
Evaluation of Nutritional Composition of Pure Filamentous Fungal Biomass as a Novel Ingredient for Fish Feed

 ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation Medium
2.2. Fungi Species
2.3. Cultivation in Shake-Flasks
2.4. Analytical Procedures
2.5. Statistical Analysis
3. Results and Discussions
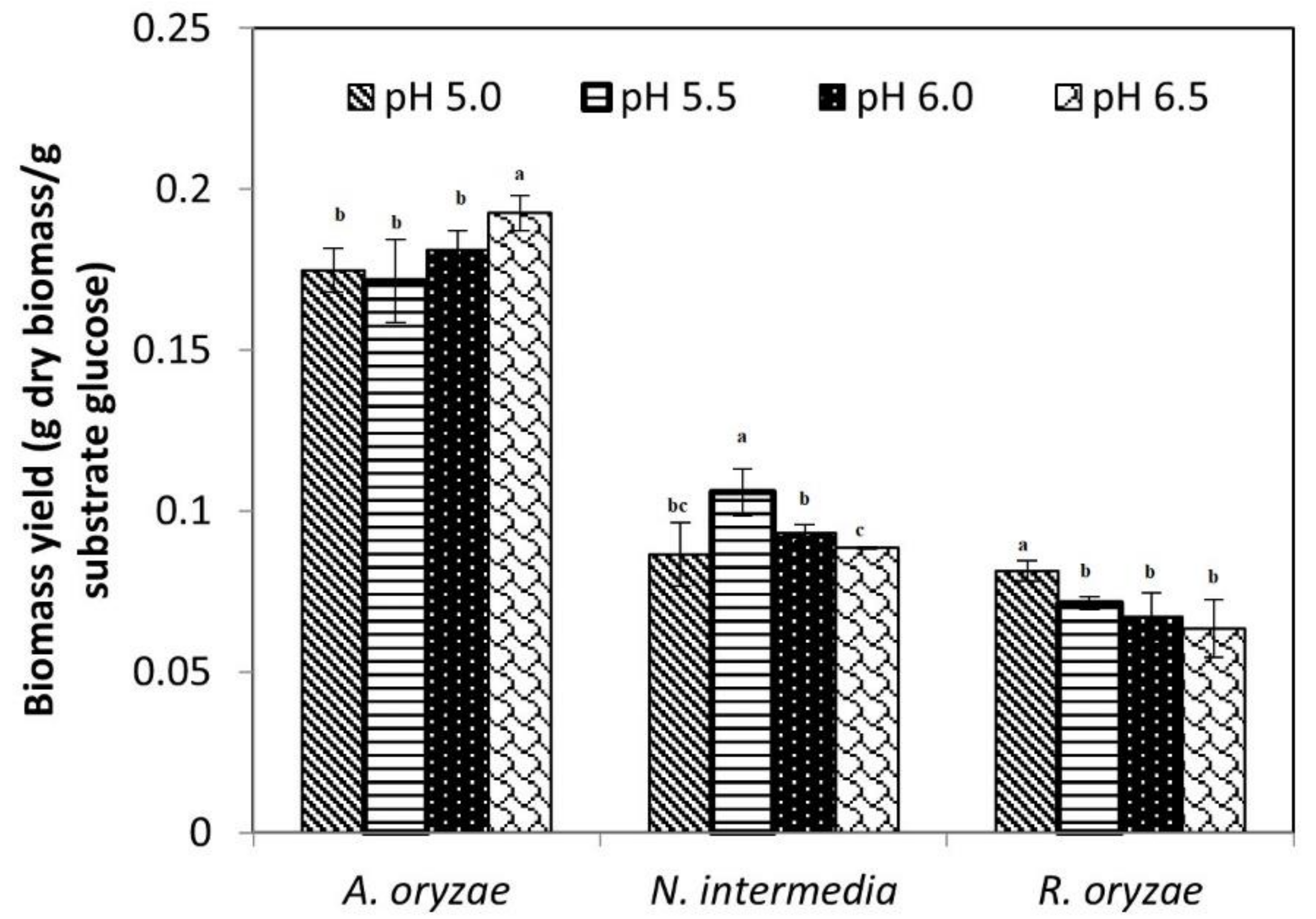
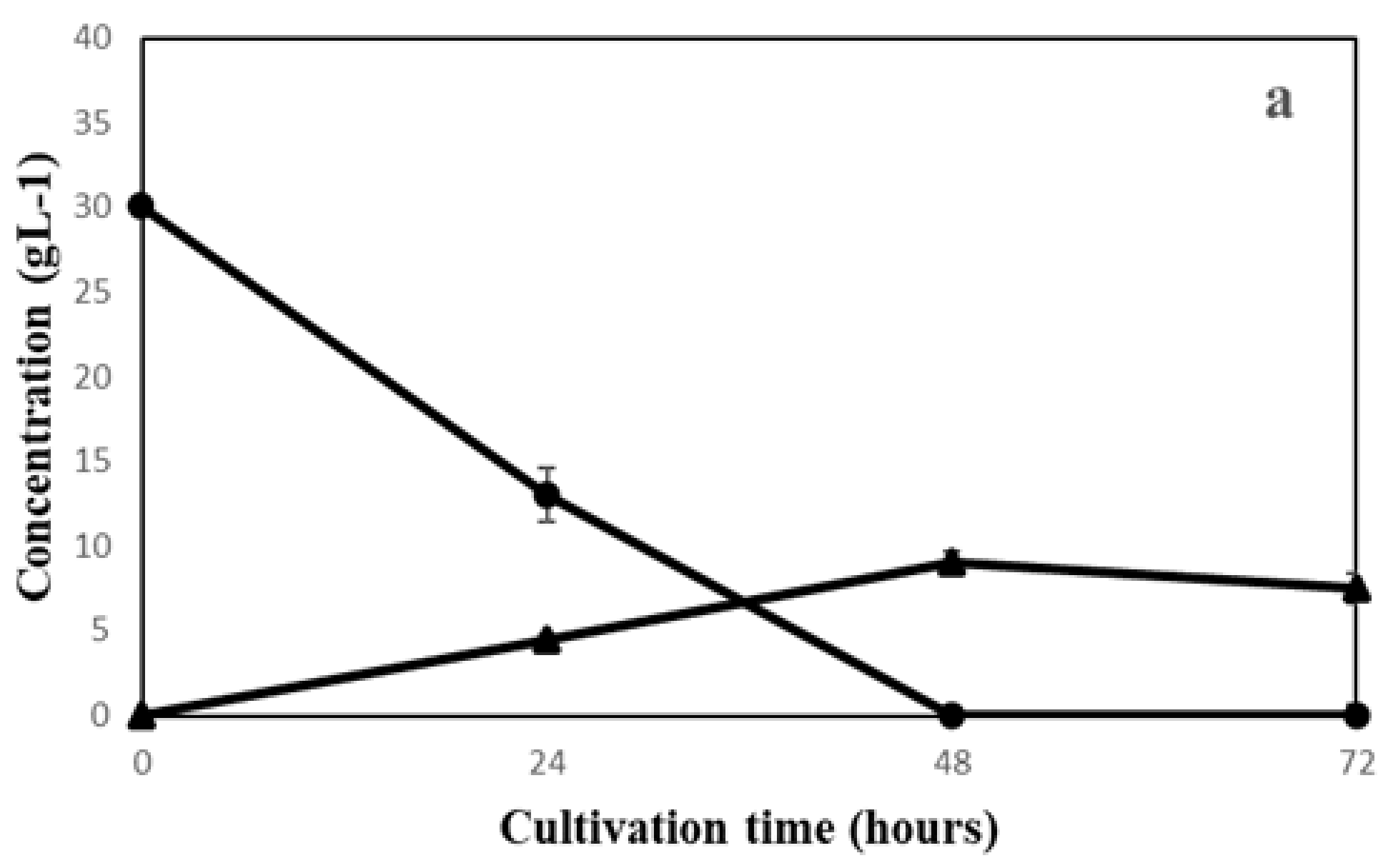
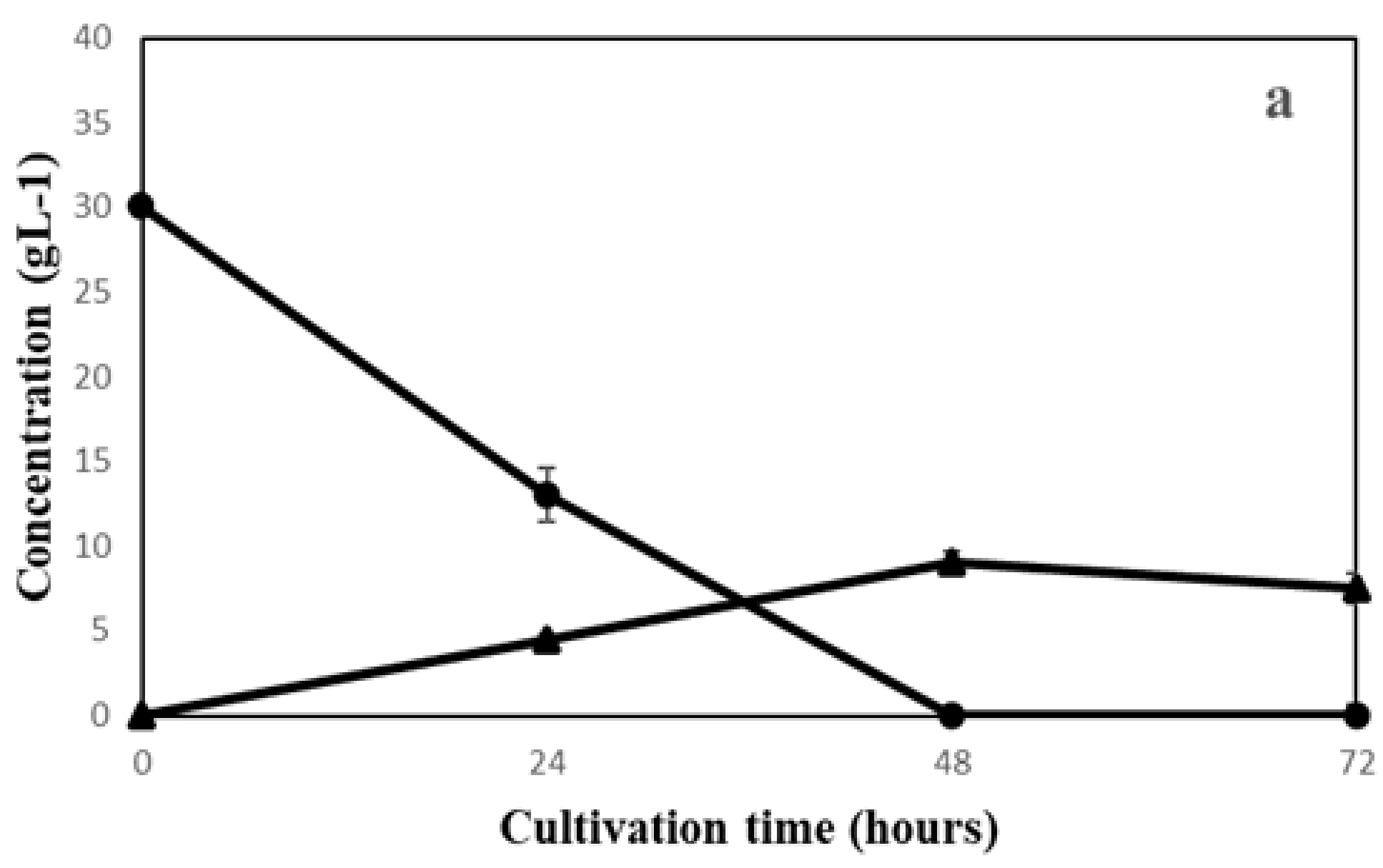
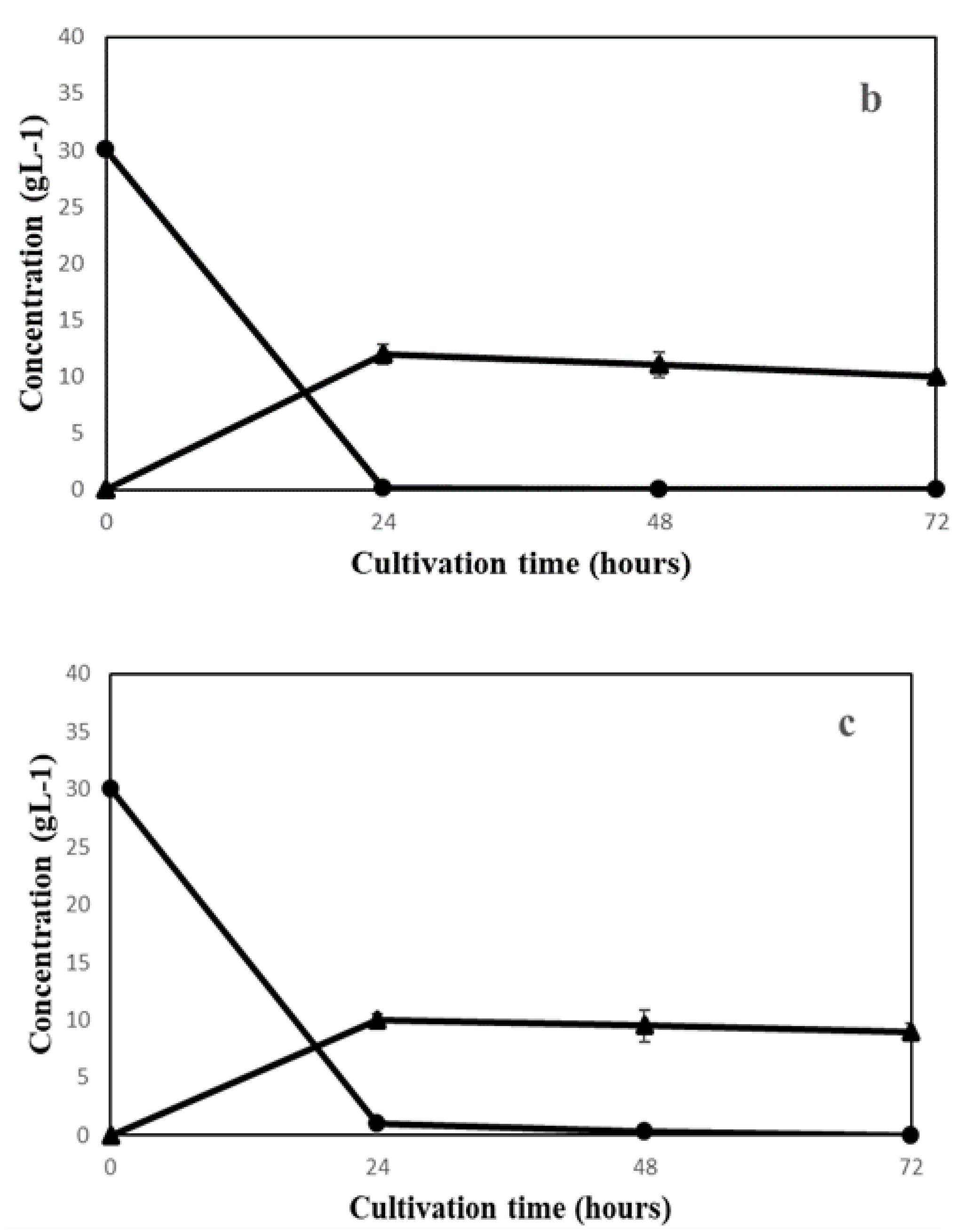
3.1. Filamentous Fungi Cultivation
3.2. Nutritional Properties of Fungal Biomass
3.2.1. Protein
3.2.2. Lipids
3.2.3. Minerals
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations. World Population Prospects: The 2017 Revision, Key Findings and Advance Tables; United Nations: New York, NY, USA, 2017. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2018. [Google Scholar]
- Oliva-Teles, A.; Enes, P.; Peres, H. Replacing fishmeal and fish oil in industrial aquafeeds for carnivorous fish. In Feed and Feeding Practices in Aquaculture; Elsevier BV: Amsterdam, The Netherlands, 2015; pp. 203–233. [Google Scholar]
- Randazzo, B.; Zarantoniello, M.; Cardinaletti, G.; Cerri, R.; Giorgini, E.; Belloni, A.; Contò, M.; Tibaldi, E.; Olivotto, I. Hermetia illucens and poultry by-product meals as alternatives to plant protein sources in gilthead seabream (Sparus aurata) diet: A multidisciplinary study on fish gut status. Animals 2021, 11, 677. [Google Scholar] [CrossRef] [PubMed]
- Tacon, A.G.; Metian, M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture 2008, 285, 146–158. [Google Scholar] [CrossRef]
- Karimi, S.; Ferreira, J.A.; Taherzadeh, M.J. The application of fungal biomass as feed. In Encyclopedia of Mycology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 601–612. [Google Scholar] [CrossRef]
- Karimi, S.; Soofiani, N.M.; Lundh, T.; Mahboubi, A.; Kiessling, A.; Taherzadeh, M.J. Evaluation of filamentous fungal biomass cultivated on vinasse as an alternative nutrient source of fish feed: Protein, lipid, and mineral composition. Fermentation 2019, 5, 99. [Google Scholar] [CrossRef] [Green Version]
- Øverland, M.; Karlsson, A.; Mydland, L.T.; Romarheim, O.H.; Skrede, A. Evaluation of Candida utilis, Kluyveromyces marxianus and Saccharomyces cerevisiae yeasts as protein sources in diets for Atlantic salmon (Salmo salar). Aquaculture 2013, 402, 1–7. [Google Scholar] [CrossRef]
- Gasco, L.; Gai, F.; Maricchiolo, G.; Genovese, L.; Ragonese, S.; Bottari, T.; Caruso, G. Feeds for the Aquaculture Sector—Current Situation and Alternative Sources; Springer International Publishing: New York, NY, USA, 2018. [Google Scholar]
- Nalage, D.; Khedkar, G.; Kalyankar, A.; Sarkate, A.; Ghodke, S.; Bedre, V. Single cell proteins. In Encyclopedia of Food and Health, 1st ed.; Oxford Academic Press: London, UK, 2016; pp. 790–794. [Google Scholar]
- Karimi, S.; Soofiani, N.M.; Mahboubi, A.; Taherzadeh, M.J. Use of organic wastes and industrial by-products to produce filamentous fungi with potential as aqua-feed ingredients. Sustainability 2018, 10, 3296. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.A.; Mahboubi, A.; Lennartsson, P.R.; Taherzadeh, M.J. Waste biorefineries using filamentous ascomycetes fungi: Present status and future prospects. Bioresour. Technol. 2016, 215, 334–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lennartsson, P.R. Zygomycetes and Cellulose Residuals: Hydrolysis, Cultivation and Applications. Ph.D. Thesis, Chalmers Tekniska Högskola, Göteborg, Sweden, 2012. [Google Scholar]
- Cavka, A.; Jönsson, L.J. Comparison of the growth of filamentous fungi and yeasts in lignocellulose-derived media. Biocatal. Agric. Biotechnol. 2014, 3, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Mahboubi, A.; Ferreira, J.A.; Taherzadeh, M.J.; Lennartsson, P.R. Production of fungal biomass for feed, fatty acids, and glycerol by Aspergillus oryzae from fat-rich dairy substrates. Fermentation 2017, 3, 48. [Google Scholar] [CrossRef]
- Halver, J.E.; Hardy, R.W. Fish Nutrition; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of structural carbohydrates and lignin in biomass. In Laboratory Analytical Procedure (LAP); NREL: Golden, CO, USA, 2008. [Google Scholar]
- Langeland, M.; Vidakovic, A.; Vielma, J.; Lindberg, J.E.; Kiessling, A.; Lundh, T. Digestibility of microbial and mussel meal for Arctic charr (Salvelinus alpinus) and Eurasian perch (Perca fluviatilis). Aquac. Nutr. 2016, 22, 485–495. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Appelqvist, L.-Å. Rapid methods of lipid extraction and fatty acid methyl ester preparation for seed and leaf tissue with special remarks on preventing the accumulation of lipid contaminants. Ark. Kemi 1968, 28, 551–570. [Google Scholar]
- NRC. Nutrient Requirements of Fish and Shrimp; 978-0-309-47322-4; The National Academies Press: Washington, DC, USA, 2011; p. 392. [Google Scholar]
- Ovie, S.; Eze, S. Lysine requirement and its effect on the body composition of Oreochromis niloticus fingerlings. J. Fish. Aquat. Sci. 2012, 8, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Zhao, H.; Huang, Y.; Cao, J.; Wang, G.; Sun, Y.; Li, Y. Effects of dietary arginine levels on growth performance, body composition, serum biochemical indices and resistance ability against ammonia-nitrogen stress in juvenile yellow catfish (Pelteobagrus fulvidraco). Anim. Nutr. 2016, 2, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Chen, N.; Qiu, X.; Zhao, M.; Jin, L. Arginine requirement and effect of arginine intake on immunity in largemouth bass, Micropterus salmoides. Aquac. Nutr. 2011, 18, 107–116. [Google Scholar] [CrossRef]
- Azizi, S.; Nematollahi, M.A.; Mojazi Amiri, B.; Vélez, E.; Lutfi Royo, E.; Navarro, I.; Gutierrez, J. Lysine and leucine deficiencies affect myocytes development and IGF signaling in gilthead sea bream (Sparus aurata). PLoS ONE 2016, 11, e0147618. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Mai, K.; Xu, W.; Zhang, Y.; Zhou, H.; Ai, Q. Effects of dietary glutamine on survival, growth performance, activities of digestive enzme, antioxidant status and hypoxia stress resistance of half-smooth tongue sole (Cynoglossus semilaevis Günther) post larvae. Aquaculture 2015, 446, 48–56. [Google Scholar] [CrossRef]
- Mohanty, B.; Mahanty, A.; Ganguly, S.; Sankar, T.V.; Chakraborty, K.; Rangasamy, A.; Sharma, A. Amino acid compositions of 27 food fishes and their importance in clinical nutrition. J. Amino Acids 2014, 2014. [Google Scholar] [CrossRef]
- Bystriansky, J.S.; Frick, N.T.; Ballantyne, J.S. Intermediary metabolism of Arctic char Salvelinus alpinus during short-term salinity exposure. J. Exp. Biol. 2007, 210, 1971–1985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, T.; Matsunari, H.; Sugita, T.; Furuita, H.; Masumoto, T.; Iwashita, Y.; Amano, S.; Suzuki, N. Optimization of the supplemental essential amino acids to a fish meal-free diet based on fermented soybean meal for rainbow trout Oncorhynchus mykiss. Fish. Sci. 2012, 78, 359–366. [Google Scholar] [CrossRef]
- Jones, N.R. The free amino acids of fish. 1-Methylhistidine and β-alanine liberation by skeletal muscle anserinase of codling (Gadus callarias). Biochem. J. 1955, 60, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Tan, X.-H.; Zhou, C.-P.; Yang, Y.-K.; Qi, C.-L.; Zhao, S.-Y.; Lin, H.-Z. Effect of dietary valine levels on the growth performance, feed utilization and immune function of juvenile golden pompano, Trachinotus ovatus. Aquac. Nutr. 2018, 24, 74–82. [Google Scholar] [CrossRef]
- Cowey, C.; Sargent, J. Lipid nutrition in fish. Comp. Biochem. Physiol. Part B Comp. Biochem. 1977, 57, 269–273. [Google Scholar] [CrossRef]
- Wu, P.; Jiang, W.; Hu, K.; Liu, Y.; Jiang, J.; Kuang, S.; Tang, L.; Tang, W.; Zhang, Y.; Zhou, X.; et al. The relationship between dietary methionine and growth, digestion, absorption, and antioxidant status in intestinal and hepatopancreatic tissues of sub-adult grass carp (Ctenopharyngodon idella). J. Anim. Sci. Biotechnol. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meletiadis, J.; Meis, J.F.G.M.; Mouton, J.W.; Verweij, P.E. Analysis of growth characteristics of filamentous fungi in different nutrient media. J. Clin. Microbiol. 2001, 39, 478. [Google Scholar] [CrossRef] [Green Version]
- Sauvant, D.; Perez, J.-M.; Tran, G. Tables of Composition and Nutritional Value of Feed Materials: Pigs, Poultry, Cattle, Sheep, Goats, Rabbits, Horses and Fish; Wageningen Academic Publishers: Wageningen, The Netherlands, 2004. [Google Scholar]
- Randazzo, B.; Zarantoniello, M.; Gioacchini, G.; Giorgini, E.; Truzzi, C.; Notarstefano, V.; Cardinaletti, G.; Huyen, K.T.; Carnevali, O.; Olivotto, I. Can insect-based diets affect zebrafish (Danio rerio) reproduction? A Multidisciplinary Study. Zebrafish 2020, 17, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Elagbar, Z.A.; Naik, R.R.; Shakya, A.K.; Bardaweel, S.K. Fatty Acids Analysis, Antioxidant and Biological Activity of Fixed Oil of Annona muricata L. Seeds. J. Chem. 2016, 2016, 1–6. [Google Scholar] [CrossRef]
- Izquierdo, M.S. Essential fatty acid requirements of cultured marine fish larvae. Aquac. Nutr. 1996, 2, 183–191. [Google Scholar] [CrossRef]
- Sargent, J.; Bell, G.; McEvoy, L.; Tocher, D.; Estévez, A. Recent developments in the essential fatty acid nutrition of fish. Aquaculture 1999, 177, 191–199. [Google Scholar] [CrossRef]
- Tocher, D. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Watanabe, T. Lipid nutrition in fish. Comp. Biochem. Physiol. Part B Comp. Biochem. 1982, 73, 3–15. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Nozzi, V.; Truzzi, C.; Giorgini, E.; Cardinaletti, G.; Freddi, L.; Ratti, S.; Girolametti, F.; Osimani, A.; et al. Physiological responses of Siberian sturgeon (Acipenser baerii) juveniles fed on full-fat insect-based diet in an aquaponic system. Sci. Rep. 2021, 11, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.; Sargent, J.R. Arachidonic acid in aquaculture feeds: Current status and future opportunities. Aquaculture 2003, 218, 491–499. [Google Scholar] [CrossRef]
- Rodriguez, C.; Cejas, J.; Martín, M.V.; Badia, P.; Samper, M.; Lorenzo, A. Influence of n-3 highly unsaturated fatty acid deficiency on the lipid composition of broodstock gilthead seabream (Sparus aurata L.) and on egg quality. Fish. Physiol. Biochem. 1998, 18, 177–187. [Google Scholar] [CrossRef]
- Salze, G.; Tocher, D.R.; Roy, W.J.; A Robertson, D. Egg quality determinants in cod (Gadus morhua L.): Egg performance and lipids in eggs from farmed and wild broodstock. Aquac. Res. 2005, 36, 1488–1499. [Google Scholar] [CrossRef] [Green Version]
- Hamre, K.; Yúfera, M.; Ronnestad, I.; Boglione, C.; Conceicao, L.; Izquierdo, M. Fish larval nutrition and feed formulation: Knowledge gaps and bottlenecks for advances in larval rearing. Rev. Aquac. 2013, 5, S26–S58. [Google Scholar] [CrossRef] [Green Version]
- Kaur, N.; Chugh, V.; Gupta, A.K. Essential fatty acids as functional components of foods—A review. J. Food Sci. Technol. 2014, 51, 2289–2303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norambuena, F.; Morais, S.; Emery, J.A.; Turchini, G. Arachidonic acid and eicosapentaenoic acid metabolism in juvenile atlantic salmon as affected by water temperature. PLoS ONE 2015, 10, e0143622. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, M.; Socorro, J.; Arantzamendi, L.; Hernandez-Cruz, C.M. Recent advances in lipid nutrition in fish larvae. Fish Physiol. Biochem. 2000, 22, 97–107. [Google Scholar] [CrossRef]
- Joint FAO/World Health Organization. Vitamin and Mineral Requirements in Human Nutrition; World Health Organization: Geneva, Switzerland, 2005. [Google Scholar]
- He, F.J.; MacGregor, G.A. Beneficial effects of potassium on human health. Physiol. Plant. 2008, 133, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Fairweather-Tait, S.J.; Cashman, K. Minerals and trace elements. Nutr. Prim. Care Provid. 2015, 111, 45–52. [Google Scholar] [CrossRef]
- Yamauchi, M.; Anderson, J.J.B.; Garner, S.C. Calcium and Phosphorus in Health and Disease; CRC Press: New York, NY, USA, 1996. [Google Scholar]
- Takeda, E.; Yamamoto, H.; Yamanaka-Okumura, H.; Taketani, Y. Dietary phosphorus in bone health and quality of life. Nutr. Rev. 2012, 70, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.; Gatlin, D.M., III. Dietary mineral requirement of fish and marine crustaceans. Rev. Fisher. Sci. 1996, 4, 75–99. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Amino Acid (gKg−1 Dry Biomass) | Amino Acid (% of Total AA Content) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A. oryzae | N. inermedia | R. oryzae | Fish Meal | Soybean Meal | A. oryzae | N. inermedia | R. oryzae | Fish Meal | Soybean Meal | Fish Requirement | ||
| Dry matter (%) | 93.6 ± 0.5 | 94.4 ± 0.4 | 93.6 ± 0.0 | 94.3 | 87.6 | |||||||
| Crude protein (%) | 45.7 ± 0.8 c | 62.2 ± 1.2 a | 50.6 ± 2.0 b | 62.6 | 43.3 | |||||||
| Crude fat (%) | 6.9 ± 0.5 b | 4.0 ± 0.7 c | 17.2 ± 1.1 a | 8.9 | 1.7 | |||||||
| Indispensable amino acids | ||||||||||||
| Arginine | 25.3 | 33.9 | 18.1 | 38.2 | 32.0 | 7.58 | 7.18 | 5.87 | 6.75 | 7.81 | 3.5–4 | |
| Histidine | 8.79 | 13.2 | 9.0 | 15.9 | 11.5 | 2.63 | 2.80 | 2.93 | 2.81 | 2.81 | 1–1.2 | |
| Isoleucine | 14.7 | 21.5 | 16.3 | 25.0 | 19.9 | 4.40 | 4.56 | 5.29 | 4.42 | 4.85 | 1.5–2.8 | |
| Leucine | 24.4 | 37.0 | 25.4 | 44.5 | 31.9 | 7.31 | 7.84 | 8.24 | 7.86 | 7.78 | 2.3–2.9 | |
| Lysine | 27.1 | 40.2 | 25.3 | 46.4 | 26.6 | 8.12 | 8.52 | 8.20 | 8.20 | 6.49 | 3–8.4 | |
| Methionine | 5.8 | 9.1 | 6.7 | 16.3 | 6.2 | 1.76 | 1.93 | 2.18 | 2.88 | 1.51 | 1.5–1.9 | |
| Phenylalanine | 14.7 | 20.7 | 15.9 | 24.3 | 21.7 | 4.40 | 4.39 | 5.16 | 4.29 | 5.29 | 2 | |
| Threonine | 16.9 | 24.3 | 16.1 | 25.6 | 17.7 | 5.06 | 5.15 | 5.22 | 4.52 | 4.32 | 2.6 | |
| Valine | 18.6 | 27.2 | 18.4 | 30.0 | 20.8 | 5.57 | 5.76 | 5.97 | 5.30 | 5.07 | 1.7–3.4 | |
| Sum | 156.3 | 227.1 | 151.2 | 266.2 | 188.3 | |||||||
| Dispensable amino acids | ||||||||||||
| Alanine | 26.0 | 32.6 | 23.6 | 39.7 | 19.0 | 7.79 | 6.91 | 7.65 | 7.02 | 4.64 | ||
| Aspargine | 32.9 | 48.4 | 33.3 | 57.7 | 49.0 | 9.86 | 10.26 | 10.80 | 10.20 | 11.95 | ||
| Cysteine | 3.6 | 6.3 | 3.3 | 5.1 | 0.5 | 1.10 | 1.35 | 1.10 | 0.90 | 0.12 | ||
| Glutamine | 51.0 | 68.9 | 41.6 | 77.3 | 77.0 | 15.28 | 14.60 | 13.49 | 13.66 | 18.79 | ||
| Glycine | 16.3 | 26.1 | 15.1 | 48.0 | 18.1 | 4.88 | 5.53 | 4.90 | 8.48 | 4.42 | ||
| Hydroxyproline | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ||
| Ornithine | 2.2 | 1.5 | ND | ND | ND | 0.67 | 0.32 | ND | 0.00 | 0.00 | ||
| Proline | 14.5 | 18.1 | 9.75 | 28.3 | 21.6 | 4.34 | 3.84 | 3.16 | 5.00 | 5.27 | ||
| Serine | 16.5 | 23.3 | 15.1 | 24.5 | 21.8 | 4.94 | 4.94 | 4.90 | 4.33 | 5.32 | ||
| Tyrosine | 14.3 | 19.5 | 15.3 | 19.0 | 14.6 | 4.28 | 4.13 | 4.96 | 3.36 | 3.56 | ||
| Sum | 177.4 | 244.8 | 157.1 | 299.6 | 221.6 | |||||||
| A. oryzae | gKg−1 | N. intermedia | gKg−1 | R. oryzae | gKg−1 | Fish Meal (%) | Fish Meal (gkg−1) | Soybean Meal (%) | Soybean Meal (gkg−1) | |
|---|---|---|---|---|---|---|---|---|---|---|
| C14:0 (Myristic acid) | 0.38 | 0.26 | 0.41 | 0.16 | 0.66 | 1.13 | 6.0 | 4.10 | 0.1 | 0.0 |
| C14:1 (Myristolic acid) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | ||||
| C16:0 (Palmitic acid) | 20.07 | 13.84 | 12.61 | 5.04 | 20.33 | 34.96 | 17.8 | 12.30 | 10.5 | 1.3 |
| C16:1 (Palmitoleic acid) | 1.58 | 1.09 | 2.77 | 1.10 | 2.78 | 4.78 | 7.2 | 5.00 | 0.2 | 0.0 |
| C18:0 (Stearic acid) | 5.89 | 4.06 | 2.95 | 1.18 | 5.12 | 8.80 | 3.6 | 2.50 | 3.8 | 0.5 |
| C18:1 (Oleic acid) | 34.36 | 23.70 | 52.31 | 20.92 | 25.44 | 43.75 | 12.3 | 8.50 | 21.7 | 2.8 |
| C18:1 (Vaccenic acid) | 0.15 | 0.10 | 1.20 | 0.48 | 1.05 | 1.80 | ||||
| C18:2 (Linoleic acid) | 35.91 | 24.77 | 25.80 | 10.32 | 37.94 | 65.25 | 2.1 | 1.40 | 53.1 | 6.8 |
| C18:3 (Linolenic acid) | 0.29 | 0.20 | 1.94 | 0.77 | 0.48 | 0.82 | 1.9 | 1.30 | 7.4 | 0.9 |
| C20:0 (Arachidic acid) | 0.31 | 0.21 | 0.00 | 0.00 | 0.00 | 0.00 | ||||
| C20:1 cis-11-Eicosenoicacid | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 6.6 | 4.5 | ||
| C20:2 (cis-11,14-Eicosadienoic acid) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | ||||
| C20:3 (11,14,17-Eicosatrienoic acid) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | ||||
| C20:4 (Arachidonic acid) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.4 | 1.7 | ||
| C20:5n-3 [36] | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 9.0 | 6.2 | ||
| C22:0 (Behenic acid) | 0.32 | 0.22 | 0.00 | 0.00 | 0.00 | 0.00 | ||||
| C22:1 (Erucic acid) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 7.7 | 5.3 | ||
| C24:0 (Lignoceric acid) | 0.75 | 0.51 | 0.00 | 0.00 | 0.00 | 0.00 | ||||
| C24:1 (Nervonic acid) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | ||||
| C22:6n-3 (Docosahexaenoic acid acid) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 6.6 | 4.5 | ||
| Total | 100 | 100.00 | 100.00 | |||||||
| % Saturated fatty acids | 26.34 | 15.97 | 26.11 | |||||||
| % Monounsaturated fatty acids | 36.09 | 56.29 | 29.27 | |||||||
| % Polyunsaturated fatty acids | 36.2 | 27.74 | 38.42 | |||||||
| Polyunsaturated fatty acids/Saturated fatty acids | 1.37:1 | 1.74:1 | 1.47:1 |
| Mineral (gKg−1) | A. oryzae | N. intermedia | R. oryzae | Fish Meal | Soybean Meal |
|---|---|---|---|---|---|
| Ca | 1.0 | 1.7 | 3.0 | 55.4 | 3.4 |
| K | 11.3 | 9.3 | 1.3 | 7.4 | 21.2 |
| P | 12.4 | 17.5 | 21.2 | 31.0 | 6.2 |
| Mg | 0.6 | 1.1 | 0.4 | 2.6 | 2.9 |
| Na | 0.3 | 0.3 | 0.3 | 11.2 | 0.0 |
| S | 4.6 | 4.7 | 3.1 | 7.4 | - |
| Sum | 30.2 | 34.6 | 29.3 | 115 | 33.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karimi, S.; Mahboobi Soofiani, N.; Mahboubi, A.; Ferreira, J.A.; Lundh, T.; Kiessling, A.; Taherzadeh, M.J. Evaluation of Nutritional Composition of Pure Filamentous Fungal Biomass as a Novel Ingredient for Fish Feed. Fermentation 2021, 7, 152. https://doi.org/10.3390/fermentation7030152
Karimi S, Mahboobi Soofiani N, Mahboubi A, Ferreira JA, Lundh T, Kiessling A, Taherzadeh MJ. Evaluation of Nutritional Composition of Pure Filamentous Fungal Biomass as a Novel Ingredient for Fish Feed. Fermentation. 2021; 7(3):152. https://doi.org/10.3390/fermentation7030152
Chicago/Turabian StyleKarimi, Sajjad, Nasrollah Mahboobi Soofiani, Amir Mahboubi, Jorge A. Ferreira, Torbjörn Lundh, Anders Kiessling, and Mohammad J. Taherzadeh. 2021. "Evaluation of Nutritional Composition of Pure Filamentous Fungal Biomass as a Novel Ingredient for Fish Feed" Fermentation 7, no. 3: 152. https://doi.org/10.3390/fermentation7030152
APA StyleKarimi, S., Mahboobi Soofiani, N., Mahboubi, A., Ferreira, J. A., Lundh, T., Kiessling, A., & Taherzadeh, M. J. (2021). Evaluation of Nutritional Composition of Pure Filamentous Fungal Biomass as a Novel Ingredient for Fish Feed. Fermentation, 7(3), 152. https://doi.org/10.3390/fermentation7030152