
Heterogeneous A40926 Self-Resistance Profile in Nonomuraea gerenzanensis Population Informs Strain Improvement

 ,
,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Cultivation Conditions
2.2. A40926 Extraction and Analysis
2.3. Minimal Inhibitory Concentrations (MICs) and Population Analysis Profile (PAP)
2.4. VanYn-Related Activity Measurement
3. Results
3.1. Heterogeneous A40926 Resistance Profile in Nonomuraea gerenzanensis Population
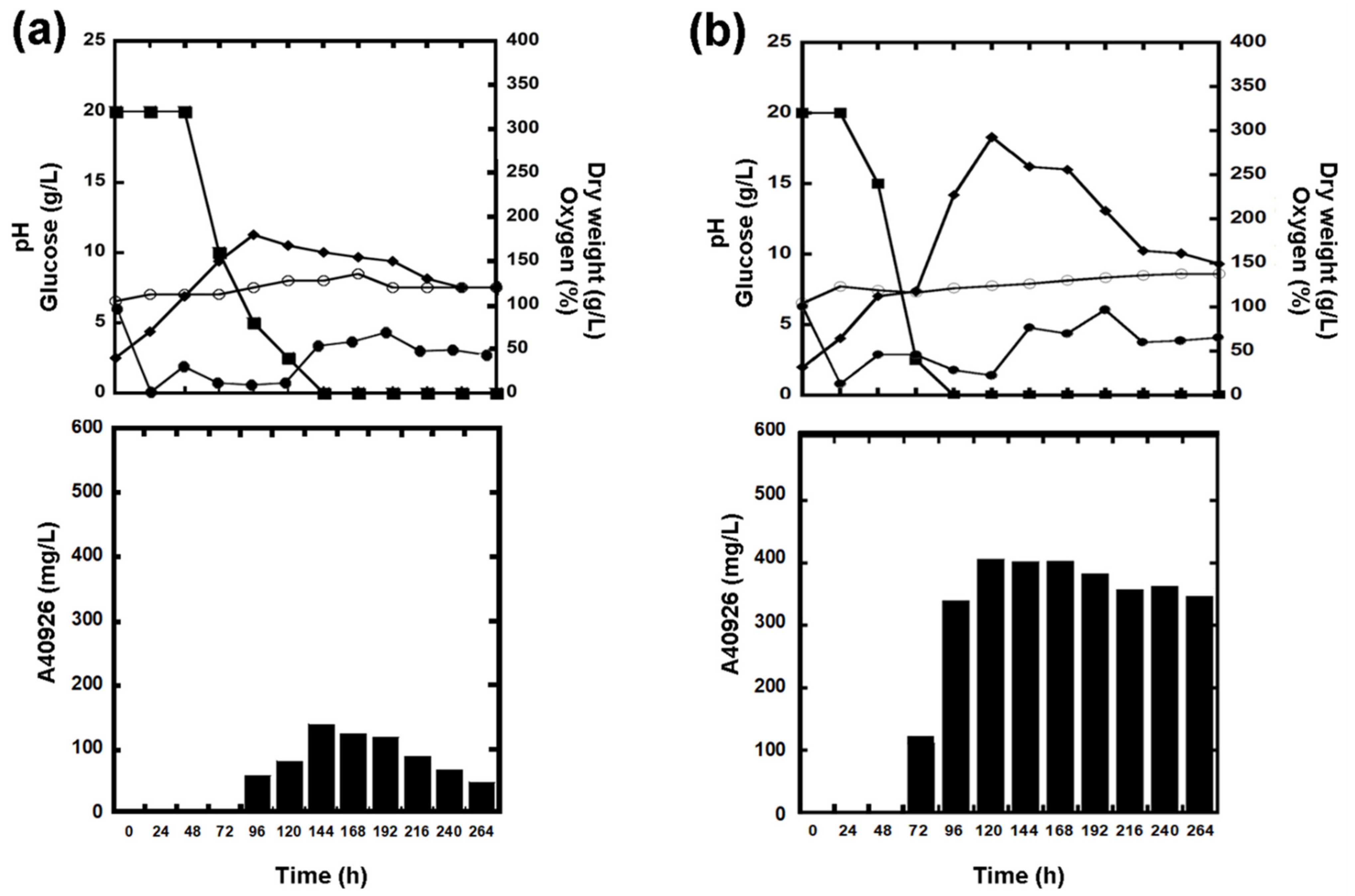
3.2. A40926 Production in N. gerenzanensis and in Its G and P Subpopulations
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goldstein, B.P.; Selva, E.; Gastaldo, L.; Berti, M.; Pallanza, R.; Ripamonti, F.; Ferrari, P.; Denaro, M.; Arioli, V.; Cassani, G. A40926, a new glycopeptide antibiotic with anti-Neisseria activity. Antimicrob. Agents Chemother. 1987, 31, 1961–1966. [Google Scholar] [CrossRef] [PubMed]
- Malabarba, A.; Ciabatti, R.; Scotti, R.; Goldstein, B.P.; Ferrari, P.; Kurz, M.; Andreini, B.P.; Denaro, M. New semisynthetic glycopeptides MDL 63,246 and MDL 63,042, and other amide derivatives of antibiotic A-40,926 active against highly glycopeptide-resistant VanA enterococci. J. Antibiot. 1995, 48, 869–883. [Google Scholar] [CrossRef][Green Version]
- Soriano, A.; Rossolini, G.M.; Pea, F. The role of dalbavancin in the treatment of acute bacterial skin and skin structure infections (ABSSSIs). Expert Rev. Anti. Infect. Ther. 2020, 18, 415–422. [Google Scholar] [CrossRef]
- Marcone, G.L.; Binda, E.; Berini, F.; Marinelli, F. Old and new glycopeptide antibiotics: From product to gene and back in the post-genomic era. Biotechnol. Adv. 2018, 36, 534–554. [Google Scholar] [CrossRef]
- Sosio, M.; Stinchi, S.; Beltrametti, F.; Lazzarini, A.; Donadio, S. The gene cluster for the biosynthesis of the glycopeptide antibiotic A40926 by Nonomuraea species. Chem. Biol. 2003, 10, 541–549. [Google Scholar] [CrossRef]
- Sosio, M.; Canavesi, A.; Stinchi, S.; Donadio, S. Improved production of A40926 by Nonomuraea sp. through deletion of a pathway-specific acetyltransferase. Appl. Microbiol. Biotechnol. 2010, 87, 1633–1638. [Google Scholar] [CrossRef]
- Lo Grasso, L.; Maffioli, S.; Sosio, M.; Bibb, M.; Puglia, A.M.; Alduina, R. Two master switch regulators trigger A40926 biosynthesis in Nonomuraea sp. strain ATCC 39727. J. Bacteriol. 2015, 197, 2536–2544. [Google Scholar] [CrossRef]
- Yushchuk, O.; Andreo-Vidal, A.; Marcone, G.L.; Bibb, M.; Marinelli, F.; Binda, E. New molecular tools for regulation and improvement of A40926 glycopeptide antibiotic production in Nonomuraea gerenzanensis ATCC 39727. Front. Microbiol. 2020, 11, 8. [Google Scholar] [CrossRef] [PubMed]
- Alt, S.; Bernasconi, A.; Sosio, M.; Brunati, C.; Donadio, S.; Maffioli, S.I. Toward single-peak dalbavancin analogs through biology and chemistry. ACS Chem. Biol. 2019, 14, 356–360. [Google Scholar] [CrossRef]
- Alduina, R.; Tocchetti, A.; Costa, S.; Ferraro, C.; Cancemi, P.; Sosio, M.; Donadio, S. A two-component regulatory system with opposite effects on glycopeptide antibiotic biosynthesis and resistance. Sci. Rep. 2020, 10, 6200. [Google Scholar] [CrossRef]
- Marcone, G.L.; Beltrametti, F.; Binda, E.; Carrano, L.; Foulston, L.; Hesketh, A.; Bibb, M.; Marinelli, F. Novel mechanism of glycopeptide resistance in the A40926 producer Nonomuraea sp. ATCC 39727. Antimicrob. Agents Chemother. 2010, 54, 2465–2472. [Google Scholar] [CrossRef]
- Marcone, G.L.; Binda, E.; Carrano, L.; Bibb, M.; Marinelli, F. Relationship between glycopeptide production and resistance in the actinomycete Nonomuraea sp. ATCC 39727. Antimicrob. Agents Chemother. 2014, 58, 5191–5201. [Google Scholar] [CrossRef]
- Binda, E.; Cappelletti, P.; Marinelli, F.; Marcone, G.L. Specificity of induction of glycopeptide antibiotic resistance in the producing actinomycetes. Antibiotics 2018, 7, 36. [Google Scholar] [CrossRef]
- Marcone, G.L.; Carrano, L.; Marinelli, F.; Beltrametti, F. Protoplast preparation and reversion to the normal filamentous growth in antibiotic-producing uncommon actinomycetes. J. Antibiot. 2010, 63, 83–88. [Google Scholar] [CrossRef]
- Beltrametti, F.; Jovetic, S.; Feroggio, M.; Gastaldo, L.; Selva, E.; Marinelli, F. Valine influences production and complex composition of glycopeptide antibiotic A40926 in fermentations of Nonomuraea sp. ATCC 39727. J. Antibiot. 2004, 57, 37–44. [Google Scholar] [CrossRef]
- Kieser, T.; Chater, K.F.; Bibb, M.J.; Buttner, M.J.; Hopwood, D.A. Practical Streptomyces Genetics; John Innes Centre: Norwich, UK, 2000. [Google Scholar]
- Binda, E.; Marcone, G.L.; Berini, F.; Pollegioni, L.; Marinelli, F. Streptomyces spp. as efficient expression system for a d,d-peptidase/d,d-carboxypeptidase involved in glycopeptide antibiotic resistance. BMC Biotechnol. 2013, 13, 24. [Google Scholar] [CrossRef]
- Binda, E.; Marcone, G.L.; Pollegioni, L.; Marinelli, F. Characterization of VanYn, a novel d,d-peptidase/d,d-carboxypeptidase involved in glycopeptide antibiotic resistance in Nonomuraea sp. ATCC 39727. FEBS J. 2012, 279, 3203–3213. [Google Scholar] [CrossRef] [PubMed]
- Sungthong, R.; Nakaew, N. The genus Nonomuraea: A review of a rare actinomycete taxon for novel metabolites. J. Basic Microbiol. 2015, 55, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Yushchuk, O.; Vior, N.M.; Andreo-Vidal, A.; Berini, F.; Rückert, C.; Busche, T.; Binda, E.; Kalinowski, J.; Truman, A.W.; Marinelli, F. Genomic-led discovery of a novel glycopeptide antibiotic by Nonomuraea coxensis DSM 45129. ACS Chem. Biol. 2021, 16, 915–928. [Google Scholar] [CrossRef]
- Lazzarini, A.; Cavaletti, L.; Toppo, G.; Marinelli, F. Rare genera of actinomycetes as potential producers of new antibiotics. Antonie Van Leeuwenhoek 2001, 79, 399–405. [Google Scholar]
- Dalmastri, C.; Gastaldo, L.; Marcone, G.L.; Binda, E.; Congiu, T.; Marinelli, F. Classification of Nonomuraea sp. ATCC 39727, an actinomycete that produces the glycopeptide antibiotic A40926, as Nonomuraea gerenzanensis sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 912–921. [Google Scholar] [CrossRef] [PubMed]
- D’Argenio, V.; Petrillo, M.; Pasanisi, D.; Pagliarulo, C.; Colicchio, R.; Talà, A.; de Biase, M.S.; Zanfardino, M.; Scolamiero, E.; Pagliuca, C.; et al. The complete 12 Mb genome and transcriptome of Nonomuraea gerenzanensis with new insights into its duplicated “magic” RNA polymerase. Sci. Rep. 2016, 6, 18. [Google Scholar] [CrossRef]
- van Bergeijk, D.A.; Terlouw, B.R.; Medema, M.H.; van Wezel, G.P. Ecology and genomics of Actinobacteria: New concepts for natural product discovery. Nat. Rev. Microbiol. 2020, 18, 546–558. [Google Scholar] [CrossRef]
- Volff, J.N.; Altenbuchner, J. Genetic instability of the Streptomyces chromosome. Mol. Microbiol. 1998, 27, 239–246. [Google Scholar] [CrossRef]
- Katz, L.; Baltz, R.H. Natural product discovery: Past, present, and future. J. Ind. Microbiol. Biotechnol. 2016, 43, 155–176. [Google Scholar] [CrossRef]
- Yanai, K.; Murakami, T.; Bibb, M. Amplification of the entire kanamycin biosynthetic gene cluster during empirical strain improvement of Streptomyces kanamyceticus. Proc. Natl. Acad. Sci. USA 2006, 103, 9661–9666. [Google Scholar] [CrossRef]
- Fedorenko, V.; Genilloud, O.; Horbal, L.; Marcone, G.L.; Marinelli, F.; Paitan, Y.; Ron, E.Z. Antibacterial discovery and development: From gene to product and back. Biomed. Res. Int. 2015, 2015, 591349. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Binda, E.; Berini, F.; Marinelli, F.; Bava, A.; Beltrametti, F. Heterogeneous A40926 Self-Resistance Profile in Nonomuraea gerenzanensis Population Informs Strain Improvement. Fermentation 2021, 7, 140. https://doi.org/10.3390/fermentation7030140
Binda E, Berini F, Marinelli F, Bava A, Beltrametti F. Heterogeneous A40926 Self-Resistance Profile in Nonomuraea gerenzanensis Population Informs Strain Improvement. Fermentation. 2021; 7(3):140. https://doi.org/10.3390/fermentation7030140
Chicago/Turabian StyleBinda, Elisa, Francesca Berini, Flavia Marinelli, Adriana Bava, and Fabrizio Beltrametti. 2021. "Heterogeneous A40926 Self-Resistance Profile in Nonomuraea gerenzanensis Population Informs Strain Improvement" Fermentation 7, no. 3: 140. https://doi.org/10.3390/fermentation7030140
APA StyleBinda, E., Berini, F., Marinelli, F., Bava, A., & Beltrametti, F. (2021). Heterogeneous A40926 Self-Resistance Profile in Nonomuraea gerenzanensis Population Informs Strain Improvement. Fermentation, 7(3), 140. https://doi.org/10.3390/fermentation7030140