

Cereal Grain Arabinoxylans: Processing Effects and Structural Changes during Food and Beverage Fermentations †


Abstract
:1. Introduction
2. Native Arabinoxylan Structure in Cereal Grains
2.1. Degree of Arabinoxylan Backbone Substitution Varies between Grain Tissues and Species
2.2. Cereal Grain Arabinoxylans Are Feruloylated
2.3. Arabinoxylan Degree of Polymerization Varies between Grain Species
3. Arabinoxylans in Beer and Distilled Spirits Production: Processing Effects and Structural Changes
3.1. Malting
3.2. Mashing
3.3. Fermentation
3.4. Filtration
3.5. Viscosity
3.6. Arabinoxylans in Commercial Beer
3.7. Ferulic Acid in Brewing
4. Arabinoxylans in Bread: Processing Effects and Structural Changes
4.1. Interactions with Water
4.1.1. Water-Holding Capacity
4.1.2. Solubility Behavior of Arabinoxylans: Dynamic Shifts during Bread-Making
4.2. Molecular Interactions between Arabinoxylans and Gluten
4.3. Interactions with Starch
5. Future Research Directions
- More structural characterization work on AX structures following food processing is warranted. The bulk of AX structural characterization has targeted AX structures in raw cereal grains and ingredients, leaving many aspects of AX structures in prepared foods unexplored.
- Studies exploring human health effects of AXs should implement AX structures from as-consumed foods instead of from raw cereal grains or extracted AXs. As discussed in this review, AX structures are changed during fermented food processing. Explorations of the human health effects of AX consumption need to use AX structures that are representative of those in prepared foods, not those in raw grains.
- Studies attempting to clarify the effects of native AX structures on fermented food production should be wary of extrapolations from model systems using extracted AXs. The use of alkaline conditions to extract WU-AXs is the most pertinent example. Alkaline conditions will cleave ester-linked ferulates and release AX-AX crosslinks. This dramatically reduces the AX molecular weight and changes the solubility characteristics of the extracted AXs relative to their native states.
- Feruloylation is a consistent component of cereal grain AXs, so the ramifications of feruloylated side-chain branching patterns on AX-linked processing and human health effects should continue to be explored.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Araf | arabinofuranosyl |
| avDP | average degree of polymerization |
| avDS | average degree of substitution |
| AX | arabinoxylan |
| A/X | arabinose/xylose ratio |
| DFA | dehydrodiferulates |
| FA | ferulic acid |
| GMP | glutenin macropolymer |
| WE-AX | water-extractable arabinoxylan |
| WU-AX | water-unextractable arabinoxylan |
| Xylp | xylopyranosyl |
References
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef]
- Kang, X.; Kirui, A.; Dickwella Widanage, M.C.; Mentink-Vigier, F.; Cosgrove, D.J.; Wang, T. Lignin-polysaccharide interactions in plant secondary cell walls revealed by solid-state NMR. Nat. Commun. 2019, 10, 347. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, R.D.; Rancour, D.M.; Marita, J.M. Grass cell walls: A story of cross-linking. Front. Plant Sci. 2017, 7, 2056. [Google Scholar] [CrossRef] [PubMed]
- Terrett, O.M.; Dupree, P. Covalent interactions between lignin and hemicelluloses in plant secondary cell walls. Curr. Opin. Biotechnol. 2019, 56, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Buanafina, M.M.d.O.; Morris, P. The impact of cell wall feruloylation on plant growth, responses to environmental stress, plant pathogens and cell wall degradability. Agronomy 2022, 12, 1847. [Google Scholar] [CrossRef]
- Rancour, D.; Marita, J.; Hatfield, R. Cell wall composition throughout development for the model grass Brachypodium distachyon. Front. Plant Sci. 2012, 3, 266. [Google Scholar] [CrossRef]
- Appeldoorn, M.M.; de Waard, P.; Kabel, M.A.; Gruppen, H.; Schols, H.A. Enzyme resistant feruloylated xylooligomer analogues from thermochemically treated corn fiber contain large side chains, ethyl glycosides and novel sites of acetylation. Carbohydr. Res. 2013, 381, 33–42. [Google Scholar] [CrossRef]
- Appeldoorn, M.M.; Kabel, M.A.; Van Eylen, D.; Gruppen, H.; Schols, H.A. Characterization of oligomeric xylan structures from corn fiber resistant to pretreatment and simultaneous saccharification and fermentation. J. Agric. Food Chem. 2010, 58, 11294–11301. [Google Scholar] [CrossRef]
- Verbruggen, M.A.; Beldman, G.; Voragen, A.G.J. The selective extraction of glucuronoarabinoxylans from sorghum endosperm cell walls using barium and potassium hydroxide solutions. J. Cereal Sci. 1995, 21, 271–282. [Google Scholar] [CrossRef]
- Huisman, M.M.H.; Schols, H.A.; Voragen, A.G.J. Glucuronoarabinoxylans from maize kernel cell walls are more complex than those from sorghum kernel cell walls. Carbohydr. Polym. 2000, 43, 269–279. [Google Scholar] [CrossRef]
- Kabel, M.A.; Carvalheiro, F.; Garrote, G.; Avgerinos, E.; Koukios, E.; Parajó, J.C.; Gírio, F.M.; Schols, H.A.; Voragen, A.G.J. Hydrothermally treated xylan rich by-products yield different classes of xylo-oligosaccharides. Carbohydr. Polym. 2002, 50, 47–56. [Google Scholar] [CrossRef]
- Vitaglione, P.; Napolitano, A.; Fogliano, V. Cereal dietary fibre: A natural functional ingredient to deliver phenolic compounds into the gut. Trends Food Sci. Technol. 2008, 19, 451–463. [Google Scholar] [CrossRef]
- Bunzel, M. Monomere und Dimere PhenolcarbonsäUren als Strukturbildende Elemente in LöSlichen und UnlöSlichen Getreideballaststoffen. Ph.D. Thesis, Staats-Und Universitätsbibliothek Hamburg Carl von Ossietzky, Hamburg, Germany, 2001. [Google Scholar]
- Ralph, J.; Quideau, S.; Grabber, J.H.; Hatfield, R.D. Identification and synthesis of new ferulic acid dehydrodimers present in grass cell walls. J. Chem. Soc. Perkin Trans. 1 1994, 3485–3498. [Google Scholar] [CrossRef]
- Saulnier, L.; Crépeau, M.J.; Lahaye, M.; Thibault, J.F.; Garcia-Conesa, M.T.; Kroon, P.A.; Williamson, G. Isolation and structural determination of two 5,5’-diferuloyl oligosaccharides indicate that maize heteroxylans are covalently cross-linked by oxidatively coupled ferulates. Carbohydr. Res. 1999, 320, 82–92. [Google Scholar] [CrossRef]
- Allerdings, E.; Ralph, J.; Schatz, P.F.; Gniechwitz, D.; Steinhart, H.; Bunzel, M. Isolation and structural identification of diarabinosyl 8-O-4-dehydrodiferulate from maize bran insoluble fibre. Phytochemistry 2005, 66, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Bunzel, M.; Allerdings, E.; Ralph, J.; Steinhart, H. Cross-linking of arabinoxylans via 8-8-coupled diferulates as demonstrated by isolation and identification of diarabinosyl 8-8(cyclic)-dehydrodiferulate from maize bran. J. Cereal Sci. 2008, 47, 29–40. [Google Scholar] [CrossRef]
- Ralph, J.; Grabber, J.H.; Hatfield, R.D. Lignin-ferulate cross-links in grasses: Active incorporation of ferulate polysaccharide esters into ryegrass lignins. Carbohydr. Res. 1995, 275, 167–178. [Google Scholar] [CrossRef]
- Bunzel, M.; Ralph, J.; Marita, J.M.; Hatfield, R.D.; Steinhart, H. Diferulates as structural components in soluble and insoluble cereal dietary fibre. J. Sci. Food Agric. 2001, 81, 653–660. [Google Scholar] [CrossRef]
- Schendel, R.R.; Meyer, M.R.; Bunzel, M. Quantitative profiling of feruloylated arabinoxylan side-chains from graminaceous cell walls. Front. Plant Sci. 2016, 6, 1249. [Google Scholar] [CrossRef]
- Kabel, M.A.; van den Borne, H.; Vincken, J.P.; Voragen, A.G.J.; Schols, H.A. Structural differences of xylans affect their interaction with cellulose. Carbohydr. Polym. 2007, 69, 94–105. [Google Scholar] [CrossRef]
- Köhnke, T.; Pujolras, C.; Roubroeks, J.P.; Gatenholm, P. The effect of barley husk arabinoxylan adsorption on the properties of cellulose fibres. Cellulose 2008, 15, 537–546. [Google Scholar] [CrossRef]
- Selig, M.J.; Thygesen, L.G.; Felby, C.; Master, E.R. Debranching of soluble wheat arabinoxylan dramatically enhances recalcitrant binding to cellulose. Biotechnol. Lett. 2015, 37, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Pollet, A.; Delcour, J.A.; Courtin, C.M. Structural determinants of the substrate specificities of xylanases from different glycoside hydrolase families. Crit. Rev. Biotechnol. 2010, 30, 176–191. [Google Scholar] [CrossRef] [PubMed]
- Brillouet, J.M.; Joseleau, J.P. Investigation of the structure of a heteroxylan from the outer pericarp (beeswing bran) of wheat kernel. Carbohydr. Res. 1987, 159, 109–126. [Google Scholar] [CrossRef]
- Dervilly-Pinel, G.; Rimsten, L.; Saulnier, L.; Andersson, R.; Åman, P. Water-extractable arabinoxylan from pearled flours of wheat, barley, rye and triticale. Evidence for the presence of ferulic acid dimers and their involvement in gel formation. J. Cereal Sci. 2001, 34, 207–214. [Google Scholar] [CrossRef]
- Gruppen, H.; Kormelink, F.J.M.; Voragen, A.G.J. Water-unextractable cell wall material from wheat flour. 3. A structural model for arabinoxylans. J. Cereal Sci. 1993, 18, 111–128. [Google Scholar] [CrossRef]
- Shewry, P.R.; Piironen, V.; Lampi, A.M.; Edelmann, M.; Kariluoto, S.; Nurmi, T.; Fernandez-Orozco, R.; Andersson, A.A.M.; Åman, P.; Fraś, A.; et al. Effects of genotype and environment on the content and composition of phytochemicals and dietary fiber components in rye in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2010, 58, 9372–9383. [Google Scholar] [CrossRef]
- Shewry, P.R.; Hawkesford, M.J.; Piironen, V.; Lampi, A.M.; Gebruers, K.; Boros, D.; Andersson, A.A.M.; Åman, P.; Rakszegi, M.; Bedo, Z.; et al. Natural variation in grain composition of wheat and related cereals. J. Agric. Food Chem. 2013, 61, 8295–8303. [Google Scholar] [CrossRef]
- Gebruers, K.; Dornez, E.; Boros, D.; Fraś, A.; Dynkowska, W.; Bedő, Z.; Rakszegi, M.; Delcour, J.A.; Courtin, C.M. Variation in the content of dietary fiber and components thereof in wheats in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2008, 56, 9740–9749. [Google Scholar] [CrossRef]
- Vinkx, C.; Reynaert, H.; Grobet, P.; Delcour, J. Physicochemical and functional properties of rye nonstarch polysaccharides. V. Variability in the structure of water-soluble arabinoxylans. Cereal Chem. 1993, 70, 311. [Google Scholar]
- Andersson, A.A.M.; Lampi, A.M.; Nyström, L.; Piironen, V.; Li, L.; Ward, J.L.; Gebruers, K.; Courtin, C.M.; Delcour, J.A.; Boros, D.; et al. Phytochemical and dietary fiber components in barley varieties in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2008, 56, 9767–9776. [Google Scholar] [CrossRef] [PubMed]
- Collins, H.M.; Burton, R.A.; Topping, D.L.; Liao, M.; Bacic, A.; Fincher, G.B. Variability in fine structures of noncellulosic cell wall polysaccharides from cereal grains: Potential importance in human health and nutrition. Cereal Chem. 2010, 87, 272–282. [Google Scholar] [CrossRef]
- Xu, H.; Reuhs, B.L.; Cantu-Jungles, T.M.; Tuncil, Y.E.; Kaur, A.; Terekhov, A.; Martens, E.C.; Hamaker, B.R. Corn arabinoxylan has a repeating structure of subunits of high branch complexity with slow gut microbiota fermentation. Carbohydr. Polym. 2022, 289, 119435. [Google Scholar] [CrossRef]
- Bunzel, M. Chemistry and occurrence of hydroxycinnamate oligomers. Phytochem. Rev. 2010, 9, 47–64. [Google Scholar] [CrossRef]
- Jilek, M.L.; Bunzel, M. Dehydrotriferulic and dehydrodiferulic acid profiles of cereal and pseudocereal flours. Cereal Chem. 2013, 90, 507–514. [Google Scholar] [CrossRef]
- Waterstraat, M.; Bunzel, M. A stable isotope dilution approach to analyze ferulic acid oligomers in plant cell walls using liquid chromatography-tandem mass spectrometry. Anal. Bioanal. Chem. 2019, 411, 5047–5062. [Google Scholar] [CrossRef]
- Grabber, J.; Hatfield, R.; Ralph, J. Diferulate cross-links impede the enzymatic degradation of non-lignified maize walls. J. Sci. Food Agric. 1998, 77, 193–200. [Google Scholar] [CrossRef]
- Gruppen, H.; Hamer, R.; Voragen, A. Water-unextractable cell wall material from wheat flour. 1. Extraction of polymers with alkali. J. Cereal Sci. 1992, 16, 41–51. [Google Scholar] [CrossRef]
- Courtin, C.M.; Roelants, A.; Delcour, J.A. Fractionation–reconstitution experiments provide insight into the role of endoxylanases in bread-making. J. Agric. Food Chem. 1999, 47, 1870–1877. [Google Scholar] [CrossRef]
- Buksa, K.; Nowotna, A.; Ziobro, R.; Praznik, W. Molecular properties of arabinoxylan fractions isolated from rye grain of different quality. J. Cereal Sci. 2014, 60, 368–373. [Google Scholar] [CrossRef]
- Izydorczyk, M.; Biliaderis, C. Structural heterogeneity of wheat endosperm arabinoxylans. Cereal Chem. 1993, 70, 641–646. [Google Scholar]
- Dervilly, G.; Saulnier, L.; Roger, P.; Thibault, J.F. Isolation of homogeneous fractions from wheat water-soluble arabinoxylans. Influence of the structure on their macromolecular characteristics. J. Agric. Food Chem. 2000, 48, 270–278. [Google Scholar] [CrossRef]
- Dervilly-Pinel, G.; Thibault, J.F.; Saulnier, L. Experimental evidence for a semi-flexible conformation for arabinoxylans. Carbohydr. Res. 2001, 330, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Maes, C.; Delcour, J.A. Structural characterisation of water-extractable and water-unextractable arabinoxylans in wheat bran. J. Cereal Sci. 2002, 35, 315–326. [Google Scholar] [CrossRef]
- Dervilly, G.; Leclercq, C.; Zimmermann, D.; Roue, C.; Thibault, J.F.; Saulnier, L. Isolation and characterization of high molar mass water-soluble arabinoxylans from barley and barley malt. Carbohydr. Polym. 2002, 47, 143–149. [Google Scholar] [CrossRef]
- Verwimp, T.; Van Craeyveld, V.; Courtin, C.M.; Delcour, J.A. Variability in the structure of rye flour alkali-extractable arabinoxylans. J. Agric. Food Chem. 2007, 55, 1985–1992. [Google Scholar] [CrossRef]
- Andersson, R.; Fransson, G.; Tietjen, M.; Åman, P. Content and molecular-weight distribution of dietary fiber components in whole-grain rye flour and bread. J. Agric. Food Chem. 2009, 57, 2004–2008. [Google Scholar] [CrossRef]
- Chanliaud, E.; Saulnier, L.; Thibault, J.F. Alkaline extraction and characterisation of heteroxylans from maize bran. J. Cereal Sci. 1995, 21, 195–203. [Google Scholar] [CrossRef]
- Mendis, M.; Simsek, S. Production of structurally diverse wheat arabinoxylan hydrolyzates using combinations of xylanase and arabinofuranosidase. Carbohydr. Polym. 2015, 132, 452–459. [Google Scholar] [CrossRef]
- Langenaeken, N.A.; De Schepper, C.F.; De Schutter, D.P.; Courtin, C.M. Carbohydrate content and structure during malting and brewing: A mass balance study. J. Inst. Brew. 2020, 126, 253–262. [Google Scholar] [CrossRef]
- Li, Y.; Lu, J.; Gu, G.; Shi, Z.; Mao, Z. Studies on water-extractable arabinoxylans during malting and brewing. Food Chem. 2005, 93, 33–38. [Google Scholar] [CrossRef]
- Sungurtas, J.; Swanston, J.; Davies, H.; McDougall, G. Xylan-degrading enzymes and arabinoxylan solubilisation in barley cultivars of differing malting quality. J. Cereal Sci. 2004, 39, 273–281. [Google Scholar] [CrossRef]
- Betts, N.S.; Wilkinson, L.G.; Khor, S.F.; Shirley, N.J.; Lok, F.; Skadhauge, B.; Burton, R.A.; Fincher, G.B.; Collins, H.M. Morphology, carbohydrate distribution, gene expression, and enzymatic activities related to cell wall hydrolysis in four barley varieties during simulated malting. Front. Plant Sci. 2017, 8, 1872. [Google Scholar] [CrossRef]
- Krahl, M.; Müller, S.; Zarnkow, M.; Back, W.; Becker, T. Arabinoxylan and fructan in the malting and brewing process. Qual. Assur. Saf. Crop. Foods 2009, 1, 246–255. [Google Scholar] [CrossRef]
- Guo, M.; Du, J.; Zhang, K.; Jin, Y. Content and molecular weight of water-extractable arabinoxylans in wheat malt and wheat malt-based wort with different Kolbach indices. J. Sci. Food Agric. 2014, 94, 2794–2800. [Google Scholar] [CrossRef] [PubMed]
- Debyser, W.; Derdelinckx, G.; Delcour, J.A. Arabinoxylan and arabinoxylan hydrolyzing activities in barley malts and worts derived from them. J. Cereal Sci. 1997, 26, 67–74. [Google Scholar] [CrossRef]
- Li, Y.; Lu, J.; Gu, G. Control of arabinoxylan solubilization and hydrolysis in mashing. Food Chem. 2005, 90, 101–108. [Google Scholar] [CrossRef]
- Peng, Z.; Jin, Y.; Du, J. Enzymatic properties of endo-1,4-β-xylanase from wheat malt. Protein Pept. Lett. 2019, 26, 332–338. [Google Scholar] [CrossRef]
- Viëtor, R.J.; Voragen, A.G.J.; Angelino, S.A.G.F. Composition of non-starch polysaccharides in wort and spent grain from brewing trials with malt from a good malting quality barley and a feed barley. J. Inst. Brew. 1993, 99, 243–248. [Google Scholar] [CrossRef]
- Michiels, P.; Delputte, N.; Debyser, W.; Langenaeken, N.A.; Courtin, C.M. The occurrence and structural heterogeneity of arabinoxylan in commercial pilsner beers and their non-alcoholic counterparts. Carbohydr. Polym. 2023, 306, 120597. [Google Scholar] [CrossRef]
- Shang, Y.L.; Li, X.M.; Cai, G.L.; Lu, J. The role of ferulic acid and arabinoxylan in inducing premature yeast flocculation. J. Inst. Brew. 2015, 121, 49–54. [Google Scholar] [CrossRef]
- Xie, Y.; Cai, G.; Xu, M.; Han, B.; Li, C.; Lu, J. The effect of barley infected with xylanase-producing filamentous fungi on premature yeast flocculation. J. Inst. Brew. 2022, 128, 162–170. [Google Scholar] [CrossRef]
- Li, J.; Du, J.; Wu, X.; Zhang, Z.; Zhang, K. Changes in crude arabinoxylan during cloudy wheat beer brewing on a production scale. J. Inst. Brew. 2017, 123, 192–198. [Google Scholar] [CrossRef]
- Martinez Amezaga, N.M.J.; Lataza Rovaletti, M.M.; Benitez, E.I. Particle size distribution of polysaccharides in beer before the filtration process. Int. J. Food Res. 2017, 5, 13–19. [Google Scholar]
- Jin, Y.L.; Speers, R.A.; Paulson, A.T.; Stewart, R.J. Effect of β-glucans and process conditions on the membrane filtration performance of beer. J. Am. Soc. Brew. Chem. 2004, 62, 117–124. [Google Scholar] [CrossRef]
- Jin, Y.L.; Speers, A.; Paulson, A.T.; Stewart, R.J. Effects of β-glucans and environmental factors on the viscosities of wort and beer. J. Inst. Brew. 2004, 110, 104–116. [Google Scholar] [CrossRef]
- Sadosky, P.; Schwarz, P.B.; Horsley, R.D. Effect of arabinoxylans, β-glucans, and dextrins on the viscosity and membrane filterability of a beer model solution. J. Am. Soc. Brew. Chem. 2002, 60, 153–162. [Google Scholar] [CrossRef]
- Egi, A.; Speers, R.; Paulson, A. The physical behavior of arabinoxylans in model brewing solutions. Tech. Q.-Master Brew. Assoc. Am. 2004, 41, 268–276. [Google Scholar]
- Lu, J.; Li, Y.; Gu, G.; Mao, Z. Effects of molecular weight and concentration of arabinoxylans on the membrane plugging. J. Agric. Food Chem. 2005, 53, 4996–5002. [Google Scholar] [CrossRef]
- Gastl, M.; Kupetz, M.; Becker, T. Determination of cytolytic malt modification – Part II: Impact on wort separation. J. Am. Soc. Brew. Chem. 2021, 79, 66–74. [Google Scholar] [CrossRef]
- Shrestha, U.R.; Smith, S.; Pingali, S.V.; Yang, H.; Zahran, M.; Breunig, L.; Wilson, L.A.; Kowali, M.; Kubicki, J.D.; Cosgrove, D.J.; et al. Arabinose substitution effect on xylan rigidity and self-aggregation. Cellulose 2019, 26, 2267–2278. [Google Scholar] [CrossRef]
- Lu, J.; Li, Y. Effects of arabinoxylan solubilization on wort viscosity and filtration when mashing with grist containing wheat and wheat malt. Food Chem. 2006, 98, 164–170. [Google Scholar] [CrossRef]
- Marconi, O.; Alfeo, V.; Tomasi, I.; Maranghi, S.; De Francesco, G.; Sileoni, V.; Perretti, G. Effects of malting process on molecular weight distribution and content of total and water-extractable arabinoxylan in barley. J. Cereal Sci. 2022, 107. [Google Scholar] [CrossRef]
- Mangan, D.; Cornaggia, C.; Liadova, A.; Draga, A.; Ivory, R.; Evans, D.E.; McCleary, B.V. Development of an automatable method for the measurement of endo-1,4-β-xylanase activity in barley malt and initial investigation into the relationship between endo-1,4-β-xylanase activity and wort viscosity. J. Cereal Sci. 2018, 84, 90–94. [Google Scholar] [CrossRef]
- Schwarz, P.B.; Han, J.Y. Arabinoxylan content of commercial beers. J. Am. Soc. Brew. Chem. 1995, 53, 157–159. [Google Scholar] [CrossRef]
- Courtin, C.M.; Broekaert, W.F.; Swennen, K.; Aerts, G.; Van Craeyveld, V.; Delcour, J.A. Occurrence of arabinoxylo-oligosaccharides and arabinogalactan peptides in beer. J. Am. Soc. Brew. Chem. 2009, 67, 112–117. [Google Scholar] [CrossRef]
- Evans, D.; Sheehan, M.; Stewart, D. The impact of malt derived proteins on beer foam quality. Part II: The influence of malt foam-positive proteins and non-starch polysaccharides on beer foam quality. J. Inst. Brew. 1999, 105, 171–178. [Google Scholar] [CrossRef]
- Li, J.; Du, J. Molecular characterization of arabinoxylan from wheat beer, beer foam and defoamed beer. Molecules 2019, 24, 1230. [Google Scholar] [CrossRef]
- Sohrabvandi, S.; Mousavi, S.M.; Razavi, S.H.; Mortazavian, A.M.; Rezaei, K. Alcohol-free beer: Methods of production, sensorial defects, and healthful effects. Food Rev. Int. 2010, 26, 335–352. [Google Scholar] [CrossRef]
- Langenaeken, N.A.; De Schutter, D.P.; Courtin, C.M. Arabinoxylan from non-malted cereals can act as mouthfeel contributor in beer. Carbohydr. Polym. 2020, 239, 116257. [Google Scholar] [CrossRef]
- Sancho, A.I.; Faulds, C.B.; Bartolomé, B.; Williamson, G. Characterisation of feruloyl esterase activity in barley. J. Sci. Food Agric. 1999, 79, 447–449. [Google Scholar] [CrossRef]
- Vanbeneden, N.; Gils, F.; Delvaux, F.; Delvaux, F.R. Variability in the release of free and bound hydroxycinnamic acids from diverse malted barley (Hordeum vulgare L.) Cultiv. Wort Prod. J. Agric. Food Chem. 2007, 55, 11002–11010. [Google Scholar] [CrossRef]
- Coghe, S.; Benoot, K.; Delvaux, F.; Vanderhaegen, B.; Delvaux, F.R. Ferulic acid release and 4-vinylguaiacol formation during brewing and fermentation: Indications for feruloyl esterase activity in Saccharomyces cerevisiae. J. Agric. Food Chem. 2004, 52, 602–608. [Google Scholar] [CrossRef]
- McLauchlan, W.R.; Garcia-Conesa, M.T.; Williamson, G.; Roza, M.; Ravestein, P.; Maat, J. A novel class of protein from wheat which inhibits xylanases. Biochem. J. 1999, 338, 441–446. [Google Scholar] [CrossRef]
- Fierens, E.; Rombouts, S.; Gebruers, K.; Goesaert, H.; Brijs, K.; Beaugrand, J.; Volckaert, G.; Van Campenhout, S.; Proost, P.; Courtin, C.; et al. TLXI, a novel type of xylanase inhibitor from wheat (Triticum aestivum) Belong. Thaum. Fam. Biochem. J. 2007, 403, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Kalb, V.; Seewald, T.; Hofmann, T.; Granvogl, M. Studies on the impact of malting and mashing on the free, soluble ester-bound, and insoluble ester-bound forms of desired and undesired phenolic acids aiming at styrene mitigation during wheat beer brewing. J. Agric. Food Chem. 2020, 68, 12421–12432. [Google Scholar] [CrossRef]
- Vanbeneden, N.; Van Roey, T.; Willems, F.; Delvaux, F.; Delvaux, F.R. Release of phenolic flavour precursors during wort production: Influence of process parameters and grist composition on ferulic acid release during brewing. Food Chem. 2008, 111, 83–91. [Google Scholar] [CrossRef]
- Poisson, L.; Schieberle, P. Characterization of the most odor-active compounds in an American bourbon whisky by application of the aroma extract dilution analysis. J. Agric. Food Chem. 2008, 56, 5813–5819. [Google Scholar] [CrossRef]
- Leys, S.; De Bondt, Y.; Bosmans, G.; Courtin, C.M. Assessing the impact of xylanase activity on the water distribution in wheat dough: A 1H NMR study. Food Chem. 2020, 325, 126828. [Google Scholar] [CrossRef]
- Wang, M.; Hamer, R.J.; van Vliet, T.; Gruppen, H.; Marseille, H.; Weegels, P.L. Effect of water-unextractable solids on gluten formation and properties: Mechanistic considerations. J. Cereal Sci. 2003, 37, 55–64. [Google Scholar] [CrossRef]
- Courtin, C.M.; Gelders, G.G.; Delcour, J.A. Use of two endoxylanases with different substrate selectivity for understanding arabinoxylan functionality in wheat flour breadmaking. Cereal Chem. 2001, 78, 564–571. [Google Scholar] [CrossRef]
- Courtin, C.M.; Delcour, J.A. Arabinoxylans and endoxylanases in wheat flour bread-making. J. Cereal Sci. 2002, 35, 225–243. [Google Scholar] [CrossRef]
- Nishitsuji, Y.; Whitney, K.; Nakamura, K.; Hayakawa, K.; Simsek, S. Changes in structure and solubility of wheat arabinoxylan during the breadmaking process. Food Hydrocoll. 2020, 109, 106129. [Google Scholar] [CrossRef]
- Biliaderis, C.G.; Izydorczyk, M.S.; Rattan, O. Effect of arabinoxylans on bread-making quality of wheat flours. Food Chem. 1995, 53, 165–171. [Google Scholar] [CrossRef]
- Pietiäinen, S.; Moldin, A.; Ström, A.; Malmberg, C.; Langton, M. Effect of physicochemical properties, pre-processing, and extraction on the functionality of wheat bran arabinoxylans in breadmaking—A review. Food Chem. 2022, 383, 132584. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, Z.; Niu, M.; Zhao, S.; Jia, C.; Wu, Y. Thermomechanical behaviors and protein polymerization in bread dough modified by bran components and transglutaminase. LWT 2020, 133, 109894. [Google Scholar] [CrossRef]
- Gudmundsson, M.; Eliasson, A.C.; Bengtsson, S.; Aman, P. The effects of water-soluble arabinoxylan on gelatinization and retrogradation of starch. Starch—Stärke 1991, 43, 5–10. [Google Scholar] [CrossRef]
- Zhang, L.; van Boven, A.; Mulder, J.; Grandia, J.; Chen, X.D.; Boom, R.M.; Schutyser, M.A.I. Arabinoxylans-enriched fractions: From dry fractionation of wheat bran to the investigation on bread baking performance. J. Cereal Sci. 2019, 87, 1–8. [Google Scholar] [CrossRef]
- Kaur, A.; Singh, B.; Yadav, M.P.; Bhinder, S.; Singh, N. Isolation of arabinoxylan and cellulose-rich arabinoxylan from wheat bran of different varieties and their functionalities. Food Hydrocoll. 2021, 112, 106287. [Google Scholar] [CrossRef]
- Li, J.; Kang, J.; Wang, L.; Li, Z.; Wang, R.; Chen, Z.X.; Hou, G.G. Effect of water migration between arabinoxylans and gluten on baking quality of whole wheat bread detected by magnetic resonance imaging (MRI). J. Agric. Food Chem. 2012, 60, 6507–6514. [Google Scholar] [CrossRef]
- Cleemput, G.; Booij, C.; Hessing, M.; Gruppen, H.; Delcour, J. Solubilisation and changes in molecular weight distribution of arabinoxylans and protein in wheat flours during bread-making, and the effects of endogenous arabinoxylan hydrolysing enzymes. J. Cereal Sci. 1997, 26, 55–66. [Google Scholar] [CrossRef]
- Zhang, D.; Rudjito, R.C.; Pietiäinen, S.; Chang, S.C.; Idström, A.; Evenäs, L.; Vilaplana, F.; Jiménez-Quero, A. Arabinoxylan supplemented bread: From extraction of fibers to effect of baking, digestion, and fermentation. Food Chem. 2023, 413, 135660. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liu, R.; Wu, T.; Zhang, M. Aggregation and rheological behavior of soluble dietary fibers from wheat bran. Food Res. Int. 2017, 102, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Nishitsuji, Y.; Whitney, K.; Nakamura, K.; Hayakawa, K.; Simsek, S. Analysis of molecular weight and structural changes in water-extractable arabinoxylans during the breadmaking process. Food Chem. 2022, 386, 132772. [Google Scholar] [CrossRef] [PubMed]
- Kiszonas, A.M.; Fuerst, E.P.; Luthria, D.; Morris, C.F. Arabinoxylan content and characterisation throughout the bread-baking process. Int. J. Food Sci. Technol. 2015, 50, 1911–1921. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, C.; Yang, M.; Ou, Z.; Lin, Y.; Zhao, F.; Han, S. Characterization and application of a novel xylanase from Halolactibacillus miurensis Wholewheat Bread Making. Front. Bioeng. Biotechnol. 2022, 10, 1018476. [Google Scholar] [CrossRef]
- Wang, P.; Hou, C.; Zhao, X.; Tian, M.; Gu, Z.; Yang, R. Molecular characterization of water-extractable arabinoxylan from wheat bran and its effect on the heat-induced polymerization of gluten and steamed bread quality. Food Hydrocoll. 2019, 87, 570–581. [Google Scholar] [CrossRef]
- Zhu, X.F.; Tao, H.; Wang, H.L.; Xu, X.M. Impact of water soluble arabinoxylan on starch-gluten interactions in dough. LWT 2023, 173, 114289. [Google Scholar] [CrossRef]
- Buksa, K.; Nowotna, A.; Ziobro, R. Application of cross-linked and hydrolyzed arabinoxylans in baking of model rye bread. Food Chem. 2016, 192, 991–996. [Google Scholar] [CrossRef]
- Figueroa-Espinoza, M.C.; Rouau, X. Oxidative cross-linking of pentosans by a fungal laccase and horseradish peroxidase: Mechanism of linkage between feruloylated arabinoxylans. Cereal Chem. 1998, 75, 259–265. [Google Scholar] [CrossRef]
- Goesaert, H.; Brijs, K.; Veraverbeke, W.S.; Courtin, C.M.; Gebruers, K.; Delcour, J.A. Wheat flour constituents: How they impact bread quality, and how to impact their functionality. Trends Food Sci. Technol. 2005, 16, 12–30. [Google Scholar] [CrossRef]
- Si, X.; Li, T.; Zhang, Y.; Zhang, W.; Qian, H.; Li, Y.; Zhang, H.; Qi, X.; Wang, L. Interactions between gluten and water-unextractable arabinoxylan during the thermal treatment. Food Chem. 2021, 345, 128785. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, Y.; Li, J.; Li, F.; Teng, C.; Li, X. Effects of water-extractable arabinoxylan on the physicochemical properties and structure of wheat gluten by thermal treatment. J. Agric. Food Chem. 2017, 65, 4728–4735. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, F.; Wang, Y.; Li, J.; Teng, C.; Wang, C.; Li, X. Effects of different molecular weight water-extractable arabinoxylans on the physicochemical properties and structure of wheat gluten. J. Food Sci. Technol. 2019, 56, 340–349. [Google Scholar] [CrossRef]
- Sun, J.; Si, X.; Li, T.; Zhao, J.; Qian, H.; Li, Y.; Zhang, H.; Qi, X.; Wang, L. The influence of water-unextractable arabinoxylan and its hydrolysates on the aggregation and structure of gluten proteins. Front. Nutr. 2022, 9, 877135. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Tao, H.; Jin, Z.; Xu, X. Impact of water extractable arabinoxylan from rye bran on the frozen steamed bread dough quality. Food Chem. 2016, 200, 117–124. [Google Scholar] [CrossRef]
- Zhao, X.; Hou, C.; Tian, M.; Zhou, Y.; Yang, R.; Wang, X.; Gu, Z.; Wang, P. Effect of water-extractable arabinoxylan with different molecular weight on the heat-induced aggregation behavior of gluten. Food Hydrocoll. 2020, 99, 105318. [Google Scholar] [CrossRef]
- Beck, M.; Jekle, M.; Selmair, P.L.; Koehler, P.; Becker, T. Rheological properties and baking performance of rye dough as affected by transglutaminase. J. Cereal Sci. 2011, 54, 29–36. [Google Scholar] [CrossRef]
- Döring, C.; Nuber, C.; Stukenborg, F.; Jekle, M.; Becker, T. Impact of arabinoxylan addition on protein microstructure formation in wheat and rye dough. J. Food Eng. 2015, 154, 10–16. [Google Scholar] [CrossRef]
- Döring, C.; Hussein, M.A.; Jekle, M.; Becker, T. On the assessments of arabinoxylan localization and enzymatic modifications for enhanced protein networking and its structural impact on rye dough and bread. Food Chem. 2017, 229, 178–187. [Google Scholar] [CrossRef]
- Piber, M.; Koehler, P. Identification of dehydro-ferulic acid-tyrosine in rye and wheat: Evidence for a covalent cross-link between arabinoxylans and proteins. J. Agric. Food Chem. 2005, 53, 5276–5284. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, S.; Kang, J.; Wang, N.; Xiao, M.; Li, Z.; Wang, C.; Guo, Q.; Hu, X. Arabinoxylan from wheat bran: Molecular degradation and functional investigation. Food Hydrocoll. 2020, 107, 105914. [Google Scholar] [CrossRef]
- Wang, P.; Li, D.; Hou, C.; Yang, T.; Yang, R.; Gu, Z.; Jiang, D. Tailormade wheat arabinoxylan reveals the role of substitution in regulating gelatinization and retrogradation behavior of wheat starch. J. Agric. Food Chem. 2022, 70, 1659–1669. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.; Zhao, X.; Tian, M.; Zhou, Y.; Yang, R.; Gu, Z.; Wang, P. Impact of water extractable arabinoxylan with different molecular weight on the gelatinization and retrogradation behavior of wheat starch. Food Chem. 2020, 318, 126477. [Google Scholar] [CrossRef]
- Rosicka-Kaczmarek, J.; Tkaczyk, M.; Makowski, B.; Komisarczyk, A.; Nebesny, E. The influence of non-starch polysaccharide on thermodynamic properties of starches from facultative wheat varieties. Eur. Food Res. Technol. 2017, 243, 2243–2253. [Google Scholar] [CrossRef]
- Ying, R.; Zhou, T.; Xie, H.; Huang, M. Synergistic effect of arabinoxylan and (1,3)(1,4)-β-glucan reduces the starch hydrolysis rate in wheat flour. Food Hydrocoll. 2023, 141, 108668. [Google Scholar] [CrossRef]
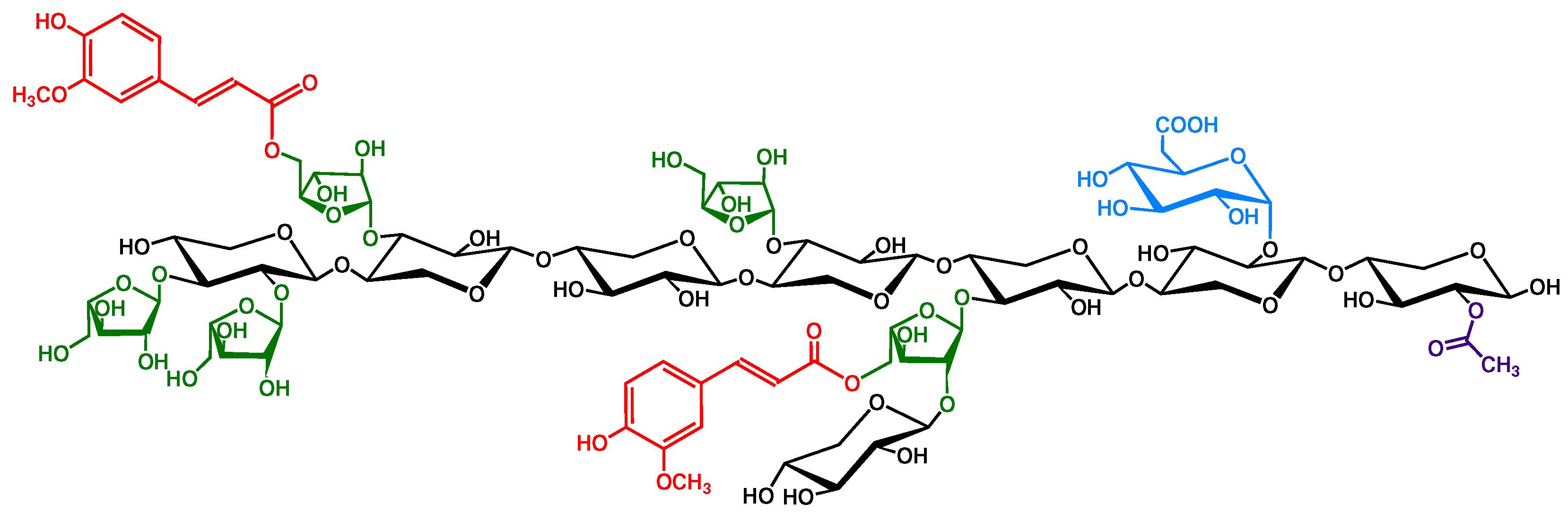

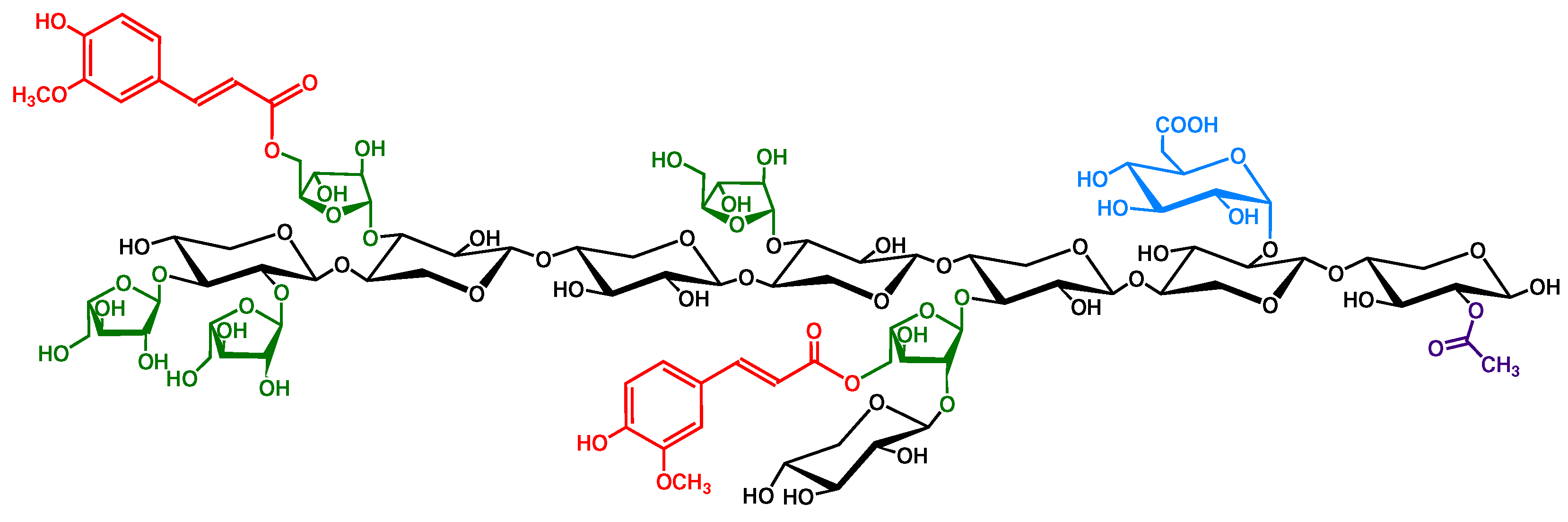{kind=link}
| Cereal Grain, Tissue (Reference) | AX Extraction Method | × 10 (g/mol) | Analytical Method |
|---|---|---|---|
| Wheat | |||
| Endosperm [44] | water-soluble (treated with 1M NaOH for 2 h after extraction to remove ester-linked phenolics) | 215–285 | HPSEC-MALLS |
| Industrial bran [45] | water-soluble | 5 and 50 | Gel filtration, RI detection, DC |
| Industrial bran [45] | alkaline (2% HO, pH 11.5) | 5–10 and 100–140 | Gel filtration, RI detection, DC |
| Barley | |||
| Endosperm [26] | water-soluble | 237–255 | HPSEC-MALLS |
| Whole grain [46] | water soluble | 340 | HPSEC-MALLS |
| Industrial bran [46] | water-soluble | 250 | HPSEC-MALLS |
| Rye | |||
| Endosperm [26] | water-soluble | 288 | HPSEC-MALLS |
| Endosperm [47] | alkaline (Ba(OH)) | 137–1306 | HPSEC-RI, DC |
| Whole grain flour [48] | water-soluble | 200 | HPSEC-MALLS |
| Whole grain flour [41] | water-soluble | 140–263 | HPSEC-RI, DC |
| Whole grain flour [41] | alkaline | 244–309 | HPSEC-RI, DC |
| Sourdough bread [48] | water-soluble | 70 | HPSEC-MALLS |
| Corn | |||
| Bran [49] | alkaline | 220–230 | HPSEC-RI, DC |
| Endosperm [10] | alkaline | 171–400 | HPSEC-RALLS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tse, T.; Schendel, R.R. Cereal Grain Arabinoxylans: Processing Effects and Structural Changes during Food and Beverage Fermentations. Fermentation 2023, 9, 914. https://doi.org/10.3390/fermentation9100914
Tse T, Schendel RR. Cereal Grain Arabinoxylans: Processing Effects and Structural Changes during Food and Beverage Fermentations. Fermentation. 2023; 9(10):914. https://doi.org/10.3390/fermentation9100914
Chicago/Turabian StyleTse, Tianbo, and Rachel R. Schendel. 2023. "Cereal Grain Arabinoxylans: Processing Effects and Structural Changes during Food and Beverage Fermentations" Fermentation 9, no. 10: 914. https://doi.org/10.3390/fermentation9100914
APA StyleTse, T., & Schendel, R. R. (2023). Cereal Grain Arabinoxylans: Processing Effects and Structural Changes during Food and Beverage Fermentations. Fermentation, 9(10), 914. https://doi.org/10.3390/fermentation9100914