
In Vitro Antioxidant and Antihypertensive Activity of Edible Insects Flours (Mealworm and Grasshopper) Fermented with Lactococcus lactis Strains


 and
and
Abstract
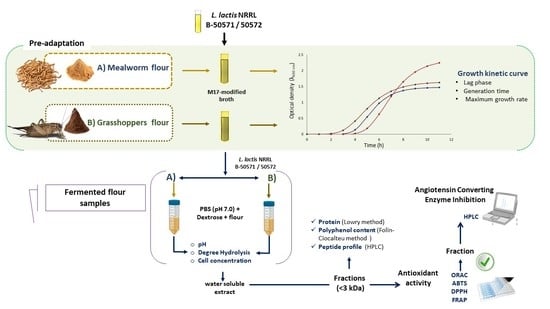
:
1. Introduction
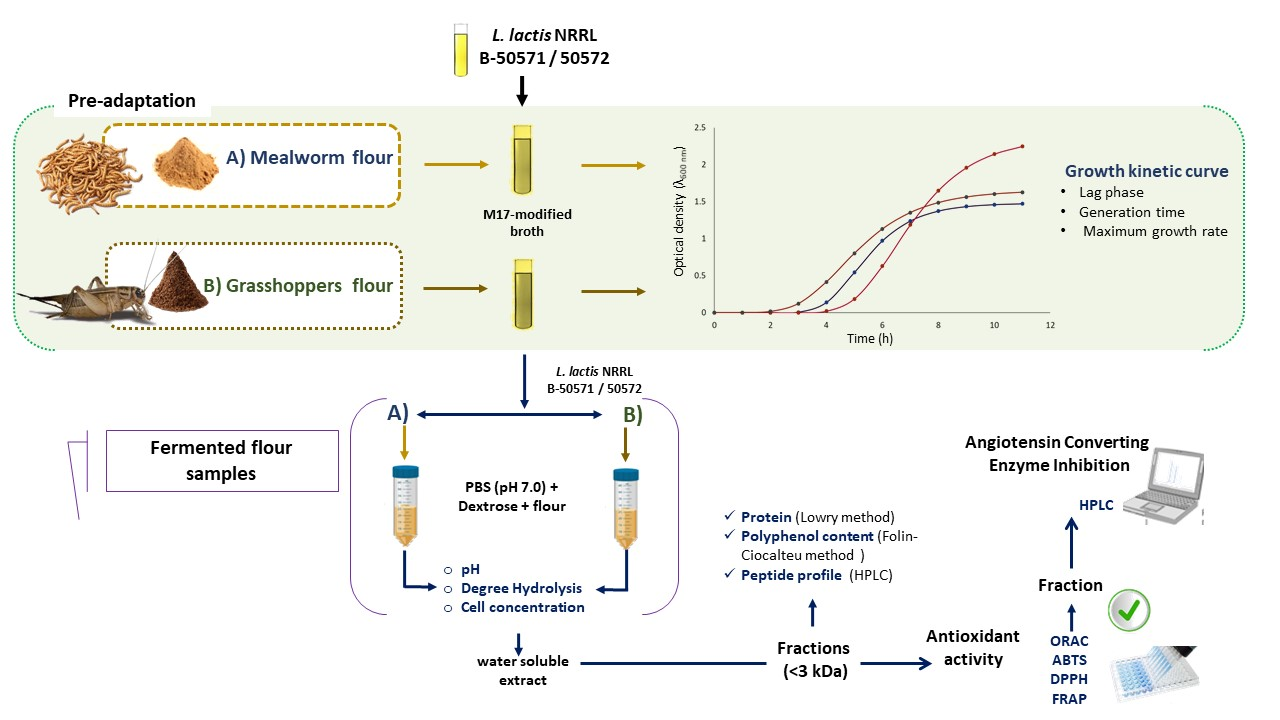
2. Materials and Methods
2.1. Lactic Acid Bacteria Strains and Insect Flours
2.2. Conditions for the Growth of L. lactis
2.3. Fermentation Conditions
2.4. Determination of Degree of Hydrolysis and Protein Content
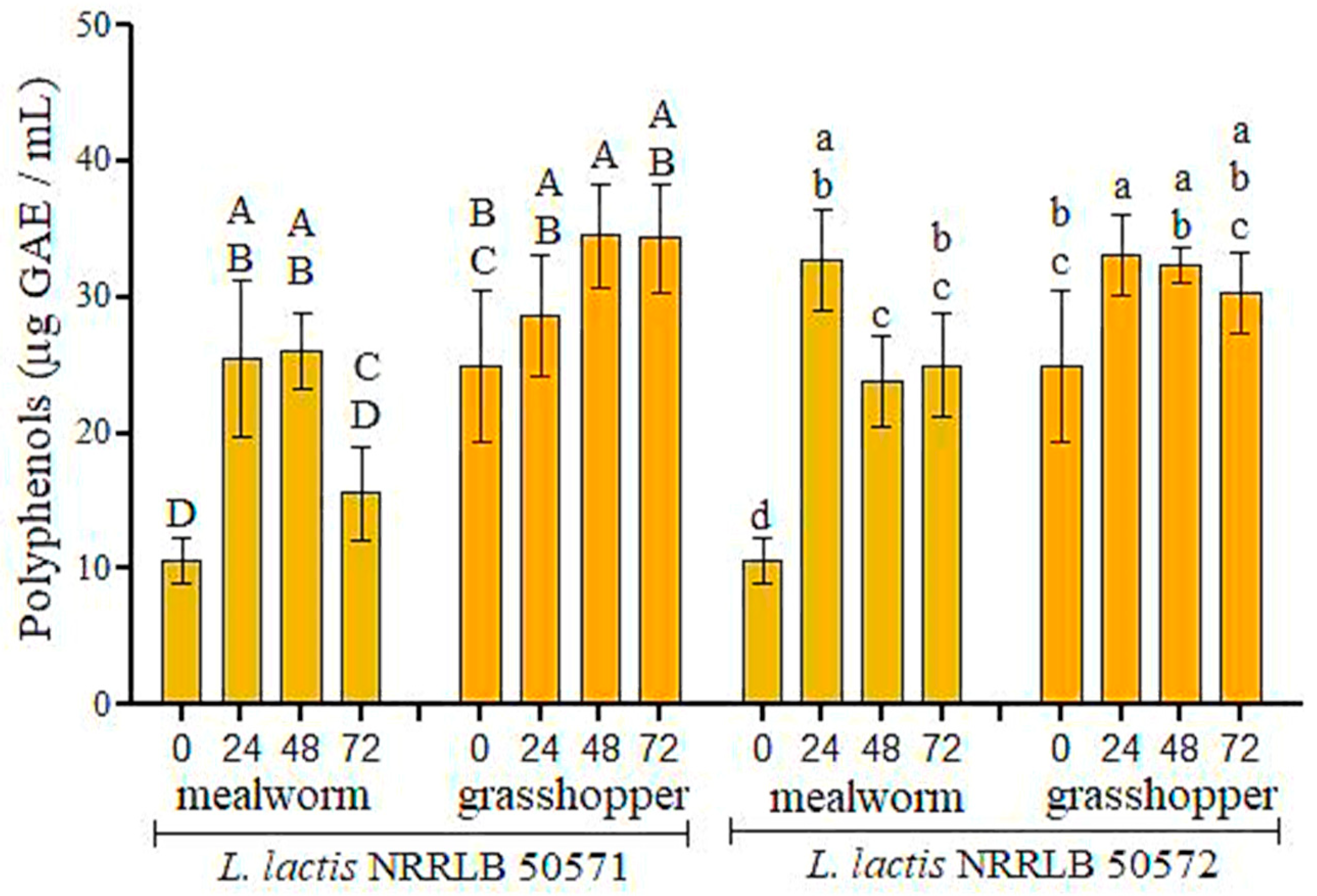
2.5. Determination of Total Polyphenol Content in Fermented Insect Flours
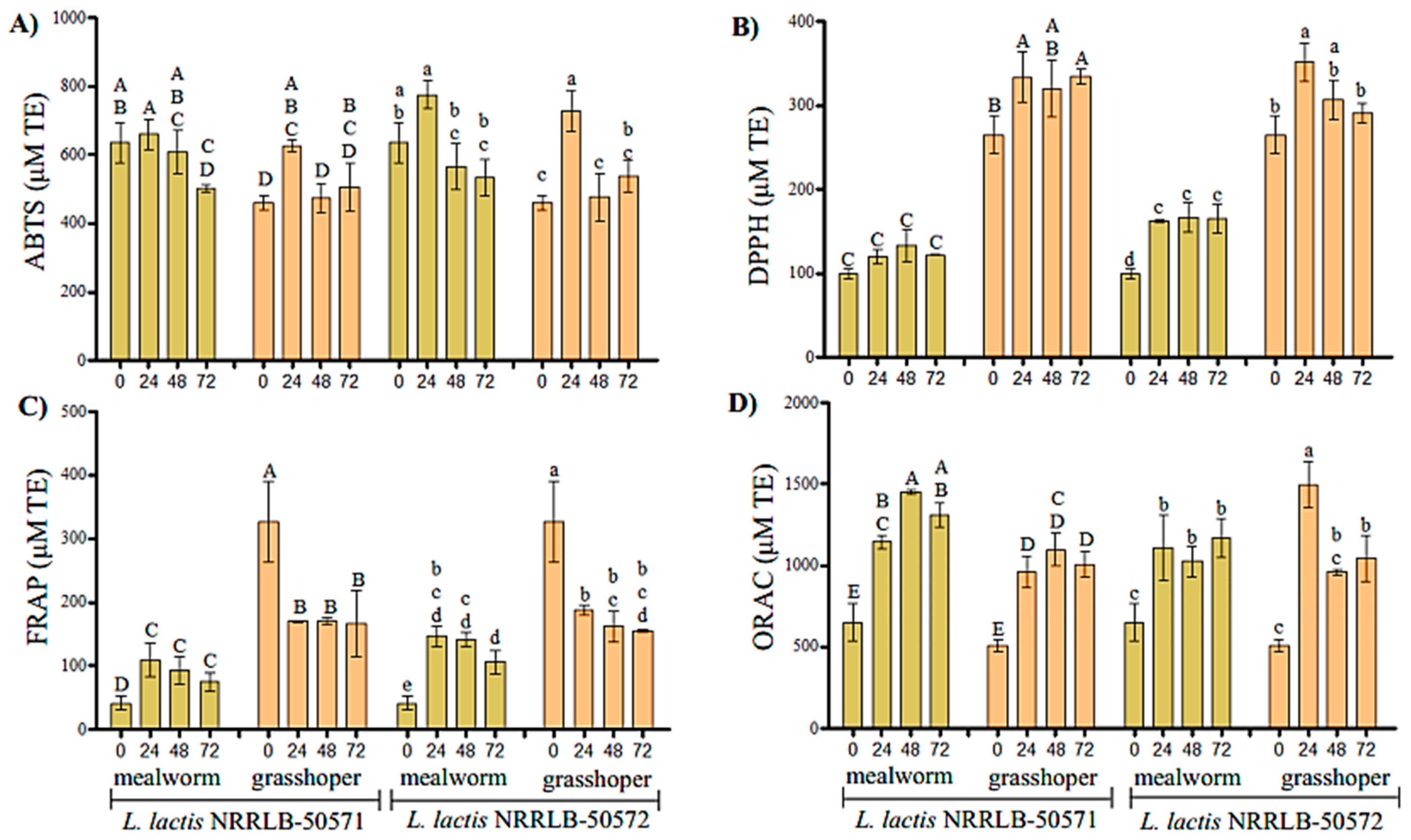
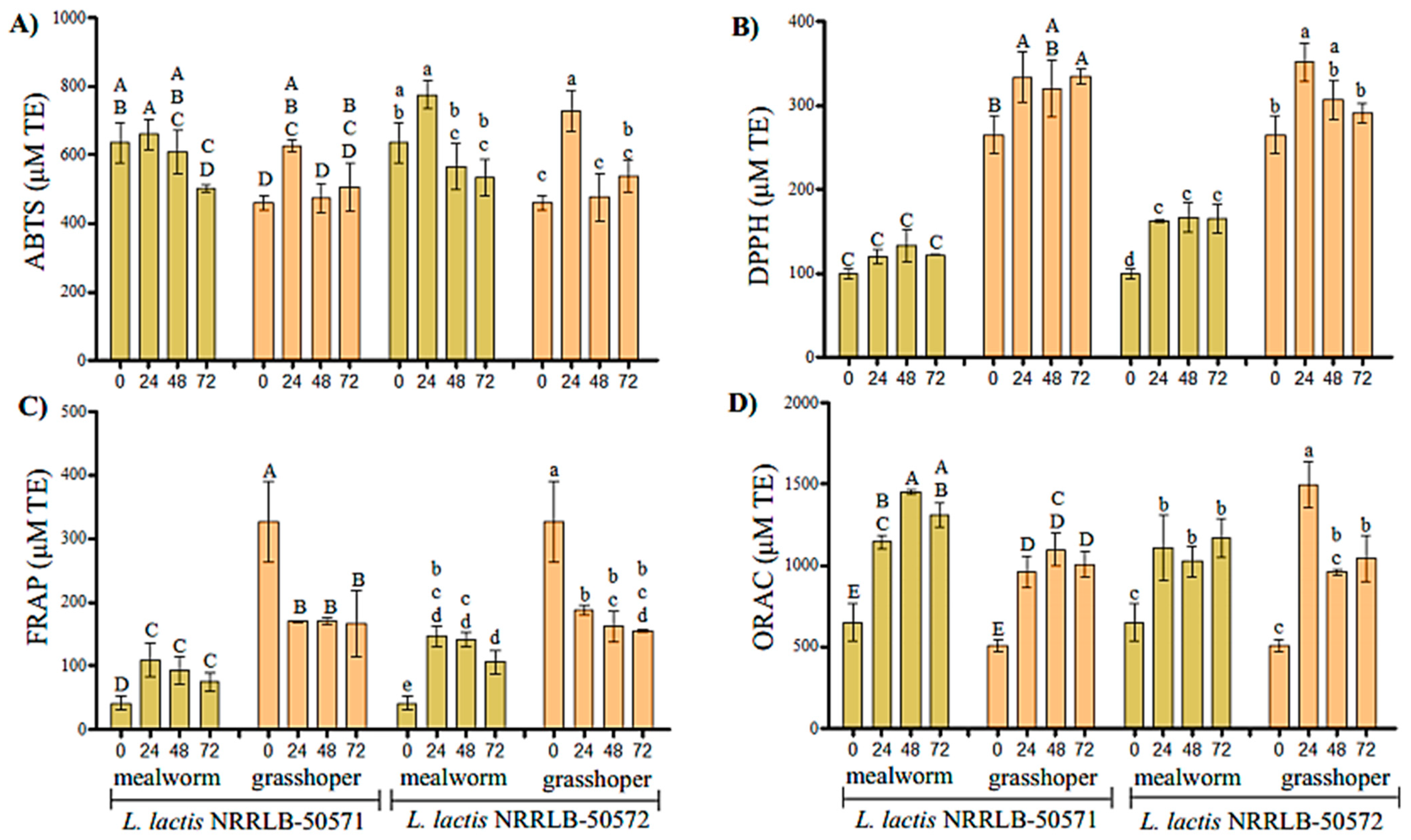
2.6. Determination of Antioxidant Activity
2.7. Determination of Angiotensin Converting Enzyme Inhibition
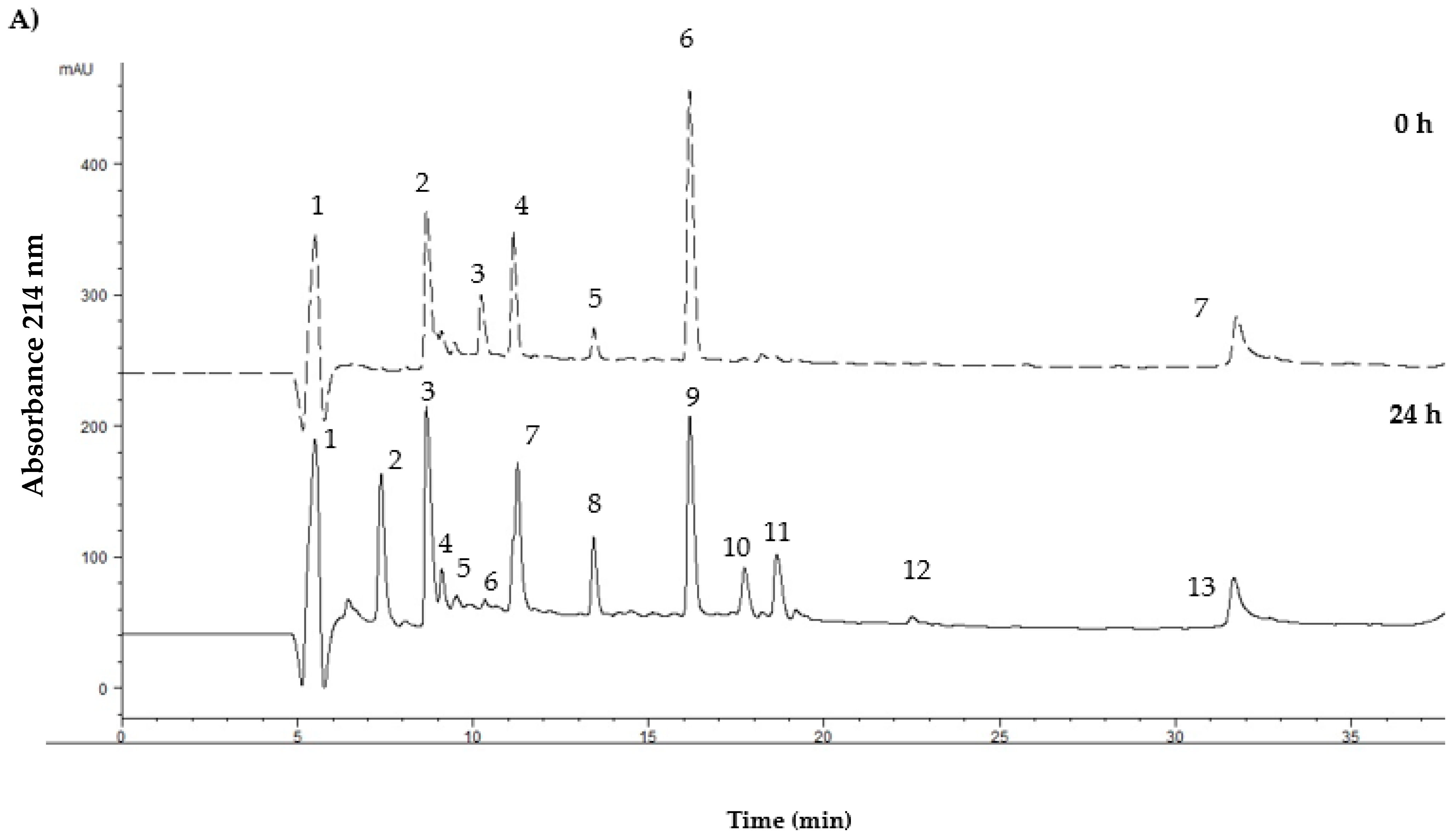
2.8. Isolation of Peptide Fractions by Reversed-Phase HPLC
2.9. Statistical Analysis
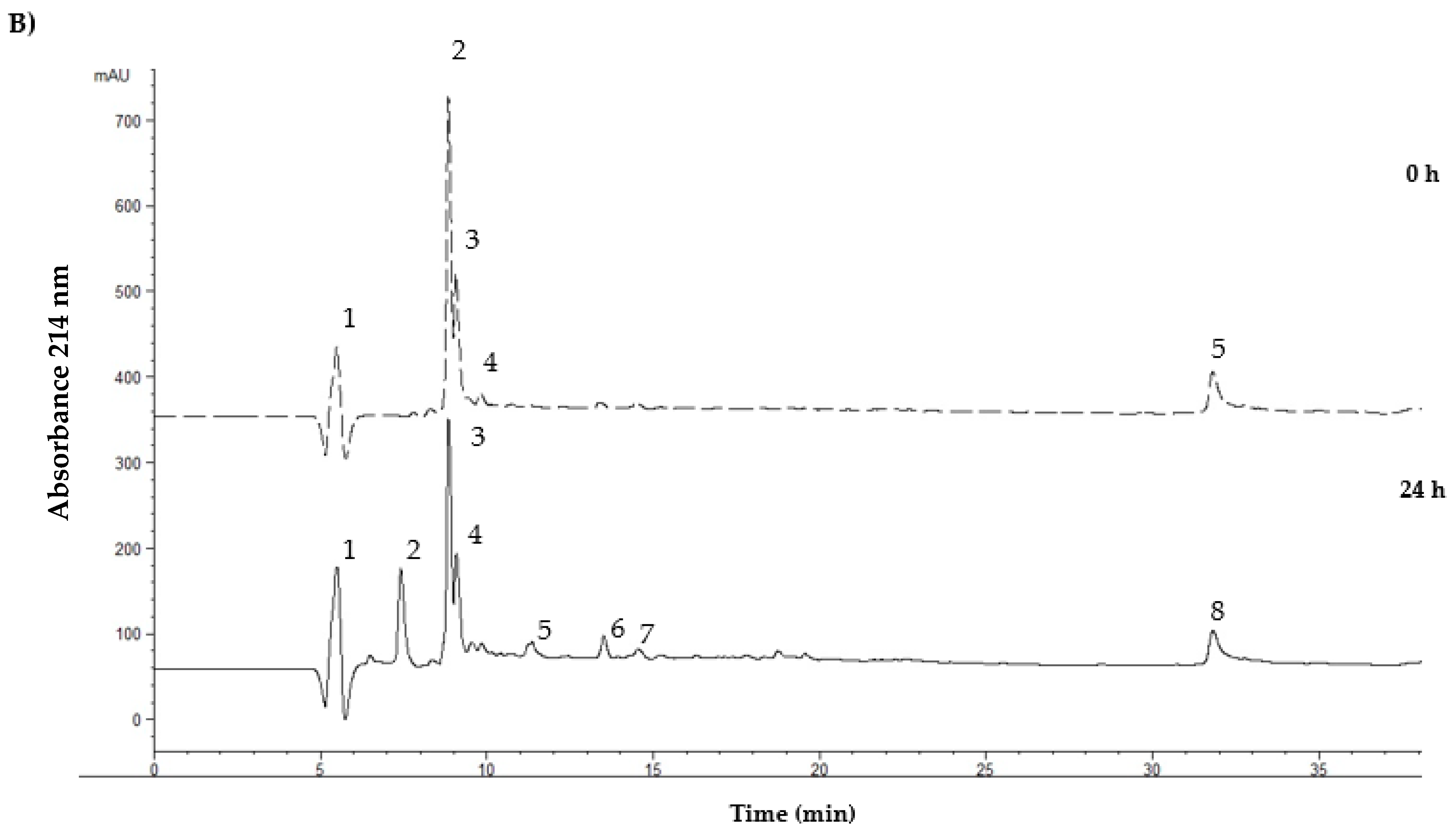
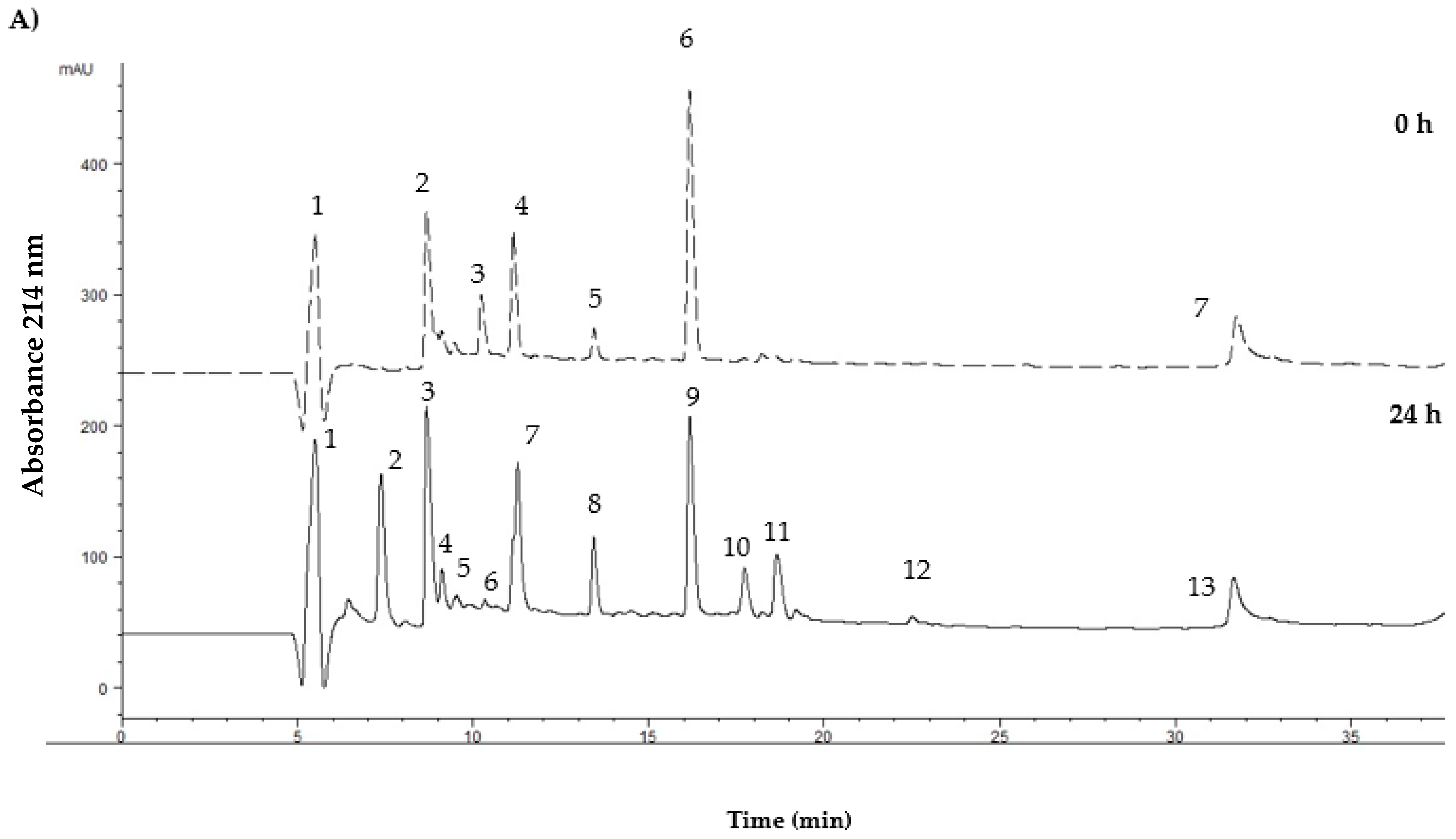
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Imathiu, S. Benefits and food safety concerns associated with consumption of edible insects. NFS J. 2020, 18, 1–11. [Google Scholar] [CrossRef]
- van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed. A Review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef] [Green Version]
- Govorushko, S. Global status of insects as food and feed source: A review. Trends Food Sci. Technol. 2019, 91, 436–445. [Google Scholar] [CrossRef]
- Tang, C.; Yang, D.; Liao, H.; Liu, C.; Wei, L.; Li, F. Edible insects as a food source: A review. Food Prod. Process. Nutr. 2019, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Oonincx, D.G.; van Itterbeeck, J.; Heetkamp, M.J.; van den Brand, H.; van Loon, J.J.; van Huis, A. An exploration on greenhouse gas and ammonia production by insect species suitable for animal or human consumption. PLoS ONE 2010, 5, e14445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-López, C.; Santiago-López, L.; Vallejo-Cordoba, B.; González-Córdova, A.F.; Liceaga, A.M.; García, H.S.; Hernández-Mendoza, A. An insight to fermented edible insects: A global perspective and prospective. Food Res. Int. 2020, 137, 109750. [Google Scholar] [CrossRef] [PubMed]
- da Silva Lucas, A.J.; de Oliveira, L.M.; da Rocha, M.; Prentice, C. Edible insects: An alternative of nutritional, functional and bioactive Compounds. Food Chem. 2020, 311, 126022. [Google Scholar] [CrossRef]
- Stull, V.J.; Finer, E.; Bergmans, R.S.; Febvre, H.P.; Longhurst, C.; Manter, D.K. Impact of edible cricket consumption on gut microbiota in healthy adults, a double-blind, randomized crossover trial. Sci. Rep. 2018, 8, 10762. [Google Scholar] [CrossRef] [Green Version]
- Vercruysse, L.; Smagghe, G.; Beckers, T.; Van Camp, J. Antioxidative and ACE inhibitory activities in enzymatic hydrolysates of the cotton leafworm, Spodoptera littoralis. Food Chem. 2009, 114, 38–43. [Google Scholar] [CrossRef]
- Tao, M.; Wang, C.; Liao, D.; Liu, H.; Zhao, Z.; Zhao, Z. Purification, modification and inhibition mechanism of angiotensin I-converting enzyme inhibitory peptide from silkworm pupa (Bombyx mori) protein hydrolysate. Process. Biochem. 2017, 54, 172–179. [Google Scholar] [CrossRef]
- Hall, F.; Johnson, P.E.; Liceaga, A. Effect of enzymatic hydrolysis on bioactive properties and allergenicity of cricket (Gryllodes sigillatus) protein. Food Chem. 2018, 262, 39–47. [Google Scholar] [CrossRef]
- Mishyna, M.; Martinez, J.J.I.; Chen, J.; Benjamin, O. Extraction, characterization and functional properties of soluble proteins from edible grasshopper (Schistocerca gregaria) and honey bee (Apis mellifera). Food Res. Int. 2019, 116, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, E.; Karaś, M.; Jakubczyk, A.; Zieliński, D.; Baraniak, B. Edible Insects as Source of Proteins. Bioact. Mol. Food 2018, 1–53. [Google Scholar]
- Hall, F.; Reddivary, L.; Liceaga, A.M. Identification and characterization of edible cricket peptides on hypertensive and glycemic in vitro inhibition and their anti-inflammatory activity on RAW 264.7 macrophage cells. Nutrients 2020, 12, 3588. [Google Scholar] [CrossRef] [PubMed]
- de Castro, R.J.S.; Ohara, A.; dos Santos Aguilar, J.G.; Domingues, M.A.F. Nutritional, functional and biological properties of insect proteins: Processes for obtaining, consumption and future challenges. Trends Food Sci. Technol. 2018, 76, 82–89. [Google Scholar] [CrossRef]
- Cho, J.H.; Zhao, H.L.; Kim, J.S.; Kim, S.H.; Chung, C.H. Characteristics of fermented seasoning sauces using Tenebrio molitor larvae. Innov. Food Sci. Emerg. Technol. 2018, 45, 186–195. [Google Scholar] [CrossRef]
- Yi, L.; Van Boekel, M.A.; Boeren, S.; Lakemond, C.M. Protein identification and in vitro digestion of fractions from Tenebrio molitor. Eur. Food Res. Technol. 2016, 242, 1285–1297. [Google Scholar] [CrossRef] [Green Version]
- Mouritsen, O.G.; Duelund, L.; Calleja, G.; Frøst, M.B. Flavour of fermented fish, insect, game, and pea sauces: Garum revisited. Int. J. Gastron. Food Sci. 2017, 9, 16–28. [Google Scholar] [CrossRef]
- Kewuyemi, Y.O.; Kesa, H.; Chinma, C.E.; Adebo, O.A. Fermented edible insects for promoting food security in Africa. Insects 2020, 11, 283. [Google Scholar] [CrossRef]
- Beltrán-Barrientos, L.M.; González-Córdova, A.F.; Hernández-Mendoza, A.; Torres-Inguanzo, E.H.; Astiazarán-García, H.; Esparza-Romero, J.; Vallejo-Cordoba, B. Randomized double-blind controlled clinical trial of the blood pressure–lowering effect of fermented milk with Lactococcus lactis: A pilot study. J. Dairy Sci. 2018, 101, 2819–2825. [Google Scholar] [CrossRef] [Green Version]
- Rendon-Rosales, M.Á.; Torres-Llanez, M.J.; González-Córdova, A.F.; Hernández-Mendoza, A.; Mazorra-Manzano, M.A.; Vallejo-Cordoba, B. In Vitro Antithrombotic and Hypocholesterolemic Activities of Milk Fermented with Specific Strains of Lactococcus lactis. Nutrients 2019, 11, 2150. [Google Scholar] [CrossRef] [Green Version]
- Association of Official Analytical Chemists International (AOAC). Official Methods pf Analysis of A.O.A.C. International: Agricultural Chemicals, Contaminants, Drugs; 17 Ed. Dr. George W. Latimer; AOAC: Washington, DC, USA, 2000. [Google Scholar]
- Chowdhury, B.R.; Chakraborty, R.; Chaudhuri, U.R. Validity of modified Gompertz and Logistic models in predicting cell growth of Pediococcus acidilacticiH during the production of bacteriocin pediocin AcH. J. Food Eng. 2007, 80, 1171–1175. [Google Scholar] [CrossRef]
- Nielse, P.M.; Petersen, D.; Dambmann, C. Improve method for determining Food protein degree of hydrolysis. J. Food Sci. 2001, 66, 641–646. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin–Ciocalteau reagent. Methods Enzym. 1999, 299, 152–178. [Google Scholar]
- Zulueta, A.; Esteve, M.J.; Frígola, A. ORAC and TEAC assays comparison to measure the antioxidant capacity of food products. Food Chem. 2009, 114, 310–316. [Google Scholar] [CrossRef]
- Herald, T.J.; Gadgil, P.; Tilley, M. High-throughput micro plate assays for screening flavonoid content and DPPH-scavenging activity in sorghum bran and flour. J. Sci. Food Agric. 2012, 92, 2326–2331. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Aluko, R.E.; Muir, A.D. Improved method for direct high-performance liquid chromatography Assay of angiotensin-converting enzyme-catalyzed reactions. J. Chromatogr. A 2002, 950, 125–130. [Google Scholar] [CrossRef]
- Widyastuti, Y.; Rohmatussolihat; Febrisiantosa, A. The role of lactic acid bacteria in milk fermentation. Food Nutr. Sci. 2014, 5, 435–442. [Google Scholar] [CrossRef] [Green Version]
- An, B.; Sam, C.; Dries, V.; Ruben, S.; Christel, V.; Borght Mik, V.D.; Bart, L.; Leen, V.C. Comparison of Six Commercial Meat Starter Cultures for the Fermentation of Yellow Mealworm (Tenebrio molitor) Paste. Microorganisms 2019, 7, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltrán-Barrientos, L.M.; Garcia, H.S.; Reyes-Díaz, R.; Estrada-Montoya, M.C.; Torres-Llanez, M.J.; Hernández-Mendoza, A.; González-Córdova, A.F.; Vallejo-Cordoba, B. Cooperation between Lactococcus lactis NRRL B-50571 and NRRL B-50572 for Aroma Formation in Fermented Milk. Foods 2019, 8, 645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Bayjanov, J.R.; Renckens, B.; Nauta, A.; Siezen, R.J. The proteolytic system of lactic acid bacteria revisited: A genomic comparison. BMC Genom. 2010, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, M.W.; Tellez, A.M. Lactobacillus helveticus: The proteolytic system. Front. Microbiol. 2013, 4, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkar, S.G.; Trower, T.M.; Stevenson, D.E. Fecal microbial metabolism of polyphenols and its effects on human gut microbiota. Anaerobe 2013, 23, 12–19. [Google Scholar] [CrossRef]
- Zhao, D.; Shah, N.P. Lactic acid bacterial fermentation modified phenolic composition in tea extracts and enhanced their antioxidant activity and cellular uptake of phenolic compounds following in vitro digestion. J. Funct. Foods 2016, 20, 182–194. [Google Scholar] [CrossRef]
- Di Mattia, C.; Battista, N.; Sacchetti, G.; Serafini, M. Antioxidant Activities in vitro of Water and Liposoluble Extracts Obtained by Different Species of Edible Insects and Invertebrates. Front. Nutr. 2019, 6, 106. [Google Scholar] [CrossRef] [Green Version]
- Nino, M.; Reddivari, L.; Osorio, C.; Kaplan, I.; Liceaga, A. Insects as a source of phenolic compounds and potential health benefits. J. Insects Food Feed. 2021, in press. [Google Scholar] [CrossRef]
- Burghardt, F.; Fiedlert, K.; Proksch, P. Uptake of flavonoids from Vicia villosa (Fabaceae) by the lycaenid butterfly, Polyommatus icarus (Lepidoptera: Lycaenidae). Biochem. Syst. Ecol. 1997, 25, 527–536. [Google Scholar] [CrossRef]
- Wiesen, B.; Krug, E.; Fiedler, K.; Wray, V.; Proksch, P. Sequestration of host-plant-derived flavonoids by lycaenid butterfly Polyommatus icarus. J. Chem. Ecol. 1994, 20, 2523–2538. [Google Scholar] [CrossRef] [PubMed]
- Everette, J.D.; Bryant, Q.M.; Green, A.M.; Abbey, Y.A.; Wangila, G.W.; Walker, R.B. A thorough of reactivity of various compounds classes towards the Folin-Ciocalteun reagent. J. Agric. Food Chem. 2010, 58, 8139–8144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikawa, M.; Schafer, T.; Dollard, C.; Sasner, J. Utilization of Folin-Ciocalteu reagent for the detection of certain nitrogen compounds. J. Agric. Food Chem. 2003, 51, 1811–1815. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh-Fokou, P.; Azzini, E.; Peluso, I.; et al. Lifestyle, oxidative stress, and antioxidants: Back and forth in the pathophysiology of chronic diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Zielińska, E.; Karaś, M.; Jakubczyk, A. Antioxidant activity of predigested protein obtained from a range of farmed edible insects. Int. J. Food Sci. 2017, 52, 306–312. [Google Scholar] [CrossRef]
- You, L.; Zheng, L.; Regenstein, J.M.; Zhao, M.; Liu, D. Effect of thermal treatment on the characteristic properties of loach peptide. Int. J. Food Sci. Technol. 2012, 47, 2574–2581. [Google Scholar] [CrossRef]
- Navarro del Hierro, J.; Gutiérrez-Docio, A.; Otero, P.; Reglero, G.; Martin, D. Characterization, antioxidant activity, and inhibitory effect on pancreatic lipase of extracts from the edible insects Acheta domesticus and Tenebrio molitor. Food Chem. 2019, 309, 125742. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Baran, T.; Asan-Ozusaglam, M.; Cakmak, Y.S.; Tozak, K.O.; Mol, A.; Mentes, A.; Sezen, G. Extraction and characterization of chitin and chitosan with antimicrobial and antioxidant activities from cosmopolitan orthoptera species (Insecta). Biotechnol. Bioproc. E. 2015, 20, 168–179. [Google Scholar] [CrossRef]
- Horvath-Szanics, E.; Perjéssy, J.; Klupács, A.; Takács, K.; Nagy, A.; Koppány-Szabó, E.; Hegyi, F.; Németh-Szerdahelyi, E.; Du, M.Y.; Wang, Z.R.; et al. Study of chinitase and chitinolytics activity of Lactobacillus strains. Acta Aliment. 2020, 49, 214–224. [Google Scholar] [CrossRef]
- Daskaya-Dikmen, C.; Yucetepe, A.; Karbancioglu-Guler, F.; Daskaya, H.; Ozcelik, B. Angiotensin-I-Converting Enzyme (ACE)-Inhibitory Peptides from Plants. Nutrients 2017, 9, 316. [Google Scholar] [CrossRef]
- Jia, J.; Wu, Q.; Yan, H.; Gui, Z. Purification and molecular docking study of a novel angiotensin-I converting enzyme (ACE) inhibitory peptide from alcalase hydrolysate of ultrasonic-pretreated silkworm pupa (Bombyx mori) protein. Process. Biochem. 2015, 50, 876–883. [Google Scholar] [CrossRef]
- Wu, Q.; Jia, J.; Yan, H.; Du, J.; Gui, Z. A novel angiotensin-I converting enzyme (ACE) inhibitory peptide from gastrointestinal protease hydrolysate of silkworm pupa (Bombyx mori) protein: Biochemical characterization and molecular docking study. Peptides 2015, 68, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Ma, H.; Luo, L.; Yin, X. Angiotensin I-converting enzyme (ACE) inhibitory peptide derived from Tenebrio molitor (L.) larva protein hydrolysate. Eur. Food Res. Technol. 2013, 236, 681–689. [Google Scholar] [CrossRef]
- Sanchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Jakubczyk, A.; Karaś, M.; Rybczyńska-Tkaczyk, K.; Zielińska, E.; Zieliński, D. Current Trends of Bioactive Peptides—New Sources and Therapeutic Effect. Foods 2020, 9, 846. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L. lactis NRRL B-50571 | ||||||||
|---|---|---|---|---|---|---|---|---|
| Mealworm | Grasshopper | |||||||
| Treatment | Insect Flour (%) | Dextrose (%) | λ (h) | μmax (h) | G (h) | λ (h) | μmax (h) | G (h) |
| A | 0.1 | 3.5 | 2.89 ± 0.14 a | 0.43 ± 0.03 b | 0.69 ± 0.05 a,b | 2.89 ± 0.14 b | 0.44 ± 0.03 a | 0.69 ± 0.05 a |
| B | 0.5 | 2.38 ± 0.07 b | 0.46 ± 0.01 b | 0.55 ± 0.02 a | 2.27 ± 0.29 a,b | 0.46 ± 0.02 a | 0.65 ± 0.02 a | |
| C | 1.0 | 2.69 ± 0.13 a | 0.41 ± 0.01 a,b | 0.71 ± 0.01 b | 2.73 ± 0.13 a | 0.60 ± 0.05 b | 0.50 ± 0.04 b | |
| D | 0.1 | 5.0 | 3.02 ± 0.33 c | 0.50 ± 0.02 c | 0.60 ± 0.08 a,b | 2.60 ± 0.08 a,b | 0.50 ± 0.02 a,b | 0.60 ± 0.02 a,b |
| E | 0.5 | 2.64 ± 0.13 a | 0.36 ± 0.03 a | 0.83 ± 0.06 c | 2.38 ± 0.24 a | 0.47 ± 0.06 a | 0.66 ± 0.09 a | |
| F | 1.0 | 2.45 ± 0.13 a | 0.42 ± 0.03 b | 0.70 ± 0.04 b | 2.65 ± 0.14 a,b | 0.45 ± 0.04 a | 0.67 ± 0.05 a | |
| L. lactis NRRL B-50572 | ||||||||
| A | 0.1 | 3.5 | 4.63 ± 0.43 a | 0.46 ± 0.05 a | 0.66 ± 0.05 a | 4.32 ± 0.46 a | 0.49 ± 0.04 a,b | 0.61 ± 0.04 a,b,c |
| B | 0.5 | 3.43 ± 0.09 b | 0.42 ± 0.04 a | 0.72 ± 0.03 a | 3.96 ± 0.56 a | 0.47 ± 0.04 a | 0.65 ± 0.06 c | |
| C | 1.0 | 4.13 ± 0.61 a | 0.38 ± 0.55 a | 0.72 ± 0.10 a | 4.56 ± 0.34 a | 0.46 ± 0.02 a | 0.65 ± 0.03 c | |
| D | 0.1 | 5.0 | 4.27 ± 0.45 a | 1.35 ± 0.32 c | 0.41 ± 0.06 b | 4.56 ± 0.39 a | 0.60 ± 0.02 b | 0.50 ± 0.02 b |
| E | 0.5 | 4.16 ± 0.30 a | 0.79 ± 0.18 b | 0.39 ± 0.05 b | 4.27 ± 0.23 a | 0.53 ± 0.08 a,b | 0.56 ± 0.07 b,c | |
| F | 1.0 | 3.88 ± 0.45 a | 0.40 ± 0.01 a | 0.97 ± 0.12 c | 4.09 ± 0.31 a | 0.41 ± 0.05 a | 0.63 ± 0.08 c | |
| L. lactis NRRL B-50571 | ||||||||
|---|---|---|---|---|---|---|---|---|
| Mealworm | Grasshopper | |||||||
| Sub-Culture | λ (h) | μmax (h) | G (h) | Cell Concentration (CFU/mL) | λ (h) | μmax (h) | G (h) | Cell Concentration (CFU/mL) |
| 1 | 2.64 ± 0.09 a | 0.46 ± 0.03 a | 0.60 ± 0.03 a | 1.5 × 109 a | 3.96 ± 0.05 a | 0.47 ± 0.07 a | 0.64 ± 0.08 a | 1.1 × 109 a |
| 2 | 2.73 ± 0.08 a | 0.77 ± 0.05 b | 0.38 ± 0.06 b | 1.7 × 109 a | 4.89 ± 0.06 b | 0.57 ± 0.05 a,b | 0.53 ± 0.05 a,b | 2.8 × 109 a |
| 3 | 1.00 ± 0.03 b | 0.29 ± 0.04 c | 1.01 ± 0.09 c | 1.9 × 109 a | 2.94 ± 0.05 c | 0.39 ± 0.05 b | 0.77 ± 0.07 b | 1.2 × 109 a |
| L. lactis NRRL B-50572 | ||||||||
| 1 | 4.16 ± 0.07 a | 0.41± 0.06 a | 0.71 ± 0.08 a | 1.5 × 109 a | 2.64 ± 0.07 a | 0.46 ± 0.08 a | 0.65 ± 0.15 a | 2.4 × 109 a |
| 2 | 2.20 ± 0.03 b | 0.36 ± 0.05 a | 0.82 ± 0.05 a | 2.6 × 109 a | 3.13 ± 0.05 b | 0.69 ± 0.04 b | 0.43 ± 0.02 b | 2.2 × 109 a |
| 3 | 2.03 ± 0.5 b | 0.38± 0.03 a | 0.79 ± 0.06 a | 2.9 × 109 a | 1.54 ± 0.03 c | 0.36 ± 0.05 a | 0.84 ± 0.06 c | 2.5 × 109 a |
| Mealworm | Grasshopper | ||||||
|---|---|---|---|---|---|---|---|
| Strain | Time (h) | pH | DH (%) | Protein (mg/mL) | pH | DH (%) | Protein (mg/mL) |
| NRRL B-50571 | 0 | 7.12 ± 0.08 a | 0.34 ± 0.03 a | 0.21 ± 0.03 a | 6.83 ± 0.02 a | 0.03 ± 0.02 a | 0.20 ± 0.05 a |
| 24 | 3.76 ± 0.02 b | 0.77 ± 0.004 b | 0.47 ± 0.05 b | 4.23 ± 0.01 b | 0.24 ± 0.02 b | 0.40 ± 0.02 b | |
| 48 | 3.65 ± 0.10 b | 0.66 ± 0.12 b | 0.56 ± 0.06 b | 4.10 ± 0.01 c | 0.22 ± 0.01 b | 0.38 ± 0.07 b | |
| 72 | 3.59 ± 0.01 b | 0.52 ± 0.01 c | 0.35 ± 0.01 c | 4.11 ± 0.03 c | 0.22 ± 0.01 b | 0.38 ± 0.03 b | |
| NRRL B-50572 | 0 | 7.12 ± 0.08 A | 0.34 ± 0.03 A | 0.21 ± 0.03 A | 6.83 ± 0.02 A | 0.03 ± 0.01 A | 0.20 ± 0.05 A |
| 24 | 3.97 ± 0.05 B | 0.53 ± 0.05 B | 0.47 ± 0.05 B | 4.11 ± 0.02 B | 0.20 ± 0.02 B | 0.45 ± 0.05 B | |
| 48 | 3.82 ± 0.01 C | 0.55 ± 0.09 B | 0.59 ± 0.13 B | 3.96 ± 0.03 C | 0.20 ± 0.02 B | 0.44 ± 0.06 B | |
| 72 | 3.71 ± 0.02 C | 0.50 ± 0.02 B | 0.32 ± 0.08 A | 3.94 ± 0.01 C | 0.25 ± 0.09 B | 0.37 ± 0.02 B | |
| Cell Concentration (CFU/mL) | |||||
|---|---|---|---|---|---|
| Strain | Insects Fermented | 0 h | 24 h | 48 h | 72 h |
| NRRL B-50571 | mealworm | 8.05 × 107 ± 3.52 × 107 a | 5.80 × 108 ± 8.29 × 107 b | 1.66 × 109 ± 2.10 × 108 c | 9.00 × 107 ± 4.97 × 106 a |
| grasshopper | 8.43 × 107 ± 2.44 × 107 a | 1.88 × 108 ± 8.66 × 106 b | 3.03 × 107 ± 4.51 ×106 c | 3.57 × 106 ± 5.77 × 104 c | |
| NRRL B-50572 | mealworm | 6.13 × 107 ± 9.84 × 106 a | 4.15 × 108 ± 8.66 × 107 b | 1.51 × 108 ± 3.07 × 107 b | 1.07 × 108 ± 1.58 × 107 a |
| grasshopper | 5.88 × 107 ± 9.22 × 106 a | 1.31 × 108 ± 1.89 × 107 b | 1.30 × 108 ± 4.62 ×107 b | 1.36 × 108 ± 3.65 × 107 b | |
| Angiotensin Converting Enzyme Inhibition | |||||
|---|---|---|---|---|---|
| 0 h | 24 h | ||||
| Insects Fermented | % | IC50 (mg/mL) | % | IC50 (mg/mL) | |
| mealworm | ND | ND | 17.25 ± 0.70 | 1.36 ± 0.06 | |
| grasshopper | 10.08 ± 2.79 a | 1.00 ± 0.04 a | 23.47 ± 2.91 b | 0.97 ± 0.05 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendoza-Salazar, A.; Santiago-López, L.; Torres-Llanez, M.J.; Hernández-Mendoza, A.; Vallejo-Cordoba, B.; Liceaga, A.M.; González-Córdova, A.F. In Vitro Antioxidant and Antihypertensive Activity of Edible Insects Flours (Mealworm and Grasshopper) Fermented with Lactococcus lactis Strains. Fermentation 2021, 7, 153. https://doi.org/10.3390/fermentation7030153
Mendoza-Salazar A, Santiago-López L, Torres-Llanez MJ, Hernández-Mendoza A, Vallejo-Cordoba B, Liceaga AM, González-Córdova AF. In Vitro Antioxidant and Antihypertensive Activity of Edible Insects Flours (Mealworm and Grasshopper) Fermented with Lactococcus lactis Strains. Fermentation. 2021; 7(3):153. https://doi.org/10.3390/fermentation7030153
Chicago/Turabian StyleMendoza-Salazar, Adilene, Lourdes Santiago-López, María J. Torres-Llanez, Adrián Hernández-Mendoza, Belinda Vallejo-Cordoba, Andrea M. Liceaga, and Aarón F. González-Córdova. 2021. "In Vitro Antioxidant and Antihypertensive Activity of Edible Insects Flours (Mealworm and Grasshopper) Fermented with Lactococcus lactis Strains" Fermentation 7, no. 3: 153. https://doi.org/10.3390/fermentation7030153
APA StyleMendoza-Salazar, A., Santiago-López, L., Torres-Llanez, M. J., Hernández-Mendoza, A., Vallejo-Cordoba, B., Liceaga, A. M., & González-Córdova, A. F. (2021). In Vitro Antioxidant and Antihypertensive Activity of Edible Insects Flours (Mealworm and Grasshopper) Fermented with Lactococcus lactis Strains. Fermentation, 7(3), 153. https://doi.org/10.3390/fermentation7030153