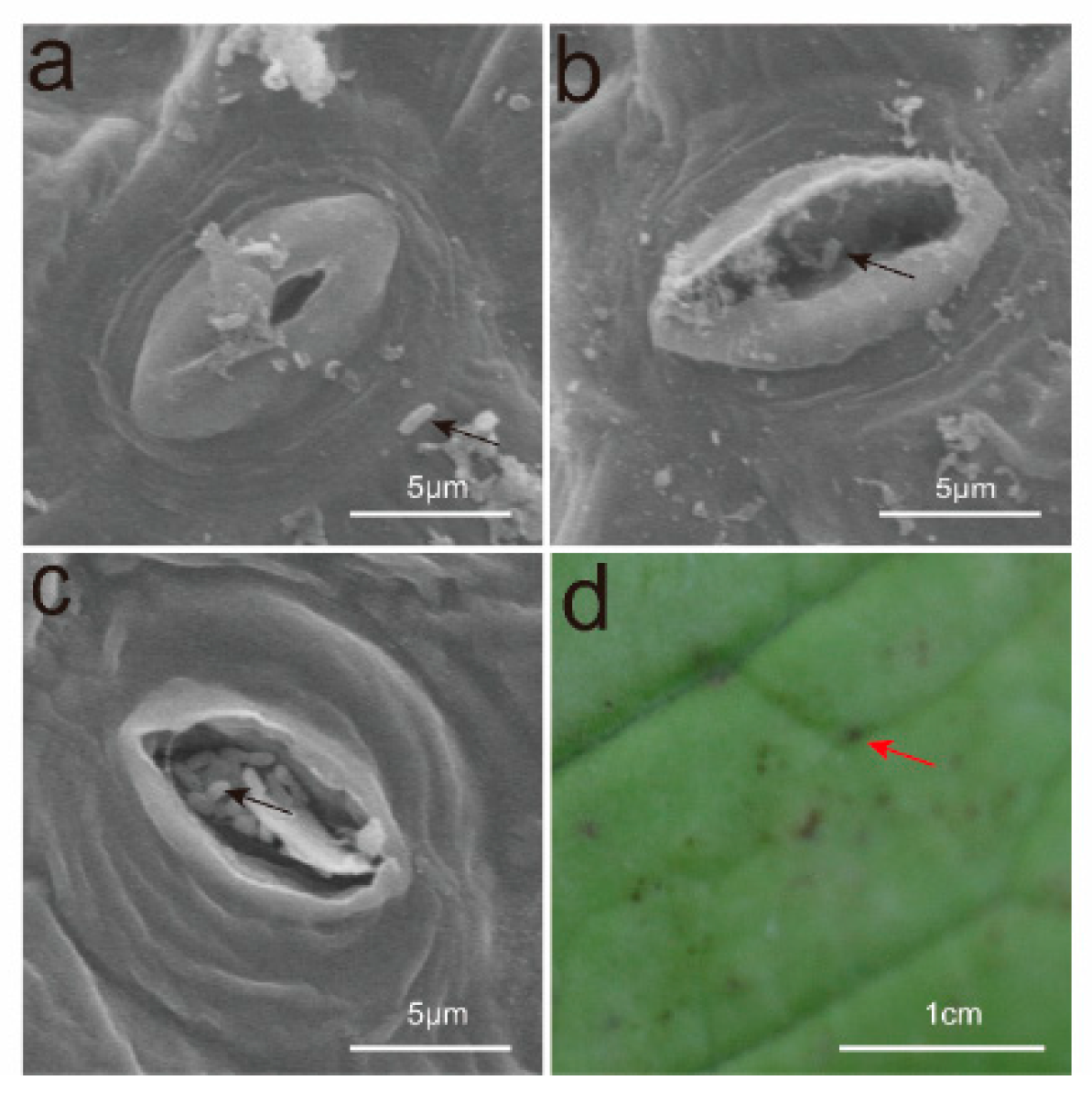
3.3. Observations on the Invasion Pathways of Xcc
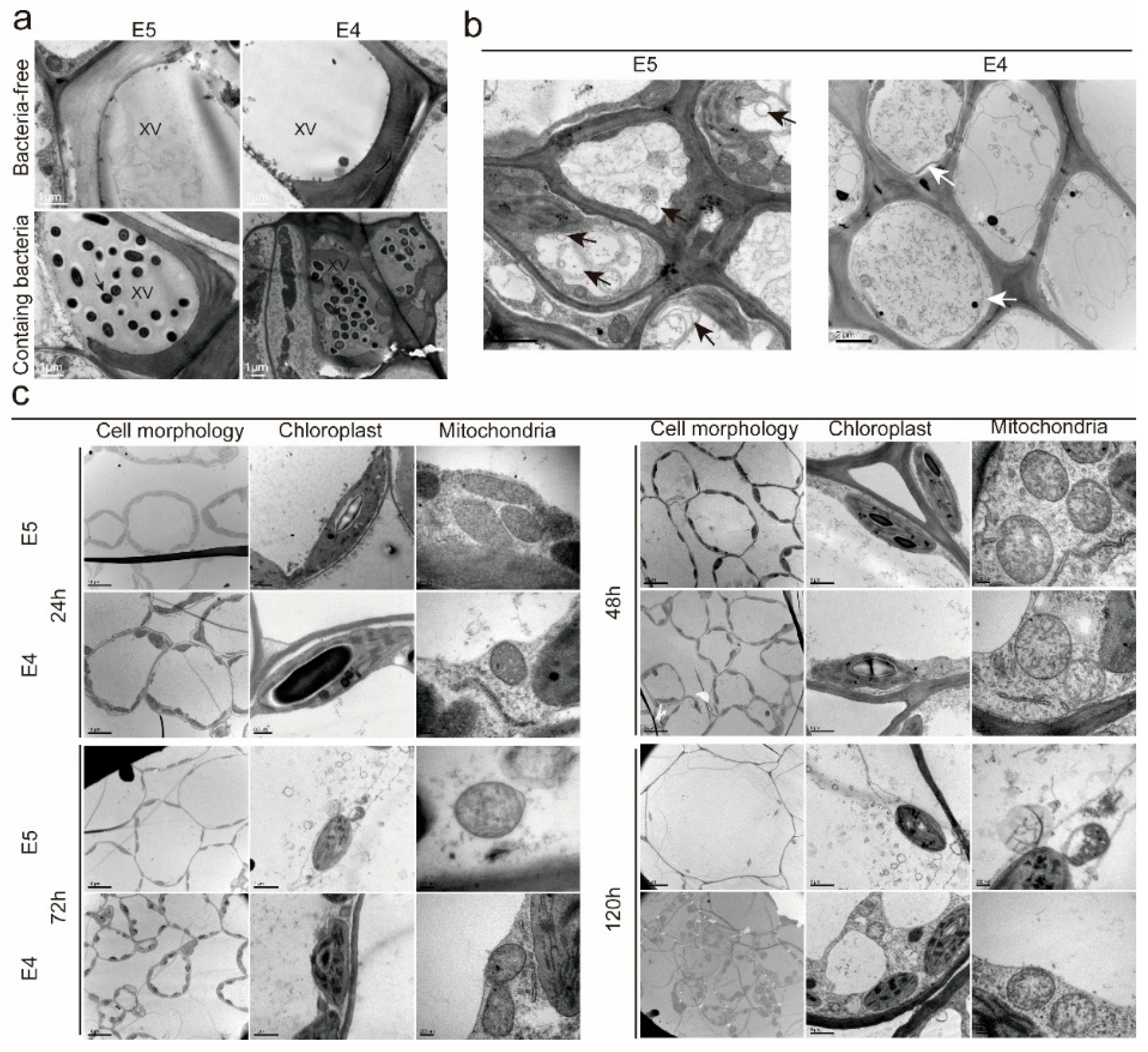
3.4. The Ultrastructure of Resistant and Susceptible B. rapa Leaves After Xcc Inoculation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lv, H.; Fang, Z.; Yang, L.; Zhang, Y.; Wang, Y. An Update on the Arsenal: Mining Resistance Genes for Disease Management of Brassica Crops in the Genomic Era. Hortic. Res. 2020, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Yuan, K.; Zhao, X.; Sun, W.; Yang, L.; Zhang, Y.; Wang, Y.; Ji, J.; Han, F.; Fang, Z.; Lv, H. Map-Based Cloning and CRISPR/Cas9-Based Editing Uncover BoNA1 as the Causal Gene for the No-Anthocyanin-Accumulation Phenotype in Curly Kale (Brassica oleracea var. sabellica). Hortic. Res. 2023, 10, uhad133. [Google Scholar] [CrossRef]
- Williams, P.H. Black Rot : A Continuing Threat to World Crucifers. Plant Dis. 1980, 64, 736–742. [Google Scholar] [CrossRef]
- Vicente, J.G.; Holub, E.B. Xanthomonas campestris pv. campestris (Cause of Black Rot of Crucifers) in the Genomic Era Is Still a Worldwide Threat to Brassica Crops. Mol. Plant Pathol. 2013, 14, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Izzah, N.K.; Jayakodi, M.; Perumal, S.; Joh, H.J.; Lee, H.J.; Lee, S.-C.; Park, J.Y.; Yang, K.-W.; Nou, I.-S.; et al. Genome-Wide SNP Identification and QTL Mapping for Black Rot Resistance in Cabbage. BMC Plant Biol. 2015, 15, 32. [Google Scholar] [CrossRef]
- Ma, Y.; Meng, Y.; Wang, Y.; Xu, L.; Chen, Y.; Yuan, Y.; Zhang, X.; Wang, L.; Wei, X.; Li, B.; et al. Research Progress on Clubroot Disease in Brassicaceae Crops—Advances and Perspectives. Veg. Res. 2024, 4, e022. [Google Scholar] [CrossRef]
- Bain, D.C. Reaction of Brassica Seedlings to Blackrot. Phytopathology 1952, 42, 497–500. [Google Scholar]
- Silva, D.; Andre, L.; Joara, S.; Eizanilda, R.; Timothy, B. Screening Cabbage Cultivars for Resistance to Black Rot Under Field Conditions. HortTechnology 2020, 30, 448–455. [Google Scholar] [CrossRef]
- Liu, Y.; Li, D.; Yang, N.; Zhu, X.; Han, K.; Gu, R.; Bai, J.; Wang, A.; Zhang, Y. Genome-Wide Identification and Analysis of CC-NBS-LRR Family in Response to Downy Mildew and Black Rot in Chinese Cabbage. Int. J. Mol. Sci. 2021, 22, 4266. [Google Scholar] [CrossRef]
- Kopta, T.; Penazova, E.; Jurica, M.; Pokluda, R. Evaluation of the Potential Yield and Primary Symptoms of Xanthomonas campestris pv. campestris Infection in Asian Vegetables Grown in the Czech Republic. Acta Agrobot. 2018, 4, 71. [Google Scholar] [CrossRef]
- Guo, H.; Dickson, M.H.; Hunter, J.E. Brassica napus Sources of Resistance to Black Rot in Crucifers and Inheritance of Resistance. HortScience 1991, 26, 1545–1547. [Google Scholar] [CrossRef]
- Griffiths, P.D.; Marek, L.F.; Robertson, L.D. Identification of Crucifer Accessions from the NC-7 and NE-9 Plant Introduction Collections That Are Resistant to Black Rot (Xanthomonas campestris pv. campestris) Races 1 and 4. Hortence Publ. Am. Soc. Hortic. Ence 2009, 44, 77–88. [Google Scholar] [CrossRef]
- Elston, R.C. The Genetic Analysis of Quantitative Trait Differences Between Two Homozygous Lines. Genetics 1984, 108, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Gai, J.; Wang, Y.; Xiaolei, W.U.; Chen, S. A Comparative Study on Segregation Analysis and QTL Mapping of Quantitative Traits in Plants—With a Case in Soybean. Front. Agric. China 2007, 1, 1–7. [Google Scholar] [CrossRef]
- Khan, M.I.; Khattak, G.S.S.; Khan, A.J.; Subhan, F.; Ali, A. Genetic Control of Flag Leaf Area in Wheat (Triticum aestivum) Crosses. Afr. J. Agric. Res. 2012, 7, 3978–3990. [Google Scholar] [CrossRef]
- Li, G.; Chen, L.; Zhang, G.; He, X.; Zhi, H.; Zhang, Y. Mixed Major-Gene Plus Polygenes Inheritance Analysis for Resistance in Soybean to Bean Pyralid (Lamprosema indicata Fabricius). Soybean Sci. 2008, 27, 33. [Google Scholar]
- Zhang, S.-Q.; Gu, X.-F.; Zhang, S.-P.; Zou, Z.-R. Inheritance of Downy Mildew Resistance in Cucumber (Cucumis sativus L.). Acta Bot. Boreali-Occident. Sin. 2010, 26, 2416–2420. [Google Scholar]
- Zheng, W.-J.; Liu, Z.-H.; Zhao, J.-M.; Chen, W.-F. Genetic Analysis of Stripe Disease Resistance in Rice Restorer Line C224 Using Major Gene Plus Polygene Mixed Effect Model. Rice Sci. 2012, 19, 202–206. [Google Scholar] [CrossRef]
- Bain, D. Resistance of Cabbage to Black Rot. Phytopathology 1955, 45, 35–37. [Google Scholar]
- Williams, P.H. Inheritance of Resistance in Cabbage to Black Rot. Phytopathology 1971, 62, 247–252. [Google Scholar] [CrossRef]
- Singh, D.; Parkash, C.; Dey, R.; Bhatia, K.; Raj, G. Inter Specific Hybridization (Brassica carinata × Brassica oleracea) for Introgression of Black Rot Resistance Genes into Indian Cauliflower (B. oleracea var. botrytis L.). Euphytica Int. J. Plant Breed. 2015, 204, 149–162. [Google Scholar]
- Sharma, B.B.; Kalia, P.; Yadava, D.K.; Singh, D.; Sharma, T.R. Genetics and Molecular Mapping of Black Rot Resistance Locus Xca1bc on Chromosome B-7 in Ethiopian Mustard (Brassica carinata A. Braun). PLoS ONE 2016, 11, e0152290. [Google Scholar] [CrossRef] [PubMed]
- Saile, E.; Mcgarvey, J.A.; Schell, M.A.; Denny, T.P. Role of Extracellular Polysaccharide and Endoglucanase in Root Invasion and Colonization of Tomato Plants by Ralstonia Solanacearum. Phytopathology 1997, 87, 1264. [Google Scholar] [CrossRef] [PubMed]
- Mcgarvey, J.A.; Denny, T.P.; Schell, M.A. Spatial-Temporal and Quantitative Analysis of Growth and EPS I Production by Ralstonia Solanacearum in Resistant and Susceptible Tomato Cultivars. Phytopathology 1999, 89, 1233. [Google Scholar] [CrossRef]
- Ayliffe, M.; Jin, Y.; Kang, Z.; Persson, M.; Steffenson, B.; Wang, S.; Leung, H. Determining the Basis of Nonhost Resistance in Rice to Cereal Rusts. Euphytica 2011, 179, 33–40. [Google Scholar] [CrossRef]
- Heath, M.C. Nonhost Resistance and Nonspecific Plant Defenses. Curr. Opin. Plant Biol. 2000, 3, 315–319. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Liu, L.; Liu, Q.; Bi, Z.; Yu, N.; Cheng, S.; Cao, L. Fine Mapping of the Lesion Mimic and Early Senescence 1 (Lmes1) in Rice (Oryza sativa). Plant Physiol. Biochem. Ppb 2014, 80, 300–307. [Google Scholar] [CrossRef]
- Soylu, S.; Brown, I.; Mansfield, J.W. Cellular Reactions in Arabidopsis Following Challenge by Strains of Pseudomonas syringae: From Basal Resistance to Compatibility. Physiol. Mol. Plant Pathol. 2005, 66, 232–243. [Google Scholar] [CrossRef]
- Yang, Y.; Xie, L.; Yan, C.Q.; Wang, X.M.; Yu, C.L.; Cheng, X.Y.; Cheng, Y.; Chen, J.P. Xylem Secondary Cell-Wall Thickening Involved in Defense Responses of Oryza Meyerianato Xanthomonas oryzae pv. oryzae. Acta Phytopathol. Sin. 2012, 42, 505–514. [Google Scholar]
- Chen, W.; Tang, L.; Li, Q.; Cai, Y.; Ahmad, S.; Wang, Y.; Tang, S.; Guo, N.; Wei, X.; Tang, S. YGL3 Encoding an IPP and DMAPP Synthase Interacts with OsPIL11 to Regulate Chloroplast Development in Rice. Rice 2024, 17, 1–16. [Google Scholar] [CrossRef]
- Tian, R.; Tian, Y.; Dang, Q.; Zhang, H.; Huang, L. Vascular Network-Mediated Systemic Spread of Pseudomonas syringae pv. actinidiae Causes the Bacterial Canker of Kiwifruit. Hortic. Plant J. 2024. [Google Scholar] [CrossRef]
- Taylor, J.D.; Conway, J.; Roberts, S.J.; Astley, D.; Vicente, J.G. Sources and Origin of Resistance to Xanthomonas campestris pv. campestris in Brassica Genomes. Phytopathology 2002, 92, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Y.; Du, Y.; Ren, W.; Li, H.; Sun, W.; Ge, C.; Zhang, Y. SEA v2.0: An R Software Package for Mixed Major Genes plus Polygenes Inheritance Analysis of Quantitative Traits. ACTA Agron. Sin. 2022, 48, 1416–1424. [Google Scholar] [CrossRef]
- Akaike, H. A New Look at the Statistical Model Identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Siedow, J.N.; Rhoads, D.M.; Ward, G.C.; Levings, C.S. The Relationship Between the Mitochondrial Gene T-Urf13 and Fungal Pathotoxin Sensitivity in Maize. Biochim. Biophys. Acta 1995, 1271, 235–240. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, H.; Wang, J.; Lv, J.; Xie, B.; Luo, S.; Wang, S.; Zhang, B.; Li, Z.; Yue, Z.; et al. Physical, Chemical, and Biological Control of Black Rot of Brassicaceae Vegetables: A Review. Front. Microbiol. 2022, 13, 1023826. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Bischoff-Schaefer, M.; Bluemel, S.; Dachbrodt-Saaydeh, S.; Dreux, L.; Jansen, J.-P.; Kiss, J.; Köhl, J.; Kudsk, P.; Malausa, T.; et al. Identifying Obstacles and Ranking Common Biological Control Research Priorities for Europe to Manage Most Economically Important Pests in Arable, Vegetable and Perennial Crops. Pest. Manag. Sci. 2017, 73, 14–21. [Google Scholar] [CrossRef]
- Lema, M.; Cartea, M.E.; Francisco, M.; Velasco, P.; Soengas, P. Screening for Resistance to Black Rot in a Spanish Collection of Brassica rapa. Plant Breed. 2015, 134, 551–556. [Google Scholar] [CrossRef]
- Kong, C.; Chen, G.; Yang, L.; Zhuang, M.; Zhang, Y.; Wang, Y.; Ji, J.; Fang, Z.; Lv, H. Germplasm Screening and Inheritance Analysis of Resistance to Cabbage Black Rot in a Worldwide Collection of Cabbage (Brassica oleracea var. capitata) Resources. Sci. Hortic. 2021, 288, 110234. [Google Scholar] [CrossRef]
- Cruz, J.; Tenreiro, R.; Cruz, L. Assessment of Diversity of Xanthomonas campestris Pathovars Affecting Cruciferous Plants in Portugal and Disclosure of Two Novel X. campestris pv. campestris Races. J. Plant Pathol. 2017, 99, 403–414. [Google Scholar] [CrossRef]
- Mao, S.; Kim, Y.-H.; Sahu, N.; Kim, S.-W.; Kim, H.-T.; Watanabe, M.; Park, J.-I. Molecular Marker Development for Specific Amplification of Xanthomonas campestris pv. campestris Race 8 Causing Black Rot Disease in Brassica Crops. J. Gen. Plant Pathol. 2025, 91, 31–40. [Google Scholar] [CrossRef]
- He, Z.; Ji, R.; Havlickova, L.; Wang, L.; Bancroft, I. Genome Structural Evolution in Brassica Crops. Nat. Plants 2021, 7, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Holland, J.B. Genetic Architecture of Complex Traits in Plants. Curr. Opin. Plant Biol. 2007, 10, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Yan, L.; Ge, C.; Li, J.; Dai, J.; Ding, H. Genetic Mapping of Quantitative Trait Loci Conferring Resistance to Race 4 of Xanthomonas campestris pv. campestris in Cabbage (Brassica oleracea var. capitata). Physiol. Mol. Plant Pathol. 2025, 138, 102660. [Google Scholar] [CrossRef]
- Sharpe, J.A.; Robson, P.J.; Taylor, M.J.; Lydiate, D.; Parkin, I.; Vicente, G. Inheritance of Race-Specific Resistance to Xanthomonas campestris pv. campestris in Brassica Genomes. Phytopathology 2002, 92, 1134–1141. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The Plant Immune System. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Saha, P.; Kalia, P.; Sharma, M.; Singh, D. New Source of Black Rot Disease Resistance in Brassica oleracea and Genetic Analysis of Resistance. Euphytica 2016, 207, 35–48. [Google Scholar] [CrossRef]
- Soengas, P.; Hand, P.; Vicente, J.G.; Pole, J.M.; Pink, D.A.C. Identification of Quantitative Trait Loci for Resistance to Xanthomonas campestris pv. campestris in Brassica Rapa. Theor. Appl. Genet. 2007, 114, 637–645. [Google Scholar] [CrossRef]
- Balint-Kurti, P. The Plant Hypersensitive Response: Concepts, Control and Consequences. Mol. Plant Pathol. 2019, 20, 1163–1178. [Google Scholar] [CrossRef]
- Kroemer, G.; Galluzzi, L.; Vandenabeele, P.; Abrams, J.; Alnemri, E.S.; Baehrecke, E.H.; Blagosklonny, M.V.; El-Deiry, W.S.; Golstein, P.; Green, D.R.; et al. Classification of Cell Death: Recommendations of the Nomenclature Committee on Cell Death 2009. Cell Death Differ. 2009, 16, 3–11. [Google Scholar] [CrossRef]
- Zhou, B.; Zeng, L. Immunity-Associated Programmed Cell Death as a Tool for the Identification of Genes Essential for Plant Innate Immunity. Plant Program. Cell Death Methods Protoc. 2018, 1743, 51–63. [Google Scholar] [CrossRef]
- Kabbage, M.; Kessens, R.; Bartholomay, L.C.; Williams, B. The Life and Death of a Plant Cell. Annu. Rev. Plant Biol. 2017, 68, 375–404. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.S.; Jang, E.; Kim, J.; Kim, S.H.; Lee, M.-H.; Nam, M.H.; Tobimatsu, Y.; Park, O.K. Pathogen-Induced Autophagy Regulates Monolignol Transport and Lignin Formation in Plant Immunity. Autophagy 2023, 19, 597–615. [Google Scholar] [CrossRef] [PubMed]
- van Doorn, W.G.; Beers, E.P.; Dangl, J.L.; Franklin-Tong, V.E.; Gallois, P.; Hara-Nishimura, I.; Jones, A.M.; Kawai-Yamada, M.; Lam, E.; Mundy, J.; et al. Morphological Classification of Plant Cell Deaths. Cell Death Differ. 2011, 18, 1241–1246. [Google Scholar] [CrossRef]
- Wu, L.; Chen, H.; Curtis, C.; Fu, Z.Q. Go in for the Kill: How Plants Deploy Effector-Triggered Immunity to Combat Pathogens. Virulence 2014, 5, 710–721. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Model | Max-Likelihood-Value | AIC Value | Model | Max-Likelihood-Value | AIC Value |
|---|---|---|---|---|---|
| 1MG-AD * | −798.0823 | 1608.165 | MX1-AD-ADI | −975.5992 | 1967.198 |
| 1MG-A | −948.3228 | 1906.646 | MX1-AD-AD | −1011.491 | 2036.982 |
| 1MG-EAD | −942.8903 | 1895.781 | MX1-A-AD | −1035.123 | 2082.247 |
| 1MG-NCD | −944.8117 | 1899.623 | MX1-EAD-AD | −1035.123 | 2082.247 |
| 2MG-ADI | −974.6907 | 1971.381 | MX1-NCD-AD | −1011.491 | 2034.983 |
| 2MG-AD | −1016.11 | 2046.22 | MX2-ADI-ADI | −973.0078 | 1970.015 |
| 2MG-A | −1142.383 | 2294.765 | MX2-ADI-AD | −974.408 | 1966.816 |
| 2MG-EA | −909.9506 | 1827.901 | MX2-AD-AD | −1011.491 | 2032.982 |
| 2MG-CD | −1003.716 | 2017.432 | MX2-A-AD | −1035.123 | 2076.247 |
| 2MG-EAD | −938.4292 | 1884.859 | MX2-EA-AD | −1035.123 | 2074.247 |
| PG-ADI | −998.8515 | 2009.703 | MX2-CD-AD | −1011.491 | 2028.983 |
| PG-AD | −1091.609 | 2193.218 | MX2-EAD-AD | −1035.123 | 2074.247 |
| Model | Number | Model | Number | Model | Number |
|---|---|---|---|---|---|
| 1MG-AD | 1 * | 2MG-CD | 15 | MX1-NCD-AD | 17 |
| 1MG-A | 4 | 2MG-EAD | 2 | MX2-ADI-ADI | 17 |
| 1MG-EAD | 1 * | PG-ADI | 7 | MX2-ADI-AD | 8 |
| 1MG-NCD | 1 * | PG-AD | 6 | MX2-AD-AD | 17 |
| 2MG-ADI | 7 | MX1-AD-ADI | 9 | MX2-A-AD | 17 |
| 2MG-AD | 16 | MX1-AD-AD | 7 | MX2-EA-AD | 17 |
| 2MG-A | 5 | MX1-A-AD | 17 | MX2-CD-AD | 17 |
| 2MG-EA | 2 | MX1-EAD-AD | 17 | MX2-EAD-AD | 17 |
| Parameters | Estimate |
|---|---|
| m (population mean) | 14.1344 |
| d (additive effect of major gene) | −14.1344 |
| h (dominant effect of major gene) | −5.199 |
| PR (potential ratio) | 0.36 |
| hmg2 (%) (major gene heritability) | 93.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, S.; Kong, C.; Ma, H.; Ji, J.; Wang, Y.; Zhang, Y.; Zhuang, M.; Yang, L.; Fang, Z.; Taranov, V.; et al. Genetic and Cellular Basis of Resistance to Black Rot Caused by Xanthomonas campestris pv. campestris in Brassica rapa. Horticulturae 2025, 11, 626. https://doi.org/10.3390/horticulturae11060626
Deng S, Kong C, Ma H, Ji J, Wang Y, Zhang Y, Zhuang M, Yang L, Fang Z, Taranov V, et al. Genetic and Cellular Basis of Resistance to Black Rot Caused by Xanthomonas campestris pv. campestris in Brassica rapa. Horticulturae. 2025; 11(6):626. https://doi.org/10.3390/horticulturae11060626
Chicago/Turabian StyleDeng, Siping, Congcong Kong, Hongxue Ma, Jialei Ji, Yong Wang, Yangyong Zhang, Mu Zhuang, Limei Yang, Zhiyuan Fang, Vasiliy Taranov, and et al. 2025. "Genetic and Cellular Basis of Resistance to Black Rot Caused by Xanthomonas campestris pv. campestris in Brassica rapa" Horticulturae 11, no. 6: 626. https://doi.org/10.3390/horticulturae11060626
APA StyleDeng, S., Kong, C., Ma, H., Ji, J., Wang, Y., Zhang, Y., Zhuang, M., Yang, L., Fang, Z., Taranov, V., Artemyeva, A. M., & Lv, H. (2025). Genetic and Cellular Basis of Resistance to Black Rot Caused by Xanthomonas campestris pv. campestris in Brassica rapa. Horticulturae, 11(6), 626. https://doi.org/10.3390/horticulturae11060626