Structural Changes Induced by Pulsed Electric Fields Increase the Concentration of Volatiles Released in Red Onion (Allium cepa L. var. Red Pearl) Bulbs


 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
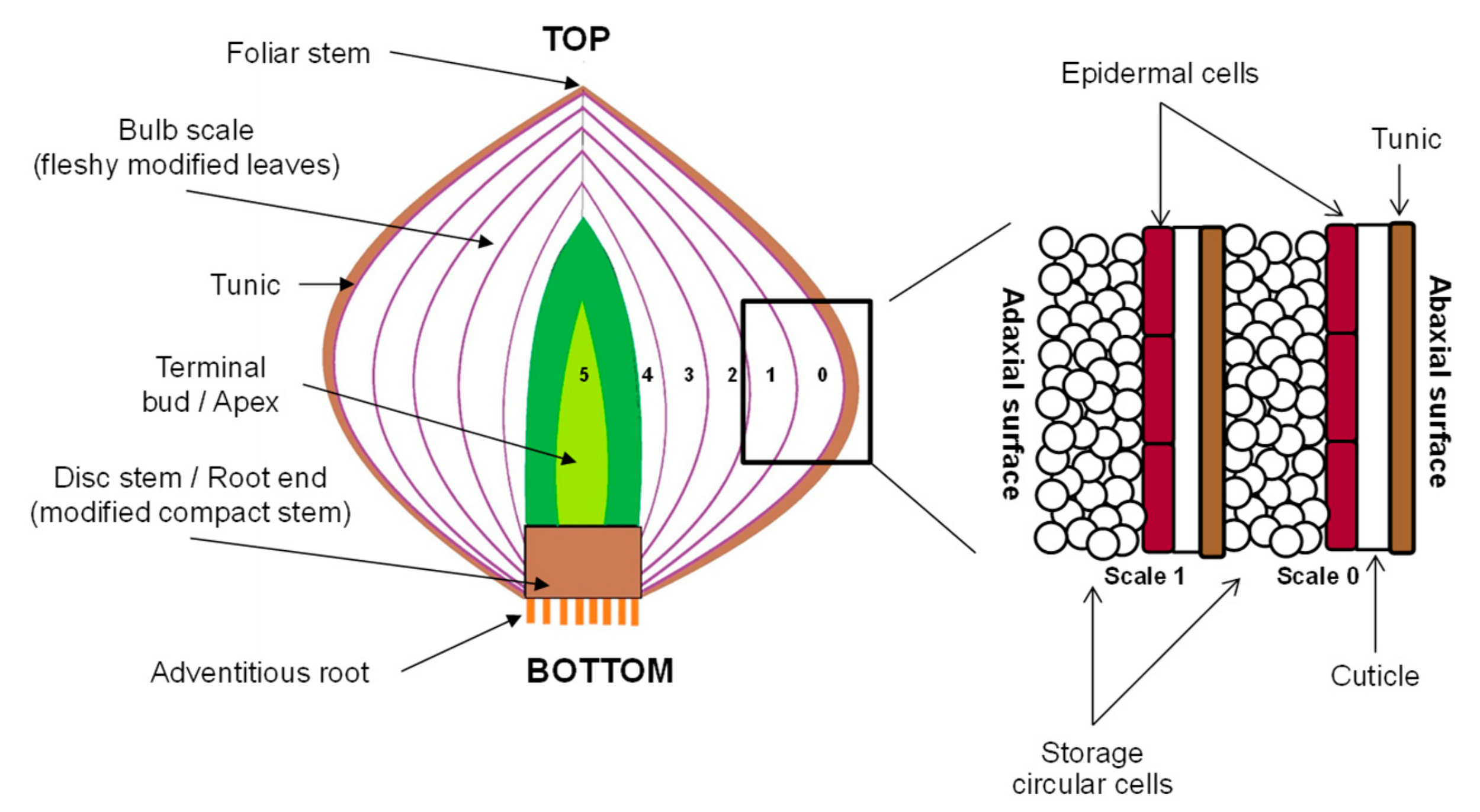
2.1. Onion Bulbs
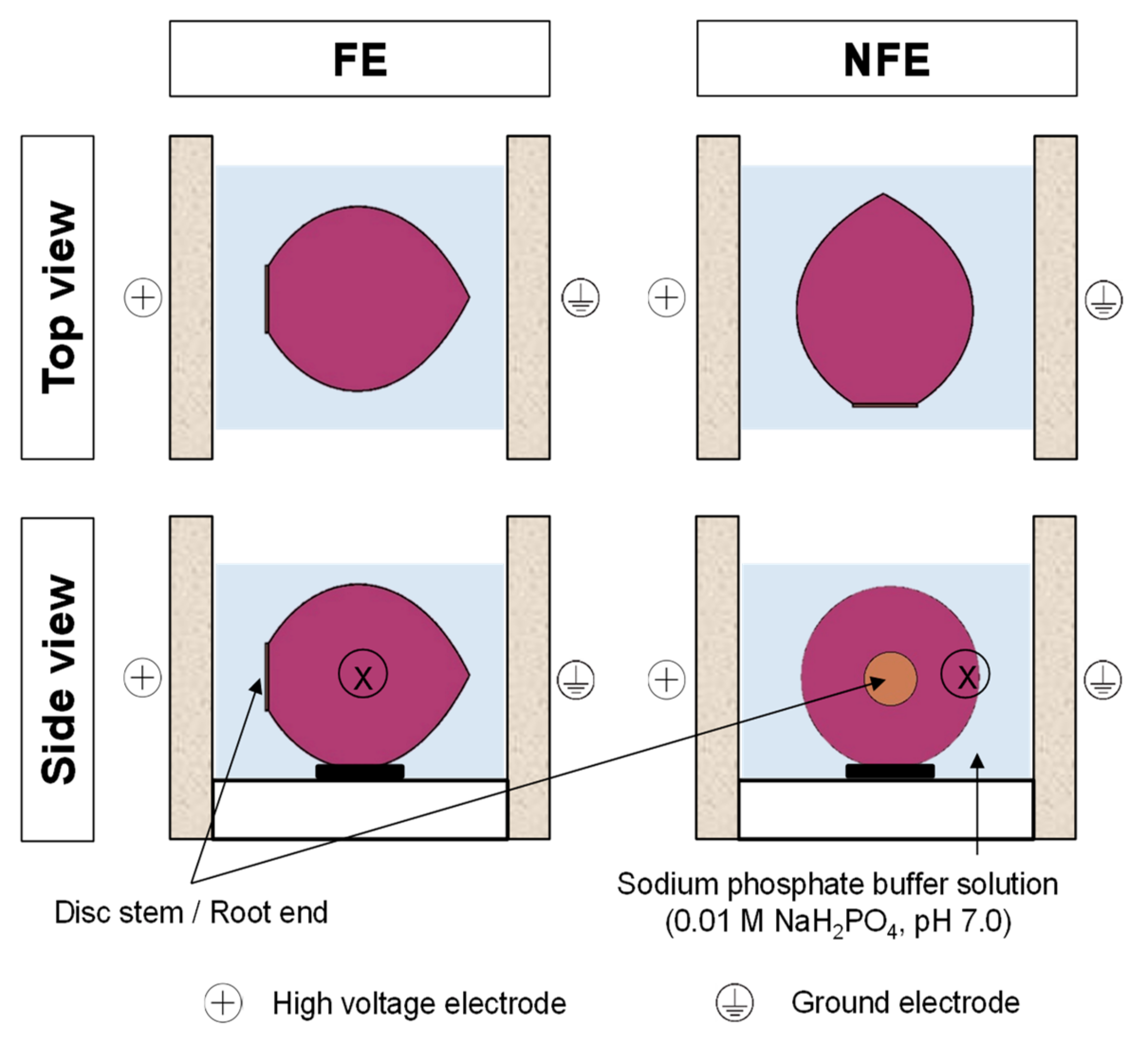
2.2. Pulsed Electric Field Treatments
2.3. Microscopic Evaluation Using Cryo-Scanning Electron Microscopy (Cryo-SEM)
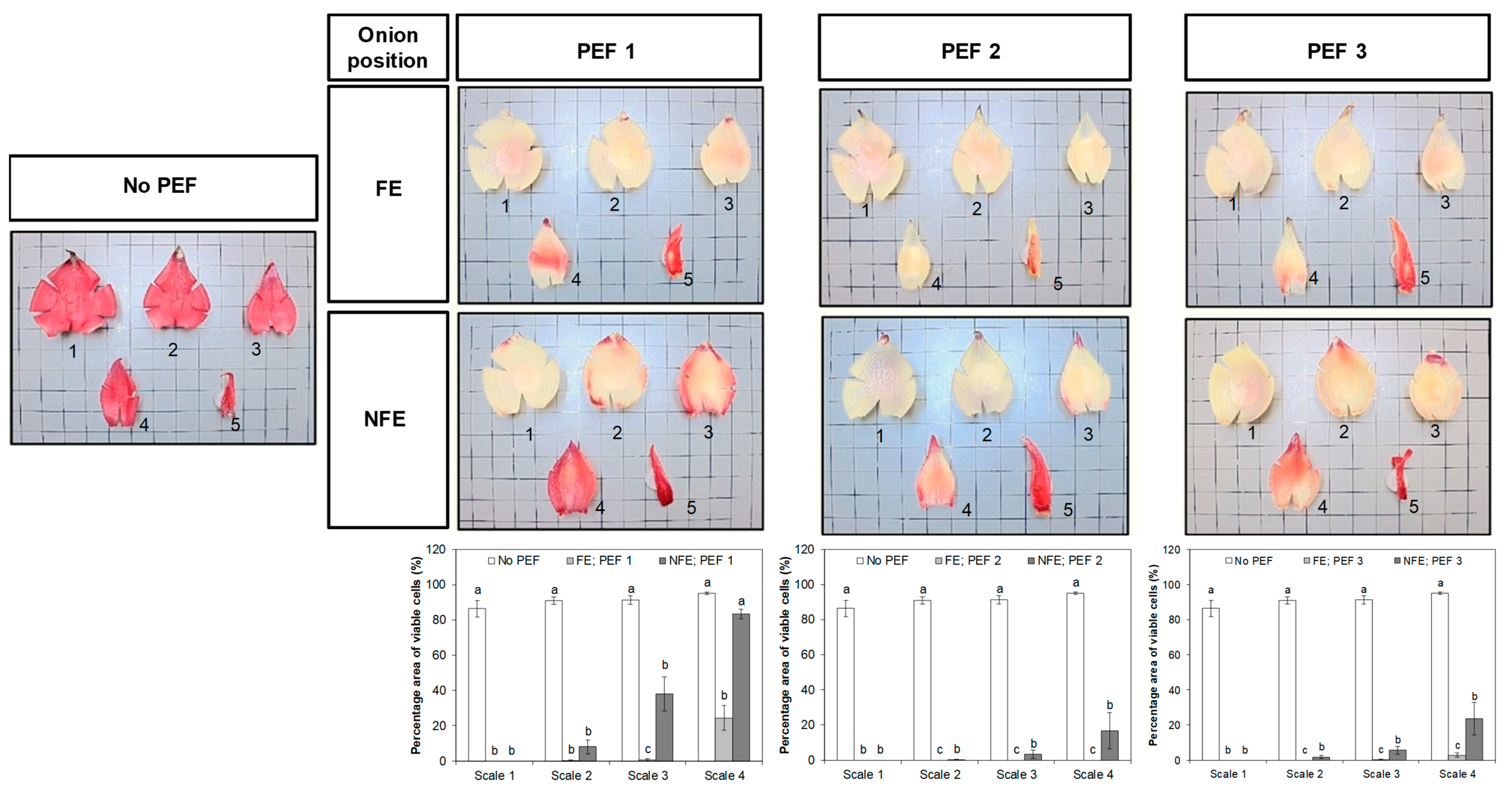
2.4. Determination of Cell Viability Using Tetrazolium Salt as a Staining Agent
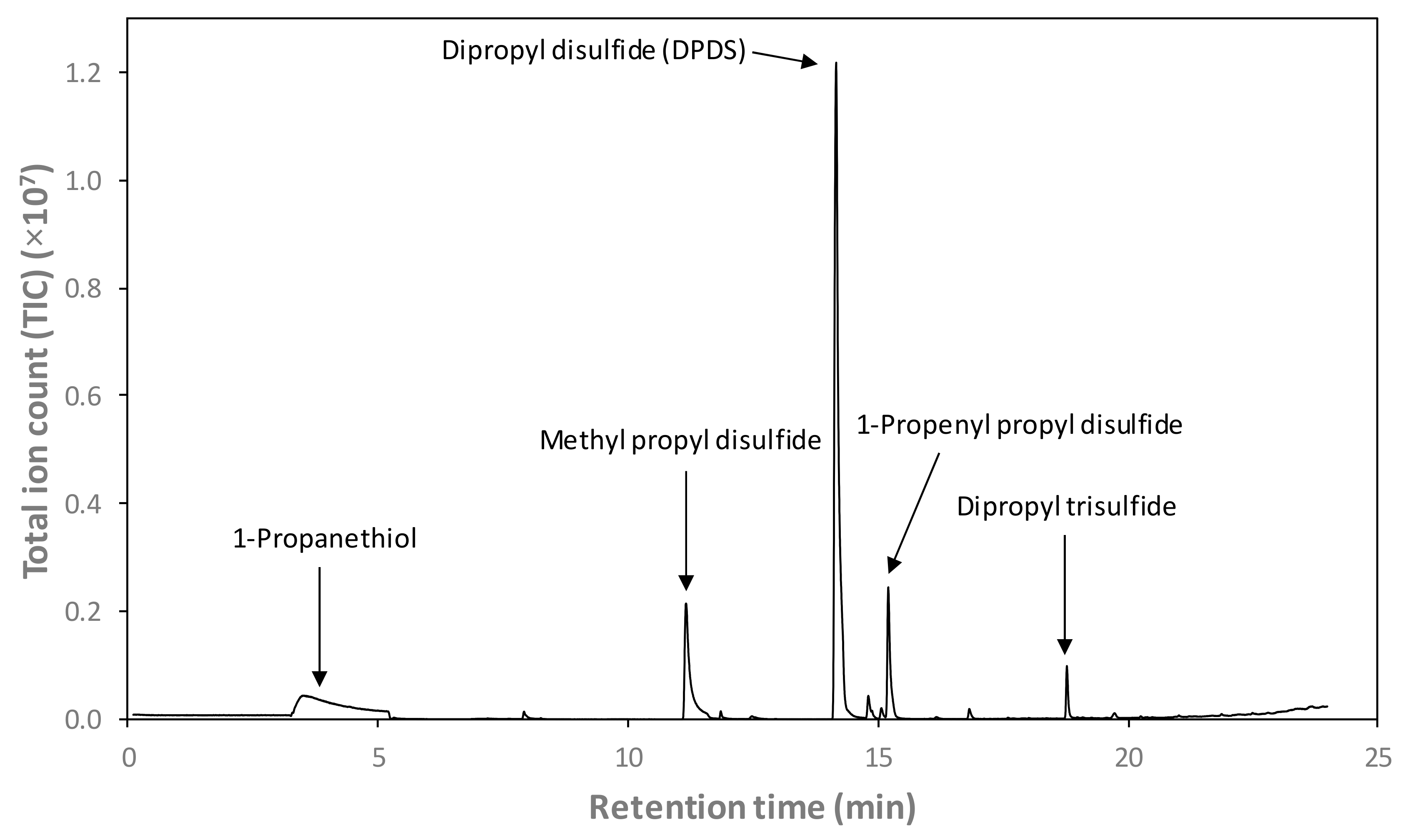
2.5. Determination of Onion Volatile Compounds Using Headspace Solid-Phase Microextraction Coupled with Gas Chromatography–Mass Spectrometry (HS-SPME-GC-MS)
2.6. Kinetic Modelling and Parameter Estimation for Volatiles Release of PEF-Treated Onion Bulbs
2.7. Data Analysis
3. Results
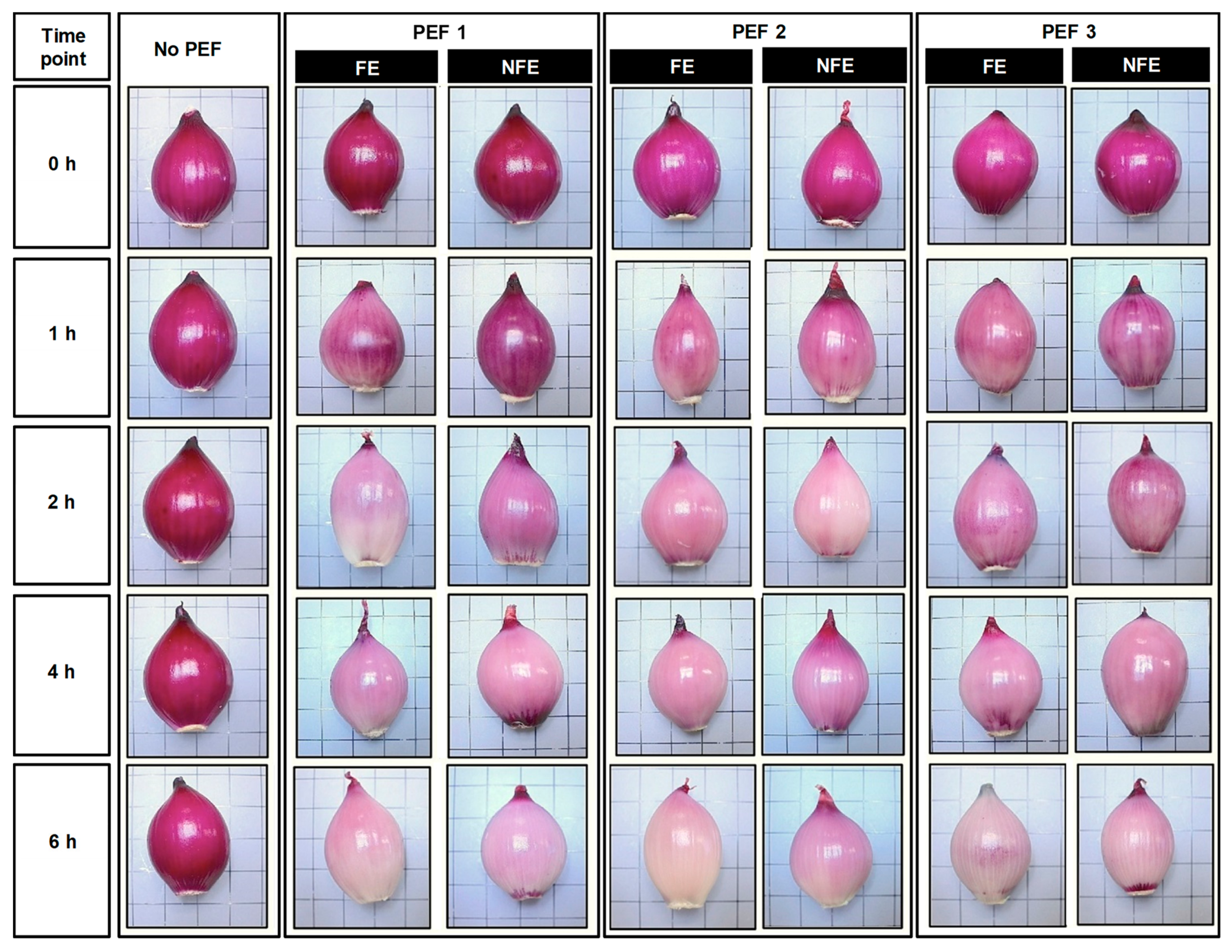
3.1. Effect of PEF on the Changes in the Purplish-Red Pigments Distribution at the Exterior of Onion Bulbs
3.2. Changes in the Onion Epidermal Cells in Response to PEF
3.3. Effect of PEF on the Viability of Onion Cells
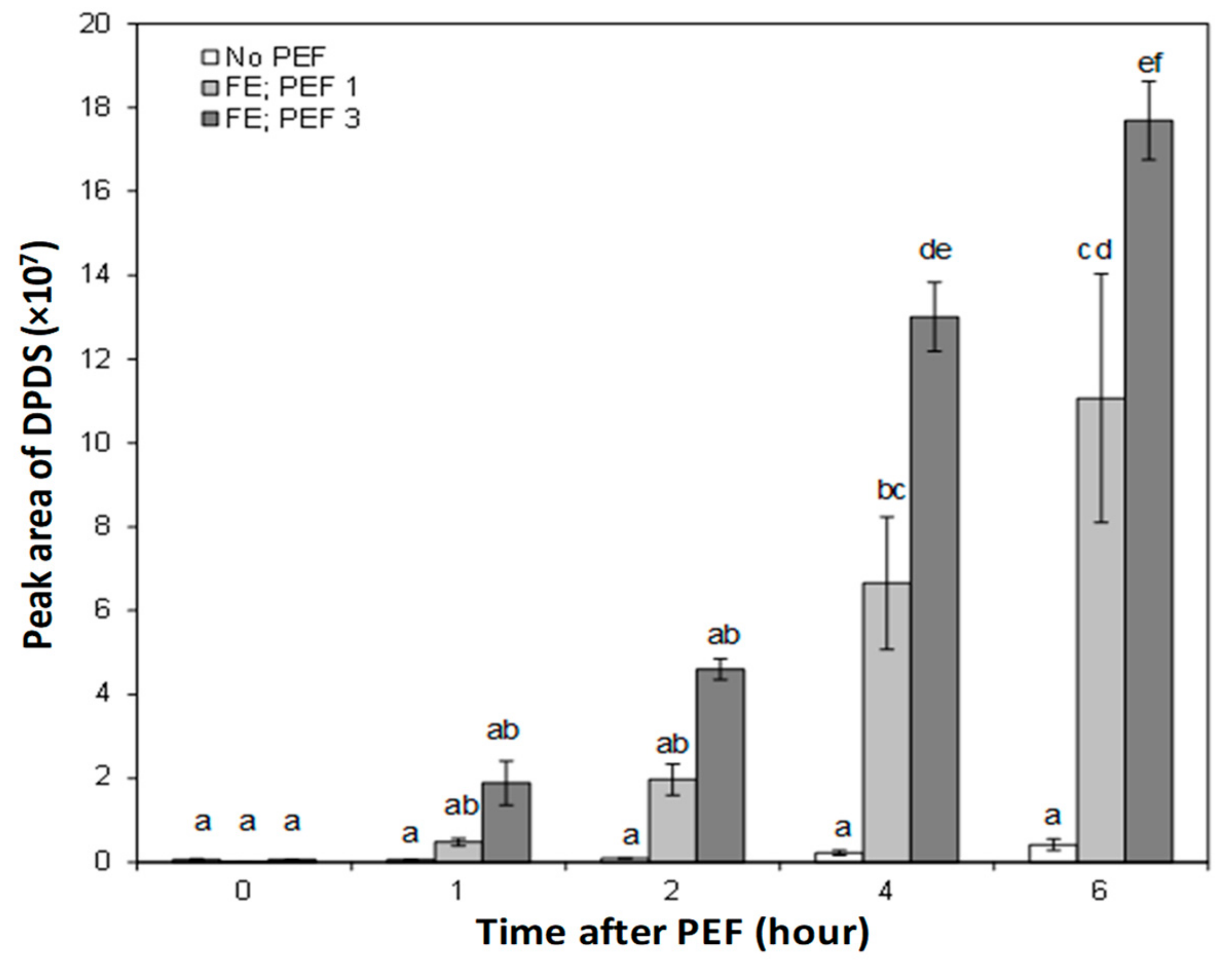
3.4. Effect of PEF on the Concentration of Headspace Onion Volatiles
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Griffiths, G.; Trueman, L.; Crowther, T.; Thomas, B.; Smith, B. Onions—A global benefit to health. Phytother. Res. 2002, 16, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Parkin, K.L. Antioxidant functions of selected Allium thiosulfinates and S-alk(en)yl-L-cysteine sulfoxides. J. Agric. Food Chem. 2002, 50, 2488–2493. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.G.; Hughes, J.; Tregova, A.; Milne, J.; Tomsett, A.B.; Collin, H.A. Biosynthesis of the flavour precursors of onion and garlic. J. Exp. Bot. 2004, 55, 1903–1918. [Google Scholar] [CrossRef] [PubMed]
- Bacon, J.R.; Moates, G.K.; Ng, A.; Rhodes, M.J.; Smith, A.C.; Waldron, K.W. Quantitative analysis of flavour precursors and pyruvate levels in different tissues and cultivars of onion (Allium cepa). Food Chem. 1999, 64, 257–261. [Google Scholar] [CrossRef]
- Gyawali, R.; Seo, H.-Y.; Lee, H.-J.; Song, H.-P.; Kim, D.-H.; Byun, M.-W.; Kim, K.-S. Effect of γ-irradiation on volatile compounds of dried Welsh onion (Allium fistulosum L.). Radiat. Phys. Chem. 2006, 75, 322–328. [Google Scholar] [CrossRef]
- Colina-Coca, C.; González-Peña, D.; Vega, E.; de Ancos, B.; Sánchez-Moreno, C. Novel approach for the determination of volatile compounds in processed onion by headspace gas chromatography-mass spectrometry (HS GC–MS). Talanta 2013, 103, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Angersbach, A.; Heinz, V.; Knorr, D. Effects of pulsed electric fields on cell membranes in real food systems. Innov. Food Sci. Emerg. Technol. 2000, 1, 135–149. [Google Scholar] [CrossRef]
- Faridnia, F.; Burritt, D.J.; Bremer, P.J.; Oey, I. Innovative approach to determine the effect of pulsed electric fields on the microstructure of whole potato tubers: Use of cell viability, microscopic images and ionic leakage measurements. Food Res. Int. 2015, 77, 556–564. [Google Scholar] [CrossRef]
- Fincan, M.; Dejmek, P. In situ visualization of the effect of a pulsed electric field on plant tissue. J. Food Eng. 2002, 55, 223–230. [Google Scholar] [CrossRef]
- Asavasanti, S.; Ersus, S.; Ristenpart, W.; Stroeve, P.; Barrett, D.M. Critical electric field strengths of onion tissues treated by pulsed electric fields. J. Food Sci. 2010, 75, E433–E443. [Google Scholar] [CrossRef]
- Ersus, S.; Oztop, M.H.; McCarthy, M.J.; Barrett, D.M. Disintegration efficiency of pulsed electric field induced effects on onion (Allium cepa L.) tissues as a function of pulse protocol and determination of cell integrity by 1H-NMR relaxometry. J. Food Sci. 2010, 75, E444–E452. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Dodds, E.; Leong, S.Y.; Eyres, G.T.; Burritt, D.J.; Oey, I. Effect of pulsed electric fields on the structure and frying quality of “kumara” sweet potato tubers. Innov. Food Sci. Emerg. Technol. 2017, 39, 197–208. [Google Scholar] [CrossRef]
- Liu, T.; Burritt, D.J.; Eyres, G.T.; Oey, I. Pulsed electric field processing reduces the oxalate content of oca (Oxalis tuberosa) tubers while retaining starch grains and the general structural integrity of tubers. Food Chem. 2018, 245, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Nandakumar, R.; Eyres, G.; Burritt, D.; Kebede, B.; Leus, M.; Oey, I. Impact of Pulsed Electric Fields on the volatile compounds produced in whole onions (Allium cepa and Allium fistulosum). Foods 2018, 7, 183. [Google Scholar] [CrossRef] [PubMed]
- Ersus, S.; Barrett, D.M. Determination of membrane integrity in onion tissues treated by pulsed electric fields: Use of microscopic images and ion leakage measurements. Innov. Food Sci. Emerg. Technol. 2010, 11, 598–603. [Google Scholar] [CrossRef]
- Liu, T.; Burritt, D.J.; Oey, I. Understanding the effect of Pulsed Electric Fields on multilayered solid plant foods: Bunching onions (Allium fistulosum) as a model system. Food Res. Int. 2019, 120, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.V.; Herst, P.M.; Tan, A.S. Tetrazolium dyes as tools in cell biology: New insights into their cellular reduction. Biotechnol. Annu. Rev. 2005, 11, 127–152. [Google Scholar] [PubMed]
- Lanzotti, V. The analysis of onion and garlic. J. Chromatogr. A 2006, 1112, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Van Boekel, M.A.J.S. Kinetic modeling of food quality: A critical review. Compr. Rev. Food Sci. Food Saf. 2008, 7, 144–158. [Google Scholar] [CrossRef]
- Leong, S.Y.; Oey, I. Effect of endogenous ascorbic acid oxidase activity and stability on vitamin C in carrots (Daucus carota subsp. sativus) during thermal treatment. Food Chem. 2012, 134, 2075–2085. [Google Scholar] [CrossRef]
- Terahara, N.; Yamaguchi, M.-A.; Honda, T. Malonylated anthocyanins from bulbs of red onion, Allium cepa L. Biosci. Biotechnol. Biochem. 1994, 58, 1324–1325. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Ovando, A.; Pacheco-Hernández, M.D.L.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical studies of anthocyanins: A review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Gonzalez, M.; Jernstedt, J.; Slaughter, D.; Barrett, D. Microscopic quantification of cell integrity in raw and processed onion parenchyma cells. J. Food Sci. 2010, 75, E402–E408. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Gutiérrez, J.L.; Plaza, L.; Hernando, I.; Sánchez-Moreno, C.; Quiles, A.; de Ancos, B.; Cano, M.P. Changes in the structure and antioxidant properties of onions by high pressure treatment. Food Funct. 2013, 4, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Valič, B.; Golzio, M.; Pavlin, M.; Schatz, A.; Faurie, C.; Gabriel, B.; Teissié, J.; Rols, M.-P.; Miklavčič, D. Effect of electric field induced transmembrane potential on spheroidal cells: Theory and experiment. Eur. Biophys. J. 2003, 32, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Ng, A.; Smith, A.C.; Waldron, K.W. Effect of tissue type and variety on cell wall chemistry of onion (Allium cepa L.). Food Chem. 1998, 63, 17–24. [Google Scholar] [CrossRef]
- Boelens, M.; De Valois, P.J.; Wobben, H.J.; Van der Gen, A. Volatile flavor compounds from onion. J. Agric. Food Chem. 1971, 19, 984–991. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PEF Treatment | Electric Field Strength (kV/cm) | Pulse Width (µs) | Pulse Frequency (Hz) | Pulse Number | Specific Energy Input (kJ/kg) |
|---|---|---|---|---|---|
| PEF1 | 0.6 | 20 | 50 | 540 | 6 |
| PEF2 | 0.6 | 20 | 50 | 5400 | 60 |
| PEF3 | 1.2 | 20 | 50 | 120 | 6 |
| PEF Treatment | k (×107 Peak Area Per Hour) | Corrected R2 |
|---|---|---|
| No PEF | 0.06 ± 0.01 | 0.6805 |
| FE; PEF 1 | 1.94 ± 0.29 | 0.7589 |
| FE; PEF 3 | 3.12 ± 0.15 | 0.9671 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tantamacharik, T.; Leong, S.Y.; Leus, M.J.; Eyres, G.T.; Burritt, D.J.; Oey, I. Structural Changes Induced by Pulsed Electric Fields Increase the Concentration of Volatiles Released in Red Onion (Allium cepa L. var. Red Pearl) Bulbs. Foods 2019, 8, 368. https://doi.org/10.3390/foods8090368
Tantamacharik T, Leong SY, Leus MJ, Eyres GT, Burritt DJ, Oey I. Structural Changes Induced by Pulsed Electric Fields Increase the Concentration of Volatiles Released in Red Onion (Allium cepa L. var. Red Pearl) Bulbs. Foods. 2019; 8(9):368. https://doi.org/10.3390/foods8090368
Chicago/Turabian StyleTantamacharik, Thirawat, Sze Ying Leong, Michelle J. Leus, Graham T. Eyres, David J. Burritt, and Indrawati Oey. 2019. "Structural Changes Induced by Pulsed Electric Fields Increase the Concentration of Volatiles Released in Red Onion (Allium cepa L. var. Red Pearl) Bulbs" Foods 8, no. 9: 368. https://doi.org/10.3390/foods8090368
APA StyleTantamacharik, T., Leong, S. Y., Leus, M. J., Eyres, G. T., Burritt, D. J., & Oey, I. (2019). Structural Changes Induced by Pulsed Electric Fields Increase the Concentration of Volatiles Released in Red Onion (Allium cepa L. var. Red Pearl) Bulbs. Foods, 8(9), 368. https://doi.org/10.3390/foods8090368