
Seven New Species of Eurotiales (Ascomycota) Isolated from Tidal Flat Sediments in China




Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Fungal Isolation
2.2. Morphological Observations
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Phylogenetic Analyses
3. Results
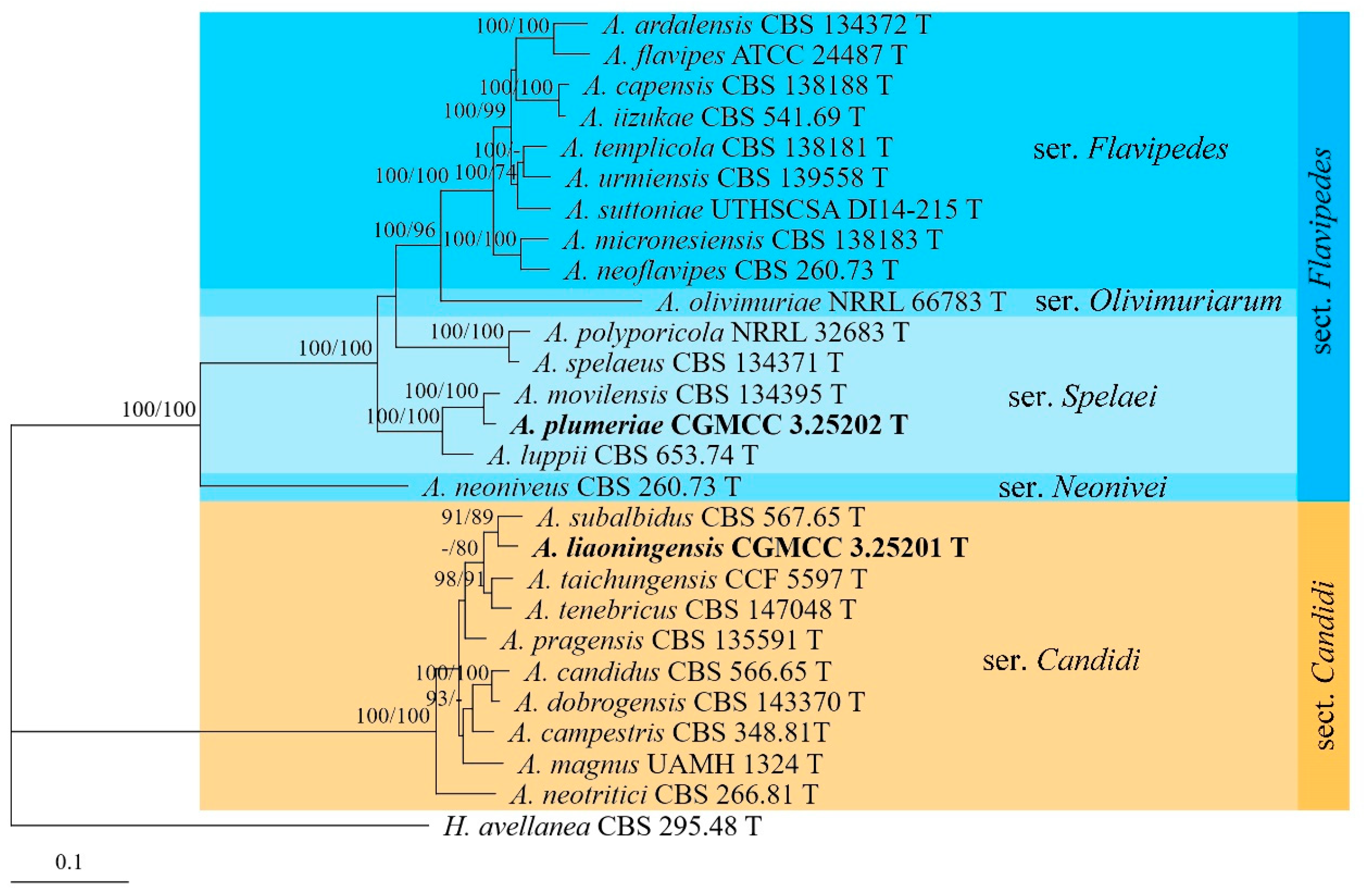
3.1. Phylogenetic Analyses
3.2. Taxonomy
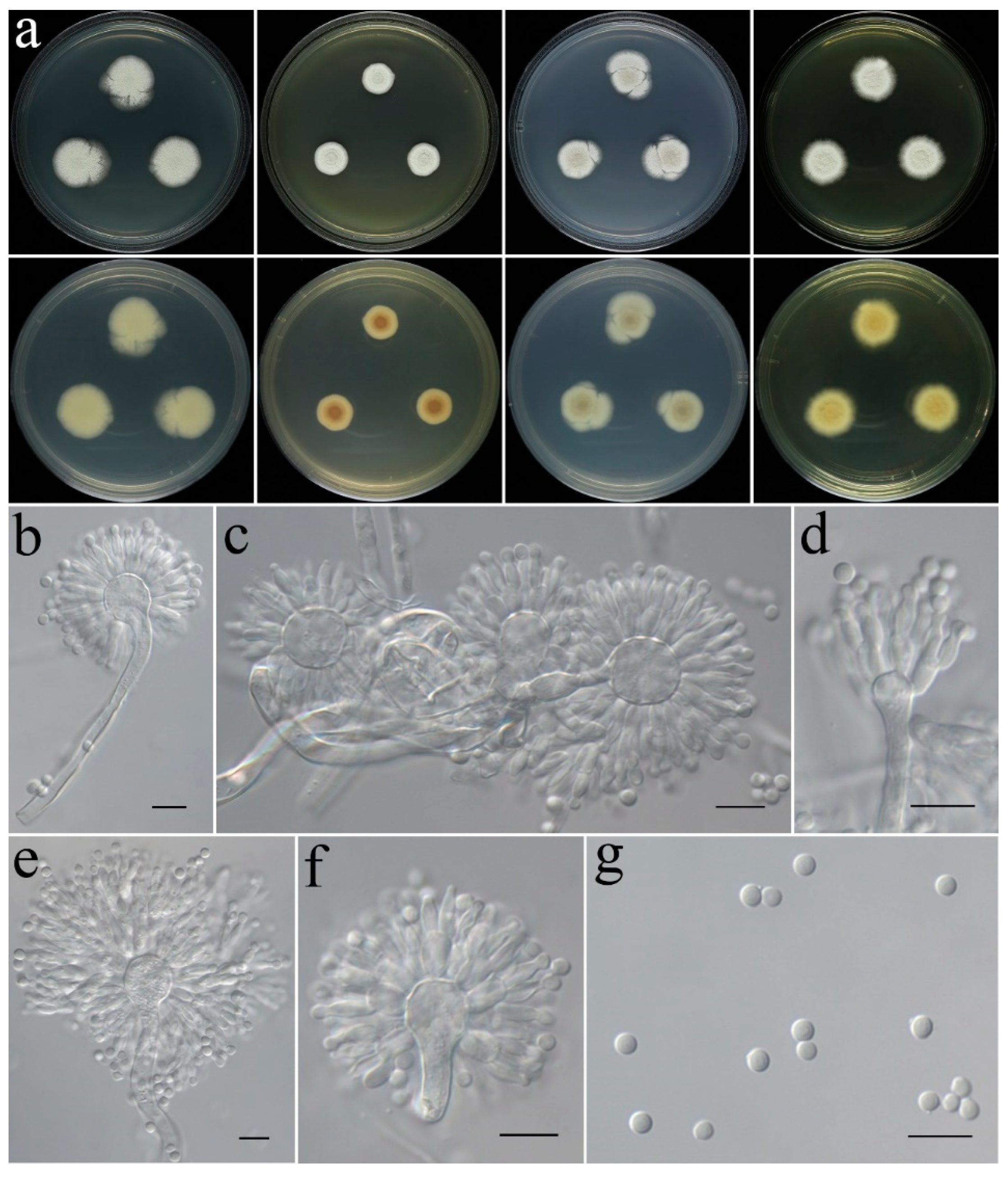
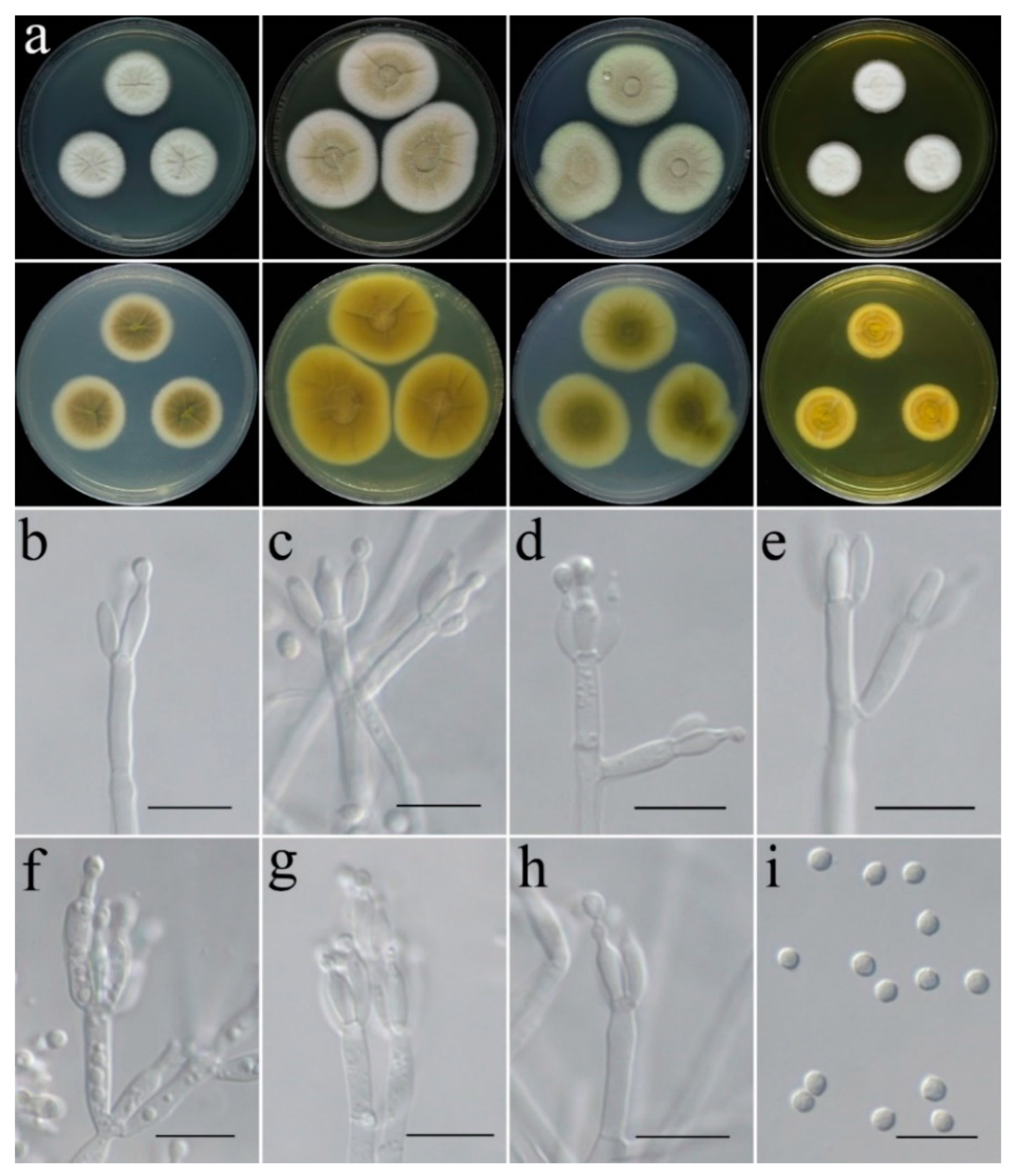
- Aspergillus plumeriae C. Liu, Z.Q. Zeng & W.Y. Zhuang, sp. nov. Figure 7.
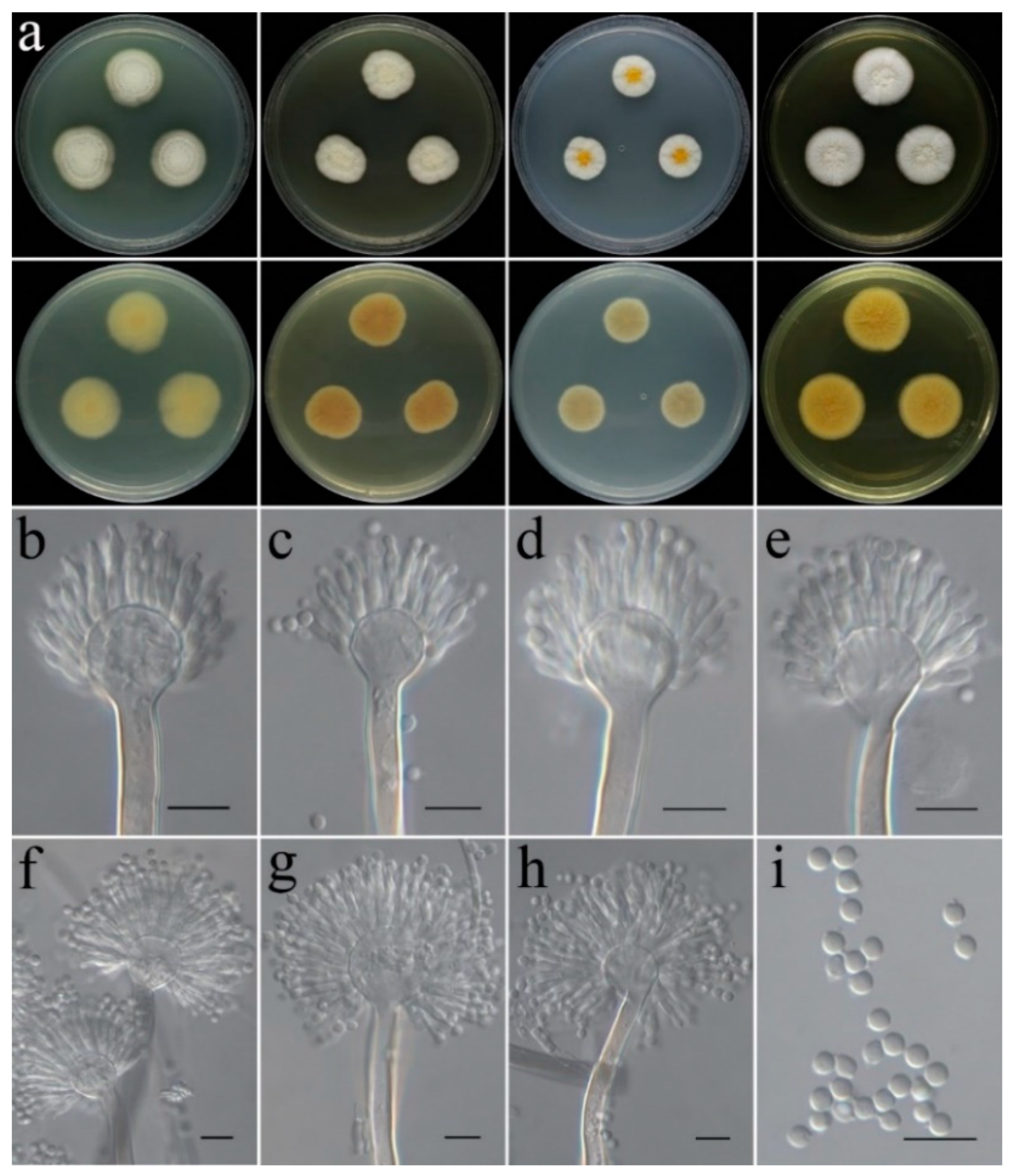
- Aspergillus subinflatus C. Liu, Z.Q. Zeng & W.Y. Zhuang, sp. nov. Figure 8.
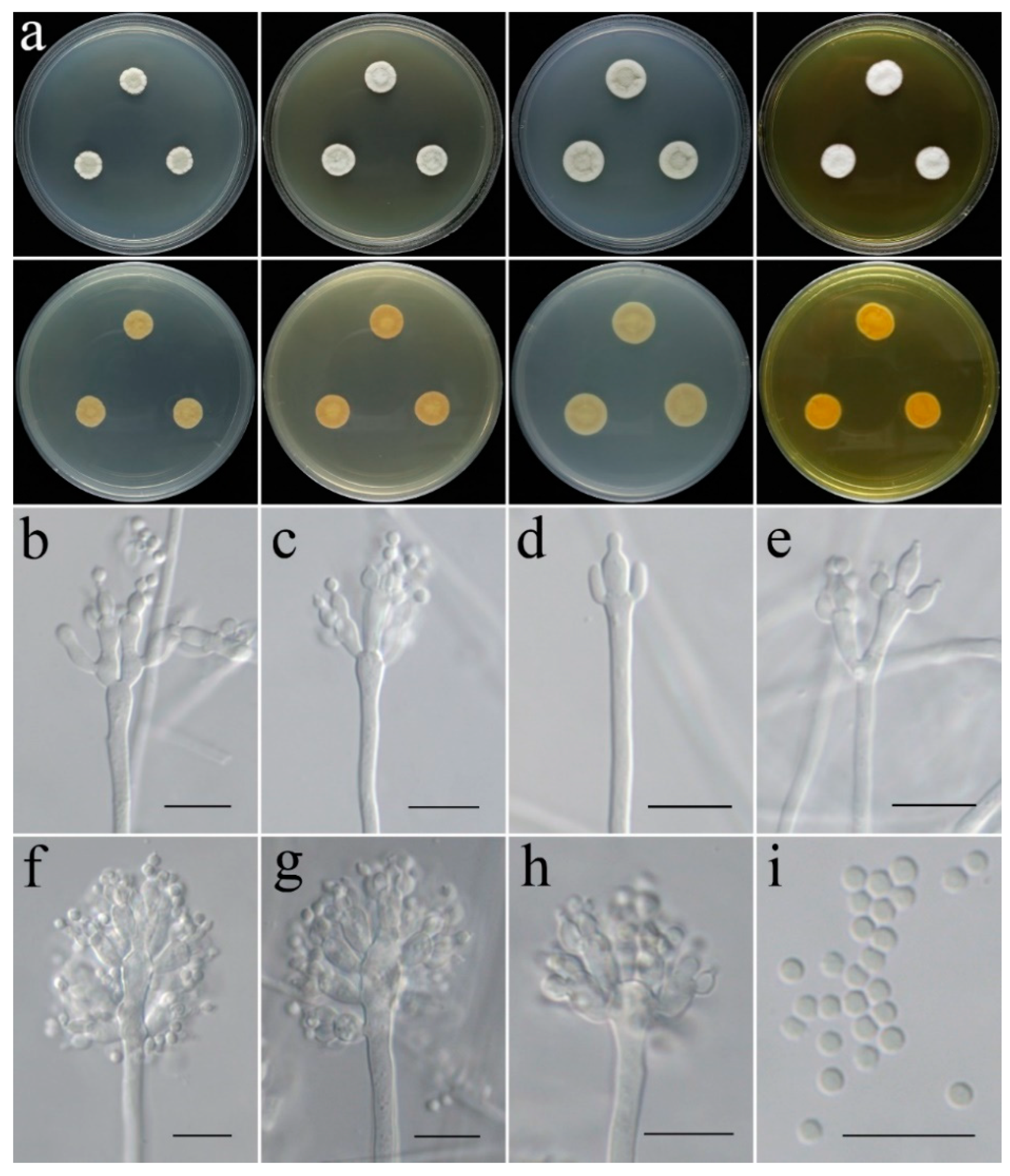
- Penicillium danzhouense C. Liu, Z.Q. Zeng & W.Y. Zhuang, sp. nov. Figure 9.
- Penicillium tenue C. Liu, Z.Q. Zeng & W.Y. Zhuang, sp. nov. Figure 10.
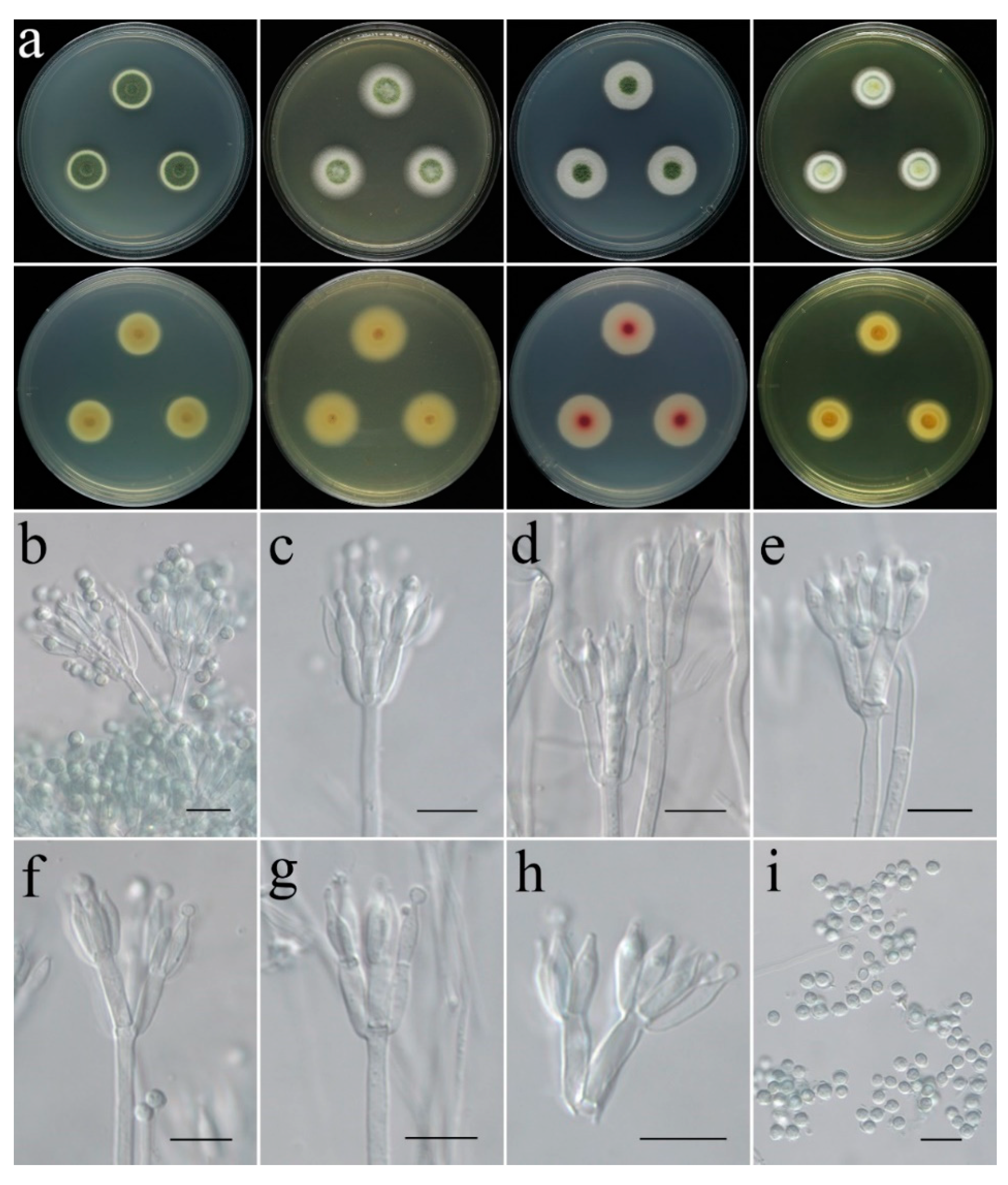
- Penicillium zhanjiangense C. Liu, Z.Q. Zeng & W.Y. Zhuang, sp. nov. Figure 11.
- Talaromyces virens C. Liu, Z.Q. Zeng & W.Y. Zhuang, sp. nov. Figure 12.
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, H.; Jia, Y.; Zhao, D.; Xiu, T.; Duan, F. A tidal flat wetlands delineation and classification method for high-resolution imagery. ISPRS Int. J. Geo-Inf. 2021, 10, 451. [Google Scholar] [CrossRef]
- Li, P.D.; Jeewon, R.; Aruna, B.; Li, H.Y.; Lin, F.C.; Wang, H.K. Metabarcoding reveals differences in fungal communities between unflooded versus tidal flat soil in coastal saline ecosystem. Sci. Total Environ. 2019, 690, 911–922. [Google Scholar] [CrossRef]
- Li, W.; Wang, M.M.; Bian, X.M.; Guo, J.J.; Cai, L. A high-level fungal diversity in the intertidal sediment of Chinese seas presents the spatial variation of community composition. Front. Microbiol. 2016, 7, 2098. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef] [PubMed]
- Rossman, A.Y.; Allen, W.C.; Braun, U.; Castlebury, L.A.; Chaverri, P.; Crous, P.W.; Hawksworth, D.L.; Hyde, K.D.; Johnston, P.; Lombard, L.; et al. Overlooked competing asexual and sexually typified generic names of Ascomycota with recommendations for their use or protection. IMA Fungus 2016, 7, 289–308. [Google Scholar] [CrossRef] [PubMed]
- Han, P.J.; Sun, J.Q.; Wang, L. Two new sexual Talaromyces species discovered in estuary soil in China. J. Fungi 2021, 8, 36. [Google Scholar] [CrossRef]
- Houbraken, J.; Seifert, K.A.; Samson, R.A. Penicillium hermansii, a new species causing smoky mould in white button mushroom production. Mycol. Prog. 2019, 18, 229–236. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Larsen, T.O. Extrolites of Aspergillus fumigatus and other pathogenic species in Aspergillus section Fumigati. Front. Microbiol. 2015, 6, 1485. [Google Scholar] [CrossRef]
- Le, T.; Chi, N.H.; Cuc, N.T.K.; Sieu, T.P.M.; Shikuma, C.M.; Farrar, J.; Day, J.N. AIDS-associated Penicillium marneffei infection of the central nervous system. Clin. Infect. Dis. 2010, 51, 1458–1462. [Google Scholar] [CrossRef]
- Fleming, A. Penicillin. Brit. Med. J. 1941, 2, 386. [Google Scholar] [CrossRef]
- Wang, X.C.; Zhuang, W.Y. New species of Aspergillus (Aspergillaceae) from tropical islands of China. J. Fungi 2022, 8, 225. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Moller, L.L.H.; Larsen, T.O.; Kumar, R.; Arnau, J. Safety of the fungal workhorses of industrial biotechnology: Update on the mycotoxin and secondary metabolite potential of Aspergillus niger, Aspergillus oryzae, and Trichoderma reesei. Appl. Microbiol. Biotechnol. 2018, 102, 9481–9515. [Google Scholar] [CrossRef]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [PubMed]
- Visagie, C.M.; Hirooka, Y.; Tanney, T.; Whitfield, E.; Mwange, K.; Meijer, M.; Amend, A.S.; Seifert, K.A.; Samson, R.A. Aspergillus, Penicillium and Talaromyces isolated from house dust samples collected around the world. Stud. Mycol. 2014, 78, 63–139. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Visagie, C.M.; Houbraken, J.; Samson, R.A. Polyphasic taxonomy of the genus Talaromyces. Stud. Mycol. 2014, 78, 175–341. [Google Scholar] [CrossRef]
- Silva, J.J.; Fungaro, M.H.P.; Wang, X.; Larsen, T.O.; Frisvad, J.C.; Taniwaki, M.H.; Iamanaka, B.T. Deep genotypic species delimitation of Aspergillus Section Flavi isolated from Brazilian foodstuffs and the description of Aspergillus annui sp. nov. and Aspergillus saccharicola sp. nov. J. Fungi 2022, 8, 1279. [Google Scholar] [CrossRef]
- Sun, B.D.; Luo, C.L.; Bills, G.F.; Li, J.B.; Huang, P.P.; Wang, L.; Jiang, X.Z.; Chen, A.J. Four new species of Aspergillus Subgenus Nidulantes from China. J. Fungi 2022, 8, 1205. [Google Scholar] [CrossRef]
- Alves, V.C.S.; Lira, R.A.; Lima, J.M.S.; Barbosa, R.N.; Bento, D.M.; Barbier, E.; Bernard, E.; Souza-Motta, C.M.; Bezerra, J.D.P. Unravelling the fungal darkness in a tropical cave: Richness and the description of one new genus and six new species. Fungal Syst. Evol. 2022, 10, 139–167. [Google Scholar] [CrossRef]
- Glässnerová, K.; Sklenář, F.; Jurjević, Ž.; Houbraken, J.; Yaguchi, T.; Visagie, C.M.; Gené, J.; Siqueira, J.P.Z.; Kubátová, A.; Kolařík, M.; et al. A monograph of Aspergillus section Candidi. Stud. Mycol. 2022, 102, 1–51. [Google Scholar] [CrossRef]
- Tan, Y.P.; Shivas, R.G. Index of Australian Fungi. Zenodo 2022, 3. [Google Scholar] [CrossRef]
- Cañete-Gibas, C.F.; Patterson, H.P.; Sanders, C.J.; Mele, J.; Fan, H.; David, M.; Wiederhold, N.P. Species distribution and antifungal susceptibilities of Aspergillus section Terrei isolates in clinical samples from the United States and description of Aspergillus pseudoalabamensis sp. nov. Pathogens 2023, 12, 579. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Boers, J.; Holdom, D.; Osieck, E.R.; Steinrucken, T.V.; Tan, Y.P.; Vitelli, J.S.; Shivas, R.G.; Barrett, M.; Boxshall, A.-G.; et al. Fungal Planet description sheets: 1383–1435. Persoonia 2022, 48, 261–371. [Google Scholar] [CrossRef]
- Visagie, C.M.; Frisvad, J.C.; Houbraken, J.; Visagie, A.; Samson, R.A.; Jacobs, K. A re-evaluation of Penicillium section Canescentia, including the description of five new species. Persoonia 2021, 46, 163–187. [Google Scholar] [CrossRef]
- Labuda, R.; Bacher, M.; Rosenau, T.; Gasparotto, E.; Gratzl, H.; Doppler, M.; Sulyok, M.; Kubátova, A.; Berger, H.; Cank, K.; et al. Polyphasic approach utilized for the identification of two new toxigenic members of Penicillium section Exilicaulis, P. krskae and P. silybi spp. nov. J. Fungi 2021, 7, e557. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.J.; Jeewon, R.; Dhandevi, P.; Durairajan, S.S.K.; Li, H.; Lin, F.C.; Wang, H.K. A novel species of Penicillium with inhibitory effects against Pyricularia oryzae and fungal pathogens inducing citrus diseases. Front. Cell. Infect. Microbiol. 2021, 10, e604504. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Andrade, E.; Stchigel, A.M.; Cano-Lira, J.F. New xerophilic species of Penicillium from soil. J. Fungi 2021, 7, e126. [Google Scholar] [CrossRef] [PubMed]
- Torres-Garcia, D.; Gené, J.; García, D. New and interesting species of Penicillium (Eurotiomycetes, Aspergillaceae) in freshwater sediments from Spain. MycoKeys 2022, 86, 103–145. [Google Scholar] [CrossRef]
- Xu, K.X.; Shan, X.N.; Ruan, Y.M.; Deng, J.X.; Wang, L. Three new Penicillium species isolated from the tidal flats of China. PeerJ 2022, 10, e13224. [Google Scholar] [CrossRef]
- Perini, L.; Gostinčar, C.; Likar, M.; Frisvad, J.C.; Kostanjšek, R.; Nicholes, M.; Williamson, C.; Anesio, A.M.; Zalar, P.; Gunde-Cimerman, N. Interactions of fungi and algae from the Greenland ice sheet. Microb. Ecol. 2023, 86, 282–296. [Google Scholar] [CrossRef]
- Silva, I.R.; Sousa, T.F.; de Queiroz, C.A.; Castro, G.D.; Caniato, F.F.; de Medeiros, L.S.; Angolini, C.F.; Hanada, R.E.; Koolen, H.H.; Silva, G.F. Penicillium amapaense sp. nov., section Exilicaulis, and new records of Penicillium labradorum in Brazil isolated from Amazon River sediments with potential applications in agriculture and biotechnology. Mycol. Prog. 2023, 22, 23. [Google Scholar] [CrossRef]
- Visagie, C.M.; Yilmaz, N. Along the footpath of Penicillium discovery: Six new species from the Woodville Big Tree Forest Trail. Mycologia 2023, 115, 87–106. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.P.; Bishop-Hurley, S.L.; Shivas, R.G.; Cowan, D.A.; Maggs-Kolling, G.; Maharachchikumbura, S.S.N.; Pinruan, U.; Bransgrove, K.L.; De la Pena-Lastra, S.; Larsson, E.; et al. Fungal Planet description sheets: 1436–1477. Persoonia 2022, 49, 261–350. [Google Scholar] [CrossRef]
- Barbosa, R.; Santos, J.E.; Bezerra, J.; Istel, Ł.; Houbraken, J.; Oliveira, N.; Souza-Motta, C. Brazilian Atlantic Forest and Pampa Biomes in the spotlight: An overview of Aspergillus, Penicillium, and Talaromyces (Eurotiales) species and the description of Penicillium nordestinense sp. nov. Acta Bot. Bras. 2022, 36. [Google Scholar] [CrossRef]
- Kirchmair, M.; Embacher, J.; Heimdörfer, D.; Walch, G.; Neuhauser, S. Penicillium poederi and P. tirolense, two new species of section Torulomyces. Fungal Syst. Evol. 2022, 10, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Sobol, M.S.; Hoshino, T.; Delgado, V.; Futagami, T.; Kadooka, C.; Inagaki, F.; Kiel Reese, B. Genome characterization of two novel deep-sea sediment fungi, Penicillium pacificagyrus sp. nov. and Penicillium pacificasedimenti sp. nov., from South Pacific Gyre subseafloor sediments, highlights survivability. BMC Genom. 2023, 24, 249. [Google Scholar] [CrossRef] [PubMed]
- Ashtekar, N.; Rajeshkumar, K.C.; Yilmaz, N.; Visagie, C.M. A new Penicillium section Citrina species and series from India. Mycol. Prog. 2022, 21, 42. [Google Scholar] [CrossRef]
- Lenz, A.R.; Balbinot, E.; de Abreu, F.P.; de Oliveira, N.S.; Fontana, R.C.; de Avila, E.S.S.; Park, M.S.; Lim, Y.W.; Houbraken, J.; Camassola, M.; et al. Taxonomy, comparative genomics and evolutionary insights of Penicillium ucsense: A novel species in series Oxalica. Antonie Van Leeuwenhoek 2022, 115, 1009–1029. [Google Scholar] [CrossRef]
- Nguyen, V.D.; Pham, T.T. Penicillium vietnamense sp. nov., the first novel marine fungi species described from Vietnam with a unique conidiophore structure and molecular phylogeny of Penicillium section Charlesia. Mycobiology 2022, 50, 155–165. [Google Scholar] [CrossRef]
- Sun, X.R.; Xu, M.Y.; Kong, W.L.; Wu, F.; Zhang, Y.; Xie, X.L.; Li, D.W.; Wu, X.Q. Fine identification and classification of a novel beneficial Talaromyces fungal species from Masson pine rhizosphere soil. J. Fungi 2022, 8, 155. [Google Scholar] [CrossRef]
- Wang, X.C.; Zhuang, W.Y. New Species of Talaromyces (Trichocomaceae, Eurotiales) from Southwestern China. J. Fungi 2022, 8, 647. [Google Scholar] [CrossRef]
- Nuankaew, S.; Chuaseeharonnachai, C.; Preedanon, S.; Somrithipol, S.; Saengkaewsuk, S.; Kwantong, P.; Phookongchai, S.; Srikitikulchai, P.; Kobmoo, N.; Wang, X.C.; et al. Two novel species of Talaromyces discovered in a karst cave in the Satun UNESCO global geopark of southern Thailand. J. Fungi 2022, 8, 825. [Google Scholar] [CrossRef]
- Guerra Sierra, B.E.; Arteaga-Figueroa, L.A.; Sierra-Pelaéz, S.; Alvarez, J.C. Talaromyces santanderensis: A new cadmium-tolerant fungus from Cacao soils in Colombia. J. Fungi 2022, 8, 1042. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.Z.; Xu, X.L.; Wang, L. Four new species of Talaromyces section Talaromyces discovered in China. Mycologia 2021, 113, 492–508. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.W.; Jurjević, Ž. New species of Talaromyces isolated from maize, indoor air, and other substrates. Mycologia 2017, 109, 537–556. [Google Scholar]
- Soares, C.; Rodrigues, P.; Peterson, S.W.; Lima, N.; Venâncio, A. Three new species of Aspergillus section Flavi isolated from almonds and maize in Portugal. Mycologia 2012, 104, 682–697. [Google Scholar] [CrossRef]
- Ramírez, C.; Martínez, A.T. Four new species of Penicillium isolated from different substrata. Mycopathologia 1981, 74, 163–171. [Google Scholar] [CrossRef]
- Murray, N.J.; Phinn, S.R.; DeWitt, M.; Ferrari, R.; Johnston, R.; Lyons, M.B.; Clinton, N.; Thau, D.; Fuller, R.A. The global distribution and trajectory of tidal flats. Nature 2019, 565, 222. [Google Scholar] [CrossRef]
- Pitt, J.I. The Genus Penicillium and Its Teleomorphic States Eupenicillium and Talaromyces; Academic Press: London, UK, 1980; pp. 1–634. [Google Scholar]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi; CBS-KNAW Fungal Biodiversity Center: Utrecht, The Netherlands, 2010; pp. 1–390. [Google Scholar]
- Frisvad, J.C. Physiological criteria and mycotoxin production as aids in identification of common asymmetric penicillia. Appl. Environ. Microb. 1981, 41, 568–579. [Google Scholar] [CrossRef]
- Wang, X.C.; Chen, K.; Qin, W.T.; Zhuang, W.Y. Talaromyces heiheensis and T. mangshanicus, two new species from China. Mycol. Prog. 2017, 16, 73–81. [Google Scholar] [CrossRef]
- Zhang, Z.K.; Wang, X.C.; Zhuang, W.Y.; Cheng, X.H.; Zhao, P. New species of Talaromyces (Fungi) isolated from soil in southwestern China. Biology 2021, 10, 745. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfland, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microb. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.B.; Cho, H.S.; Shin, H.D.; Frisvad, J.C.; Samson, R.A. Novel Neosartorya species isolated from soil in Korea. Int. J. Syst. Evol. Microbiol. 2006, 56, 477–486. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and other Methods), Version 4b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest v2; Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Page, R.D. TreeView: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar]
- Cunningham, C.W. Can three incongruence tests predict when data should be combined? Mol. Biol. Evol. 1997, 14, 733–740. [Google Scholar] [CrossRef]
- Hubka, V.; Novakova, A.; Kolarík, M.; Jurjević, Ž.; Peterson, S.W. Revision of Aspergillus section Flavipedes: Seven new species and proposal of section Jani sect. nov. Mycologia 2015, 107, 169–208. [Google Scholar] [CrossRef]
- Stolk, A.C.; Malla, D.S. Penicillium inflatum sp. nov. Persoonia 1971, 6, 197–200. [Google Scholar]
- De Scott, B. Studies on the genus Eupenicillium Ludwig IV. New species from soil. Mycopath. Mycol. Appl. 1968, 36, 1–27. [Google Scholar] [CrossRef]
- Stolk, A.C. Four new species of Penicillium. Antonie Van Leeuwenhoek 1969, 35, 261–274. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Seifert, K.A.; Samson, R.A.; Jacobs, K. Four new Penicillium species isolated from the fynbos biome in South Africa, including a multigene phylogeny of section Lanata-Divaricata. Mycol. Prog. 2015, 14, 1–23. [Google Scholar] [CrossRef]
- Wang, X.C.; Chen, K.; Xia, Y.W.; Wang, L.; Li, T.H.; Zhuang, W.Y. A new species of Talaromyces (Trichocomaceae) from the Xisha Islands, Hainan, China. Phytotaxa 2016, 267, 187–200. [Google Scholar] [CrossRef]
- Kagiyama, I.; Kato, H.; Nehira, T.; Frisvad, J.C.; Sherman, D.H.; Williams, R.M.; Tsukamoto, S. Taichunamides: Prenylated indole alkaloids from Aspergillus taichungensis (IBT 19404). Angew. Chem. Int. Ed. 2016, 55, 1128–1132. [Google Scholar] [CrossRef]
- Hubka, V.; Novakova, A.; Jurjevic, Z.; Sklenář, F.; Frisvad, J.C.; Houbraken, J.; Arendrup, M.C.; Jørgensen, K.M.; Siqueira, J.P.Z.; Gené, J.; et al. Polyphasic data support the plitting of Aspergillus candidus into two species; proposal of Aspergillus dobrogensis sp. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 995–1011. [Google Scholar] [CrossRef]
- Kato, H.; Nakahara, T.; Sugimoto, K.; Matsuo, K.; Kagiyama, I.; Frisvad, J.C.; Sherman, D.H.; Williams, R.M.; Tsukamoto, S. Correction to “Isolation of notoamide S and enantiomeric 6-epi-stephacidin A from the terrestrial fungus Aspergillus amoenus: Biogenetic implications”. Org. Lett. 2018, 20, 315. [Google Scholar] [CrossRef]
- Peterson, S.W.; Corneli, S.; Hjelle, J.T.; Miller-Hjelle, M.A.; Nowak, D.M.; Bonneau, P.A. Penicillium pimiteouiense: A new species isolated from polycystic kidney cell cultures. Mycologia 1999, 91, 269–277. [Google Scholar] [CrossRef]
- Visagie, C.M.; Seifert, K.A.; Houbraken, J.; Samson, R.A.; Jacobs, K. A phylogenetic revision of Penicillium sect. Exilicaulis, including nine new species from fynbos in South Africa. IMA Fungus 2016, 7, 75–117. [Google Scholar]
- Janso, J.E.; Bernan, V.S.; Greenstein, M.; Bugni, T.S.; Ireland, C.M. Penicillium dravuni, a new marine-derived species from an alga in Fiji. Mycologia 2005, 97, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.S.; Pan, J.H.; Tang, W.C.; Chen, Q.J.; Lin, Y.C. Biodiversity and biotechnological potential of mangrove-associated fungi. J. Forestry Res. 2009, 20, 63–72. [Google Scholar] [CrossRef]
- Jang, H.J.; Song, J.; Joung, Y.; Cho, J.C. Pelagibacterium sediminicola sp. nov., isolated from tidal flat sediment. Int. J. Syst. Evol. Microbiol. 2019, 69, 2651–2657. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Wang, Y.; Liu, P.; Dong, T.H.; Zhu, W.M. Thiodiketopiperazines from the marine-derived fungus Phoma sp. OUCMDZ-1847. J. Nat. Prod. 2014, 77, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Ameen, F.; Al-Homaidan, A.A. Compost inoculated with fungi from a mangrove habitat improved the growth and disease defense of vegetable plants. Sustainability 2021, 13, 124. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain Numbers | ITS | BenA | CaM | RPB2 |
|---|---|---|---|---|---|
| sect. Candidi | |||||
| A. campestris | CBS 348.81 T | EF669577 | EU014091 | EF669535 | EF669619 |
| A. candidus | CBS 566.65 T | EF669592 | EU014089 | EF669550 | EF669634 |
| A. dobrogensis | CBS 143370 T | LT626959 | LT627027 | LT558722 | LT627028 |
| A. liaoningensis | CGMCC 3.25201 T | ON563148 | ON231293 | ON470836 | ON470844 |
| A. magnus | UAMH 1324 T | ON156376 | ON164570 | ON164619 | ON164517 |
| A. neotritici | CCF 3853 T | FR727136 | FR775327 | HE661598 | LT627021 |
| A. pragensis | CBS 135591 T | FR727138 | HE661604 | FR751452 | LN849445 |
| A. subalbidus | CBS 567.65 T | EF669593 | KP987050 | EF669551 | EF669635 |
| A. taichungensis | CCF 5597 T | LT626957 | EU076297 | HG916679 | LT627016 |
| A. tenebricus | CBS 147048 T | ON156389 | ON164584 | ON164623 | ON164532 |
| sect. Flavipedes | |||||
| A. ardalensis | CBS 134372 T | FR733808 | HG916683 | HG916725 | HG916704 |
| A. capensis | CBS 138188 T | KJ775550 | KJ775072 | KJ775279 | KP987020 |
| A. flavipes | ATCC 24487 T | EF669591 | EU014085 | EF669549 | EF669633 |
| A. iizukae | CBS 541.69 T | EF669597 | EU014086 | EF669555 | EF669639 |
| A. luppii | CBS 653.74 T | EF669617 | EU014079 | EF669575 | EF669659 |
| A. micronesiensis | CBS 138183 T | KJ775548 | KJ775085 | KP987067 | KP987023 |
| A. movilensis | CBS 134395 T | KP987089 | HG916697 | HG916740 | HG916718 |
| A. neoflavipes | CBS 260.73 T | EF669614 | EU014084 | EF669572 | EF669656 |
| A. neoniveus | CBS 261.73 T | EF669612 | EU014098 | EF669570 | KP987024 |
| A. olivimuriae | NRRL 66783 | MH298877 | MH492010 | MH492011 | MH492012 |
| A. plumeriae | CGMCC 3.25202 T | ON563147 | ON231292 | ON470835 | ON470843 |
| A. polyporicola | NRRL 32683 T | EF669595 | EU014088 | EF669553 | EF669637 |
| A. spelaeus | CBS 134371 T | HG915905 | HG916698 | HG916741 | HG916719 |
| A. suttoniae | UTHSCSA DI14-215 | LT899487 | LT899536 | LT899589 | LT899644 |
| A. templicola | CBS 138181 T | KJ775545 | KJ775092 | KJ775394 | KP987017 |
| A. urmiensis | CBS 139558 T | KP987073 | KP987041 | KP987056 | KP987030 |
| sect. Cremei | |||||
| A. arxii | CBS 525.83 T | MN431361 | MN969365 | MN969223 | JN121529 |
| A. brunneouniseriatus | NRRL 4273 T | EF652141 | EF652123 | EF652138 | EF652089 |
| A. chaetosartoryae | NRRL 5501 T | EF652144 | EF652117 | EF652129 | EF652099 |
| A. chrysellus | NRRL 5084 T | EF652155 | EF652109 | EF652136 | EF652090 |
| A. citocrescens | CBS 140566 T | FR727121 | FR775317 | LN878969 | MN969163 |
| A. cremeus | NRRL 5081 T | EF652149 | EF652120 | EF652125 | EF652101 |
| A. dimorphicus | NRRL 3650 T | EF652154 | EF652111 | EF652135 | EF652096 |
| A. europaeus | CBS 134393T | LN908996 | LN909006 | LN909007 | LT548274 |
| A. flaschentraegeri | NRRL 5042 T | EF652150 | EF652113 | EF652130 | EF652102 |
| A. gorakhpurensis | NRRL 3649 T | EF652145 | EF652114 | EF652126 | EF652097 |
| A. inflatus | CBS 682.70 T | FJ531054 | FJ531008 | FJ531090 | JN406529 |
| A. itaconicus | NRRL 161 T | EF652147 | EF652118 | EF652140 | EF652103 |
| A. koreanus | NIBR EML-GSNP1-1 | KX216525 | KX216530 | KX216528 | KX216531 |
| A. lebretii | URM 8451 T | ON862928 | OP672381 | OP290539 | OP290510 |
| A. pulvinus | NRRL 5078 T | EF652159 | EF652121 | EF652139 | EF652104 |
| A. stromatoides | CBS 500.65 T | EF652146 | FJ531038 | EF652127 | EF652098 |
| A. subinflatus | CGMCC 3.25203 T | ON563146 | ON231291 | ON470834 | ON470845 |
| A. tardus | CBS 433.93 T | FJ531045 | FJ531001 | FJ531084 | n.a. |
| A. wentii | NRRL 375 T | EF652151 | EF652106 | EF652131 | EF652092 |
| Sect. Lanata-divaricata | |||||
| P. abidjanum | CBS 246.67 T | GU981582 | GU981650 | KF296383 | JN121469 |
| P. alagoense | URM 8086 T | MK804503 | MK802333 | MK802336 | MK802338 |
| P. amphipolaria | DAOMC 250551 T | KT887872 | KT887833 | KT887794 | n.a. |
| P. annulatum | CBS 135126 T | JX091426 | JX091514 | JX141545 | KF296410 |
| P. araracuaraense | CBS 113149 T | GU981597 | GU981642 | KF296373 | KF296414 |
| P. ausonanum | FMR 16948 T | LR655808 | LR655809 | LR655810 | LR655811 |
| P. austrosinense | CGMCC 3.18797 T | KY495007 | KY495116 | KY494947 | KY495061 |
| P. bissettii | DAOMC 167011 T | KT887845 | KT887806 | KT887767 | MN969107 |
| P. brasilianum | CBS 253.55 T | GU981577 | GU981629 | MN969239 | KF296420 |
| P. brefeldianum | CBS 235.81 T | AF033435 | GU981623 | EU021683 | KF296421 |
| P. camponotum | KAS 2177 T | KT887855 | KT887816 | KT887777 | MN969179 |
| P. caperatum | CBS 443.75 T | KC411761 | GU981660 | KF296392 | KF296422 |
| P. cataractum | DAOMC 250534 T | KT887847 | KT887808 | KT887769 | n.a. |
| P. cluniae | CBS 326.89 T | KF296406 | KF296471 | KF296402 | KF296424 |
| P. coeruleum | CBS 141.45 T | GU981606 | GU981655 | KF296393 | KF296425 |
| P. cremeogriseum | CBS 223.66 T | GU981586 | GU981624 | KF296403 | KF296426 |
| P. curticaule | CBS 135127 T | FJ231021 | JX091526 | JX141536 | KF296417 |
| P. daleae | CBS 211.28 T | GU981583 | GU981649 | KF296385 | KF296427 |
| P. diatomitis | CCF 3904 T | FJ430748 | HE651133 | LT970912 | LT797560 |
| P. donggangicum | AS3.15900 T | MW946996 | MZ004914 | MZ004918 | MW979253 |
| P. echinulonalgiovense | CBS 328.59 T | GU981587 | GU981631 | KX961269 | KX961301 |
| P. ehrlichii | CBS 324.48 T | AF033432 | GU981652 | KF296395 | KF296428 |
| P. elleniae | CBS 118135 T | GU981612 | GU981663 | KF296389 | KF296429 |
| P. excelsum | ITAL7572 T | KR815341 | KP691061 | KR815342 | MN969166 |
| P. flaviroseum | CGMCC 3.18805 T | KY495032 | KY495141 | KY494972 | KY495083 |
| P. fructuariae-cellae | CBS 145110 T | MK039434 | KU554679 | MK045337 | n.a. |
| P. glaucoroseum | CBS 138908 T | MN431390 | MN969383 | MN969257 | MN969119 |
| P. globosum | CGMCC 3.18800 T | KY495014 | KY495123 | KY494954 | KY495067 |
| P. griseoflavum | CGMCC 3.18799 T | KY495011 | KY495120 | KY494951 | KY495064 |
| P. griseopurpureum | CBS 406.65 T | KF296408 | KF296467 | KF296384 | KF296431 |
| P. guaibinense | CCDCA 11512 T | MH674389 | MH674391 | MH674393 | n.a. |
| P. guangxiense | CGMCC 3.18793 T | KY494986 | KY495095 | KY494926 | n.a. |
| P. hainanense | CGMCC 3.18798 T | KY495009 | KY495118 | KY494949 | n.a. |
| P. hepuense | AS3.16039 T | MW946994 | MZ004912 | MZ004916 | MW979254 |
| P. infrabuccalum | DAOMC 250537 T | KT887856 | KT887817 | KT887778 | n.a. |
| P. janthinellum | CBS 340.48 T | GU981585 | GU981625 | KF296401 | JN121497 |
| P. javanicum | CBS 341.48 T | GU981613 | GU981657 | KF296387 | JN121498 |
| P. jianfenglingense | CGMCC 3.18802 T | KY495016 | KY495125 | KY494956 | KY495069 |
| P. jiaozhouwanicum | AS3.16038 T | MW946993 | MZ004911 | MZ004915 | MW979252 |
| P. koreense | KACC 47721 T | KJ801939 | KM000846 | MN969317 | MN969159 |
| P. laevigatum | CGMCC 3.18801 T | KY495015 | KY495124 | KY494955 | KY495068 |
| P. levitum | CBS 345.48 T | GU981607 | GU981654 | KF296394 | KF296432 |
| P. limosum | CBS 339.97 T | GU981568 | GU981621 | KF296398 | KF296433 |
| P. lineolatum | CBS 188.77 T | GU981579 | GU981620 | KF296397 | KF296434 |
| P. ludwigii | CBS 417.68 T | KF296409 | KF296468 | KF296404 | KF296435 |
| P. malacosphaerulum | CBS 135120 T | FJ231026 | JX091524 | JX141542 | KF296438 |
| P. mariae-crucis | CBS 271.83 T | GU981593 | GU981630 | KF296374 | KF296439 |
| P. meloforme | CBS 445.74 T | KC411762 | GU981656 | KF296396 | KF296440 |
| P. newtonturnerae | BRIP74909a T | OP903478 | OP921964 | OP921962 | OP921963 |
| P. nordestinense | URM 8423 T | OV265270 | OV265324 | OV265272 | OM927721 |
| P. ochrochloron | CBS 357.48 T | GU981604 | GU981672 | KF296378 | KF296445 |
| P. onobense | CBS 174.81 T | GU981575 | GU981627 | KF296371 | KF296447 |
| P. ortum | CBS 135669 T | JX091427 | JX091520 | JX141551 | KF296443 |
| P. oxalicum | CBS 219.30 T | AF033438 | KF296462 | KF296367 | JN121456 |
| P. panissanguineum | DAOMC 250562 T | KT887862 | KT887823 | KT887784 | n.a. |
| P. paraherquei | CBS 338.59 T | AF178511 | KF296465 | KF296372 | KF296449 |
| P. pedernalense | CBS 140770 T | KU255398 | KU255396 | MN969322 | MN969184 |
| P. penarojense | CBS 113178 T | GU981570 | GU981646 | KF296381 | KF296450 |
| P. piscarium | CBS 362.48 T | GU981600 | GU981668 | KF296379 | KF296451 |
| P. pulvillorum | CBS 280.39 T | AF178517 | GU981670 | KF296377 | KF296452 |
| P. raperi | CBS 281.58 T | AF033433 | GU981622 | KF296399 | KF296453 |
| P. reticulisporum | CBS 122.68 T | AF033437 | GU981665 | KF296391 | KF296454 |
| P. rolfsii | CBS 368.48 T | JN617705 | GU981667 | KF296375 | KF296455 |
| P. rubriannulatum | CGMCC 3.18804 T | KY495029 | KY495138 | KY494969 | KY495080 |
| P. setosum | CBS 144865 T | KT852579 | MF184995 | MH105905 | MH016196 |
| P. simplicissimum | CBS 372.48 T | GU981588 | GU981632 | KF296368 | JN121507 |
| P. singorense | CBS 138214 T | KJ775674 | KJ775167 | KJ775403 | n.a. |
| P. skrjabinii | CBS 439.75 T | GU981576 | GU981626 | KF296370 | EU427252 |
| P. soliforme | CGMCC 3.18806 T | KY495038 | KY495147 | KY494978 | n.a. |
| P. soosanum | CCF 3778 T | FJ430745 | FM865811 | LT970913 | LT797561 |
| P. spinuliferum | CGMCC 3.18807 T | KY495040 | KY495149 | KY494980 | KY495090 |
| P. subfuscum | CMW56196 T | MT949907 | MT957412 | MT957454 | MT957480 |
| P. subrubescens | CBS 132785 T | KC346350 | KC346327 | KC346330 | KC346306 |
| P. svalbardense | CBS 122416 T | GU981603 | KC346325 | KC346338 | KF296457 |
| P. tanzanicum | CBS 140968 T | KT887841 | KT887802 | KT887763 | MN969183 |
| P. terrarumae | CBS 131811 T | MN431397 | KX650295 | MN969323 | MN969185 |
| P. ucsense | 2HH T | OM914583 | ON024157 | ON024158 | ON024159 |
| P. uruguayense | CBS 143247 T | LT904729 | LT904699 | LT904698 | MN969200 |
| P. vanderhammenii | CBS 126216 T | GU981574 | GU981647 | KF296382 | KF296458 |
| P. vasconiae | CBS 339.79 T | GU981599 | GU981653 | KF296386 | KF296459 |
| P. vickeryae | BRIP72552a T | OP903479 | OP921966 | n.a. | OP921965 |
| P. viridissimum | CGMCC 3.18796 T | KY495004 | KY495113 | KY494944 | KY495059 |
| P. wotroi | CBS 118171 T | GU981591 | GU981637 | KF296369 | KF296460 |
| P. yunnanense | CGMCC 3.18794 T | KY494990 | KY495099 | KY494930 | KY495048 |
| P. zhanjiangense | CGMCC 3.25206 T | ON563149 | ON231294 | ON470837 | n.a. |
| P. zonatum | CBS 992.72 T | GU981581 | GU981651 | KF296380 | KF296461 |
| sect. Exilicaulis | |||||
| P. allaniae | BRIP 74886a T | OP903475 | OP921956 | OP921954 | OP921955 |
| P. alutaceum | CBS 317.67 T | AF033454 | KJ834430 | KP016768 | JN121489 |
| P. amapaense | URM 8639 T | OL764382 | OL782590 | OL782584 | ON854925 |
| P. aotearoae | PDD 107543 T | KT887874 | KT887835 | KT887796 | MN969174 |
| P. arabicum | CBS 414.69 T | KC411758 | KP016750 | KP016770 | KP064574 |
| P. archerae | BRIP 72549c T | OP903477 | OP921961 | n.a. | OP921960 |
| P. atrolazulinum | CBS 139136 T | JX140913 | JX141077 | JX157416 | KP064575 |
| P. atrosanguineum | CBS 380.75 T | JN617706 | KJ834435 | KP016771 | JN406557 |
| P. burgense | CBS 325.89 T | KC411736 | KJ834437 | KP016772 | JN406572 |
| P. canis | NRRL 62798 T | KJ511291 | KF900167 | KF900177 | KF900196 |
| P. catenatum | CBS 352.67 T | KC411754 | KJ834438 | KP016774 | JN121504 |
| P. chalabudae | CBS 219.66 T | KP016811 | KP016748 | KP016767 | KP064572 |
| P. cinerascens | NRRL 748 T | AF033455 | JX141041 | JX157405 | MN969112 |
| P. cinereoatrum | CBS 222.66 T | KC411700 | KJ834442 | KP125335 | JN406608 |
| P. citreonigrum | CBS 258.29 T | AF033456 | EF198621 | EF198628 | JN121474 |
| P. citreosulfuratum | IMI 92228 T | KP016814 | KP016753 | KP016777 | KP064615 |
| P. consobrinum | CBS 139144 T | JX140888 | JX141135 | JX157453 | KP064619 |
| P. corylophilum | CBS 312.48 T | AF033450 | JX141042 | KP016780 | KP064631 |
| P. cravenianum | CBS 139138 T | JX140900 | JX141076 | JX157418 | KP064636 |
| P. danzhouense | CGMCC 3.25204 T | ON563150 | ON231295 | ON470838 | n.a. |
| P. decumbens | CBS 230.81 T | AY157490 | KJ834446 | KP016782 | JN406601 |
| P. diabolicalicense | CBS 140967 T | KT887840 | KT887801 | KT887762 | MN969175 |
| P. dimorphosporum | CBS 456.70 T | AF081804 | KJ834448 | KP016783 | JN121517 |
| P. dravuni | F01V25 T | AY494856 | n.a. | n.a. | n.a. |
| P. erubescens | CBS 318.67 T | AF033464 | HQ646566 | EU427281 | JN121490 |
| P. fagi | CBS 689.77 T | AF481124 | KJ834449 | KP016784 | JN406540 |
| P. fundyense | CBS 140980 T | KT887853 | KT887814 | KT887775 | MN969176 |
| P. guttulosum | NRRL 907 T | HQ646592 | HQ646576 | HQ646587 | MG386247 |
| P. hemitrachum | CBS 139134 T | FJ231003 | JX141048 | JX157526 | KP064642 |
| P. hermansii | CBS 124296 T | MG333472 | MG386214 | MG386229 | MG386242 |
| P. heteromorphum | CBS 226.89 T | KC411702 | KJ834455 | KP016786 | JN406605 |
| P. katangense | CBS 247.67 T | AF033458 | KP016757 | KP016788 | KP064646 |
| P. kurssanovii | CBS 625.67 T | EF422849 | KP016758 | KP016789 | KP064647 |
| P. labradorum | CBS 145775 T | MK881918 | MK887898 | MK887899 | MK887900 |
| P. laeve | CBS 136665 T | KF667369 | KF667365 | KF667367 | KF667371 |
| P. lapidosum | CBS 343.48 T | MN431392 | KJ834465 | FJ530984 | JN121500 |
| P. maclennaniae | CBS 198.81 T | KC411689 | KJ834468 | KP016791 | KP064648 |
| P. melinii | CBS 218.30 T | AF033449 | KJ834471 | KP016792 | JN406613 |
| P. menonorum | NRRL 50410 T | HQ646591 | HQ646573 | HQ646584 | KF900194 |
| P. meridianum | CBS 314.67 T | AF033451 | KJ834472 | KP016794 | JN406576 |
| P. momoii | CBS 139157 T | JX140895 | JX141073 | JX157479 | KP064673 |
| P. namyslowskii | CBS 353.48 T | AF033463 | JX141067 | KP016795 | JF417430 |
| P. nepalense | CBS 203.84 T | KC411692 | KJ834474 | KP016796 | JN121453 |
| P. ovatum | CBS 136664 T | KF667370 | KF667366 | KF667368 | KF667372 |
| P. pagulum | CBS 139166 T | JX140898 | JX141070 | JX157519 | KP064655 |
| P. parvofructum | CBS 141690 T | LT559091 | LT627645 | LT627646 | MN969197 |
| P. parvum | CBS 359.48 T | AF033460 | HQ646568 | KF900173 | JN406559 |
| P. philippinense | CBS 623.72 T | KC411770 | KJ834482 | KP016799 | JN406543 |
| P. pimiteouiense | CBS 102479 T | AF037431 | HQ646569 | HQ646580 | JN406650 |
| P. punicae | JMRC:SF:12421 T | n.a. | KX839673 | KX839671 | KX839675 |
| P. raciborskii | CBS 224.28 T | AF033447 | JX141069 | KP016800 | JN406607 |
| P. repensicola | CBS 139160 T | JX140893 | JX141150 | JX157490 | KP064660 |
| P. restrictum | CBS 367.48 T | AF033457 | KJ834486 | KP016803 | JN121506 |
| P. rubefaciens | CBS 145.83 T | KC411677 | KJ834487 | KP016804 | JN406627 |
| P. rubidurum | CBS 609.73 T | AF033462 | HQ646574 | HQ646585 | JN406545 |
| P. smithii | CBS 276.83 T | KC411723 | KJ834492 | KP016806 | JN406589 |
| P. striatisporum | CBS 705.68 T | AF038938 | MN969401 | KP016807 | JN406538 |
| P. subturcoseum | CBS 139132 T | FJ231006 | JX141161 | JX157532 | KP064674 |
| P. tenue | CGMCC 3.25205 T | ON563151 | ON231296 | ON470839 | ON470842 |
| P. terrenum | CBS 313.67 T | AM992111 | KJ834496 | KP016808 | JN406577 |
| P. velutinum | CBS 250.32 T | AF033448 | JX141170 | MT478037 | KP064682 |
| P. vinaceum | CBS 389.48 T | AF033461 | HQ646575 | HQ646586 | JN406555 |
| P. xanthomelinii | CBS 139163 T | JX140921 | JX141120 | JX157495 | KP064683 |
| sect. Talaromyces | |||||
| T. aculeatus | CBS 289.48 T | KF741995 | KF741929 | KF741975 | MH793099 |
| T. adpressus | CGMCC 3.18211 T | KU866657 | KU866844 | KU866741 | KU867001 |
| T. alveolaris | CBS 142379 T | LT558969 | LT559086 | LT795596 | LT795597 |
| T. amazonensis | CBS 140373 T | KX011509 | KX011490 | KX011502 | MN969186 |
| T. amestolkiae | CBS 132696 T | JX315660 | JX315623 | KF741937 | JX315698 |
| T. angelicae | KACC 46611 T | KF183638 | KF183640 | KJ885259 | KX961275 |
| T. annesophieae | CBS 142939 T | MF574592 | MF590098 | MF590104 | MN969199 |
| T. apiculatus | CBS 312.59 T | JN899375 | KF741916 | KF741950 | KM023287 |
| T. argentinensis | NRRL 28750 T | MH793045 | MH792917 | MH792981 | MH793108 |
| T. aspriconidius | CBS 141835 T | MN864274 | MN863343 | MN863320 | MN863332 |
| T. atkinsoniae | BRIP 72528a T | OP059084 | OP087524 | n.a. | OP087523 |
| T. aurantiacus | CBS 314.59 T | JN899380 | KF741917 | KF741951 | KX961285 |
| T. aureolinus | CGMCC 3.15865 T | MK837953 | MK837937 | MK837945 | MK837961 |
| T. australis | CBS 137102 T | KF741991 | KF741922 | KF741971 | KX961284 |
| T. bannicus | CGMCC 3.15862 T | MK837955 | MK837939 | MK837947 | MK837963 |
| T. beijingensis | CGMCC 3.18200 T | KU866649 | KU866837 | KU866733 | KU866993 |
| T. brevis | CBS 141833 T | MN864269 | MN863338 | MN863315 | MN863328 |
| T. calidicanius | CBS 112002 T | JN899319 | HQ156944 | KF741934 | KM023311 |
| T. californicus | NRRL 58168 T | MH793056 | MH792928 | MH792992 | MH793119 |
| T. cnidii | KACC 46617 T | KF183639 | KF183641 | KJ885266 | KM023299 |
| T. coprophilus | CBS 142756 T | LT899794 | LT898319 | LT899776 | LT899812 |
| T. cucurbitiradicus | ACCC 39155 T | KY053254 | KY053228 | KY053246 | n.a. |
| T. derxii | CBS 412.89 T | JN899327 | JX494306 | KF741959 | KM023282 |
| T. dimorphus | CGMCC 3.15692 T | KY007095 | KY007111 | KY007103 | KY112593 |
| T. domesticus | NRRL 58121 T | MH793055 | MH792927 | MH792991 | MH793118 |
| T. duclauxii | CBS 322.48 T | JN899342 | JX091384 | KF741955 | JN121491 |
| T. euchlorocarpius | CBM PF1203 T | AB176617 | KJ865733 | KJ885271 | KM023303 |
| T. flavovirens | CBS 102801 T | JN899392 | JX091376 | KF741933 | KX961283 |
| T. flavus | CBS 310.38 T | JN899360 | JX494302 | KF741949 | JF417426 |
| T. francoae | CBS 113134 T | KX011510 | KX011489 | KX011501 | MN969188 |
| T. funiculosus | CBS 272.86 T | JN899377 | MN969408 | KF741945 | KM023293 |
| T. fuscoviridis | CBS 193.69 T | KF741979 | KF741912 | KF741942 | MN969156 |
| T. fusiformis | CGMCC 3.18210 T | KU866656 | KU866843 | KU866740 | KU867000 |
| T. galapagensis | CBS 751.74 T | JN899358 | JX091388 | KF741966 | KX961280 |
| T. ginkgonis | 10725 T | OL638158 | OL689844 | OL689846 | OL689848 |
| T. haitouensis | HR1-7 | MZ045695 | MZ054634 | MZ054637 | MZ054631 |
| T. indigoticus | CBS 100534 T | JN899331 | JX494308 | KF741931 | KX961278 |
| T. intermedius | CBS 152.65 T | JN899332 | JX091387 | KJ885290 | KX961282 |
| T. kabodanensis | CBS 139564 T | KP851981 | KP851986 | KP851995 | MN969190 |
| T. kendrickii | CBS 136666 T | KF741987 | KF741921 | KF741967 | MN969158 |
| T. lentulus | CGMCC 3.15689 T | KY007088 | KY007104 | KY007096 | KY112586 |
| T. liani | CBS 225.66 T | JN899395 | JX091380 | KJ885257 | KX961277 |
| T. louisianensis | NRRL 35823 T | MH793052 | MH792924 | MH792988 | MH793115 |
| T. macrosporus | CBS 317.63 T | JN899333 | JX091382 | KF741952 | KM023292 |
| T. mae | CGMCC 3.15690 T | KY007090 | KY007106 | KY007098 | KY112588 |
| T. malicola | NRRL 3724 T | MH909513 | MH909406 | MH909459 | MH909567 |
| T. mangshanicus | CGMCC 3.18013 T | KX447531 | KX447530 | KX447528 | KX447527 |
| T. marneffei | CBS 388.87 T | JN899344 | JX091389 | KF741958 | KM023283 |
| T. muroii | CBS 756.96 T | MN431394 | KJ865727 | KJ885274 | KX961276 |
| T. mycothecae | CBS 142494 T | MF278326 | LT855561 | LT855564 | LT855567 |
| T. nanjingensis | CCTCCM 2012167 T | MW130720 | MW147759 | MW147760 | MW147762 |
| T. neofusisporus | CGMCC 3.15415 T | KP765385 | KP765381 | KP765383 | MN969165 |
| T. oumae-annae | CBS 138208 T | KJ775720 | KJ775213 | KJ775425 | KX961281 |
| T. panamensis | CBS 128.89 T | JN899362 | HQ156948 | KF741936 | KM023284 |
| T. penicillioides | CGMCC 3.15822 T | MK837956 | MK837940 | MK837948 | MK837964 |
| T. pinophilus | CBS 631.66 T | JN899382 | JX091381 | KF741964 | KM023291 |
| T. pratensis | NRRL 62170 T | MH793075 | MH792948 | MH793012 | MH793139 |
| T. primulinus | CBS 321.48 T | JN899317 | JX494305 | KF741954 | KM023294 |
| T. pseudofuniculosus | CBS 143041 T | LT899796 | LT898323 | LT899778 | LT899814 |
| T. purgamentorum | CBS 113145 T | KX011504 | KX011487 | KX011500 | MN969189 |
| T. purpureogenus | CBS 286.36 T | JN899372 | JX315639 | KF741947 | JX315709 |
| T. qii | CGMCC 3.15414 T | KP765384 | KP765380 | KP765382 | MN969164 |
| T. rapidus | CBS 142382 T | LT558970 | LT559087 | LT795600 | LT795601 |
| T. rosarhiza | GUCC 190040.1 T | MZ221603 | MZ333143 | MZ333137 | MZ333141 |
| T. ruber | CBS 132704 T | JX315662 | JX315629 | KF741938 | JX315700 |
| T. rubicundus | CBS 342.59 T | JN899384 | JX494309 | KF741956 | KM023296 |
| T. rufus | CBS 141834 T | MN864272 | MN863341 | MN863318 | MN863331 |
| T. santanderensis | HF05 T | OP082331 | OP067657 | OP067656 | OP067655 |
| T. sayulitensis | CBS 138204 T | KJ775713 | KJ775206 | KJ775422 | MN969146 |
| T. shilinensis | XCW_SN259 T | OL638159 | OL689845 | OL689847 | OL689849 |
| T. siamensis | CBS 475.88 T | JN899385 | JX091379 | KF741960 | KM023279 |
| T. soli | NRRL 62165 T | MH793074 | MH792947 | MH793011 | MH793138 |
| T. sparsus | CGMCC 3.16003 T | MT077182 | MT083924 | MT083925 | MT083926 |
| T. stellenboschiensis | CBS 135665 T | JX091471 | JX091605 | JX140683 | MN969157 |
| T. stipitatus | CBS 375.48 T | JN899348 | KM111288 | KF741957 | KM023280 |
| T. stollii | CBS 408.93 T | JX315674 | JX315633 | JX315646 | JX315712 |
| T. striatoconidium | CBS 550.89 T | MN431418 | MN969441 | MN969360 | MT156347 |
| T. thailandensis | CBS 133147 T | JX898041 | JX494294 | KF741940 | KM023307 |
| T. tumuli | NRRL 62151 T | MH793071 | MH792944 | MH793008 | MH793135 |
| T. veerkampii | CBS 500.78 T | KF741984 | KF741918 | KF741961 | KX961279 |
| T. verruculosus | NRRL 1050 T | KF741994 | KF741928 | KF741944 | KM023306 |
| T. versatilis | IMI 134755 T | MN431395 | MN969412 | MN969319 | MN969161 |
| T. virens | CGMCC 3.25207 T | ON563150 | ON231297 | ON470840 | ON470841 |
| T. viridis | CBS 114.72 T | AF285782 | JX494310 | KF741935 | JN121430 |
| T. viridulus | CBS 252.87 T | JN899314 | JX091385 | KF741943 | JF417422 |
| T. wushanicus | CS17.05 T | MZ356356 | MZ361347 | MZ361354 | MZ361361 |
| T. xishaensis | CGMCC 3.17995 T | KU644580 | KU644581 | KU644582 | MZ361364 |
| T. yunnanensis | KUMCC 18-0208 T | MT152339 | MT161683 | MT178251 | n.a. |
| T. zhenhaiensis | ZH3-18 T | MZ045697 | MZ054636 | MZ054639 | MZ054633 |
| Outgroup | |||||
| A. glaucus | CBS 516.65 T | EF652052 | EF651887 | EF651989 | EF651934 |
| Hamigera avellanea | CBS 295.48T | AF454075 | EU021664 | EU021682 | EU021627 |
| P. expansum | CBS 325.48T | AY373912 | AY674400 | DQ911134 | JF417427 |
| P. glabrum | CBS 125543 T | GU981567 | GU981619 | KM089152 | JF417447 |
| T. chongqingensis | CS26-67 T | MZ358001 | MZ361343 | MZ361350 | MZ361357 |
| T. trachyspermus | CBS 373.48 T | JN899354 | KF114803 | KJ885281 | JF417432 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Wang, X.-C.; Yu, Z.-H.; Zhuang, W.-Y.; Zeng, Z.-Q. Seven New Species of Eurotiales (Ascomycota) Isolated from Tidal Flat Sediments in China. J. Fungi 2023, 9, 960. https://doi.org/10.3390/jof9100960
Liu C, Wang X-C, Yu Z-H, Zhuang W-Y, Zeng Z-Q. Seven New Species of Eurotiales (Ascomycota) Isolated from Tidal Flat Sediments in China. Journal of Fungi. 2023; 9(10):960. https://doi.org/10.3390/jof9100960
Chicago/Turabian StyleLiu, Chang, Xin-Cun Wang, Zhi-He Yu, Wen-Ying Zhuang, and Zhao-Qing Zeng. 2023. "Seven New Species of Eurotiales (Ascomycota) Isolated from Tidal Flat Sediments in China" Journal of Fungi 9, no. 10: 960. https://doi.org/10.3390/jof9100960
APA StyleLiu, C., Wang, X.-C., Yu, Z.-H., Zhuang, W.-Y., & Zeng, Z.-Q. (2023). Seven New Species of Eurotiales (Ascomycota) Isolated from Tidal Flat Sediments in China. Journal of Fungi, 9(10), 960. https://doi.org/10.3390/jof9100960