
Step Enzymatic Hydrolysis and In Silico Screening-Assisted Preparation of Bioactive Peptides from Abalone

Abstract

1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Abalone Dry Powder and Abalone Enzymatic Hydrolysates
2.3. Optimization of Enzymatic Hydrolysis of Abalone
2.4. HPLC Peptide MW Distribution
2.5. Separation of Enzymatic Hydrolysis Products and Determination of Amino Acid Composition
2.6. Determination of ACE Inhibitory Activity and ABTS Antioxidant Activity
2.7. Peptidomic Analysis
2.8. Molecular Docking
2.9. PeptideRanker Tool for Screening Potential Bioactive Peptides
2.10. Statistical Analysis
3. Results
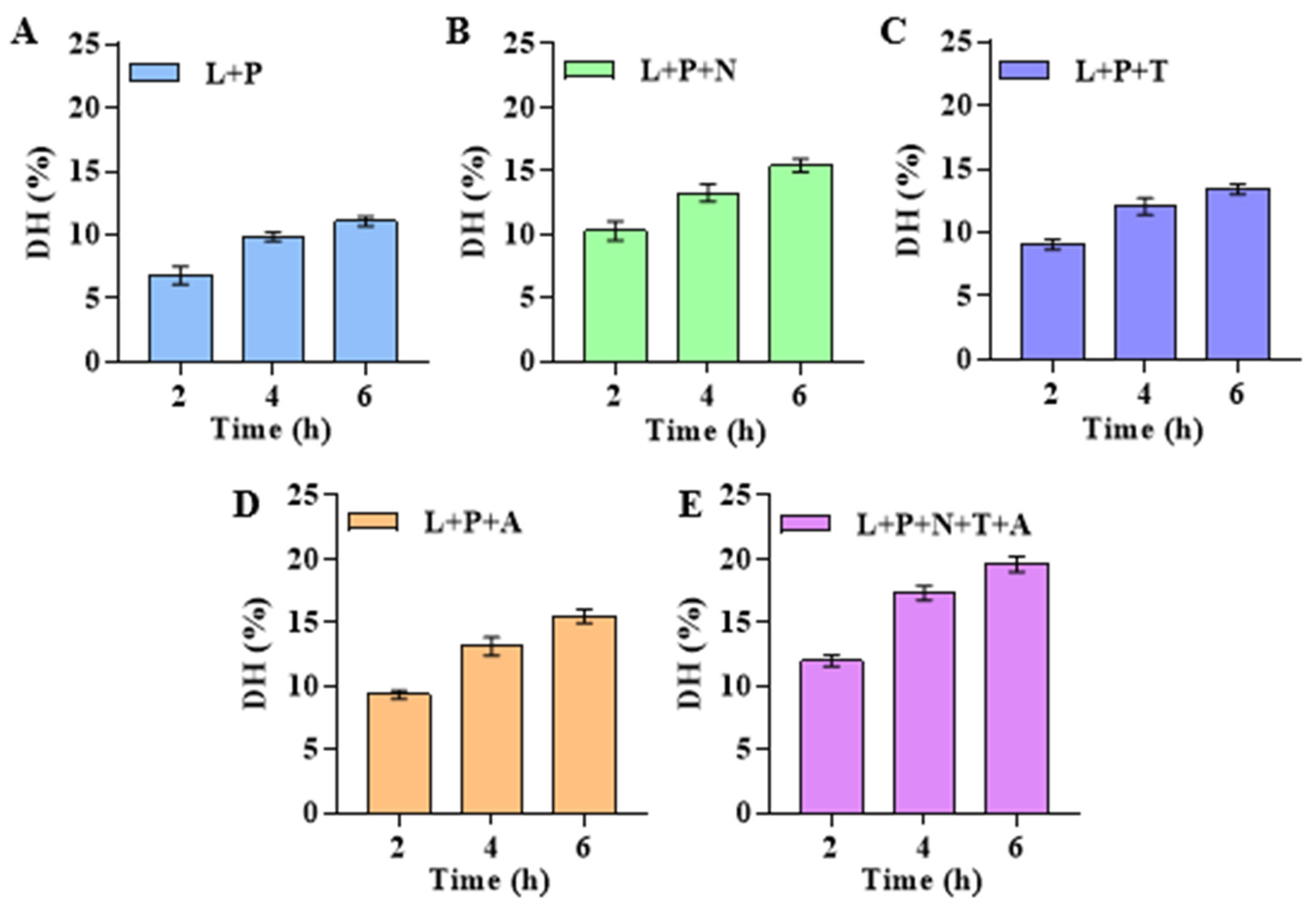
3.1. Establishment and Optimization of Enzymatic Hydrolysis Process
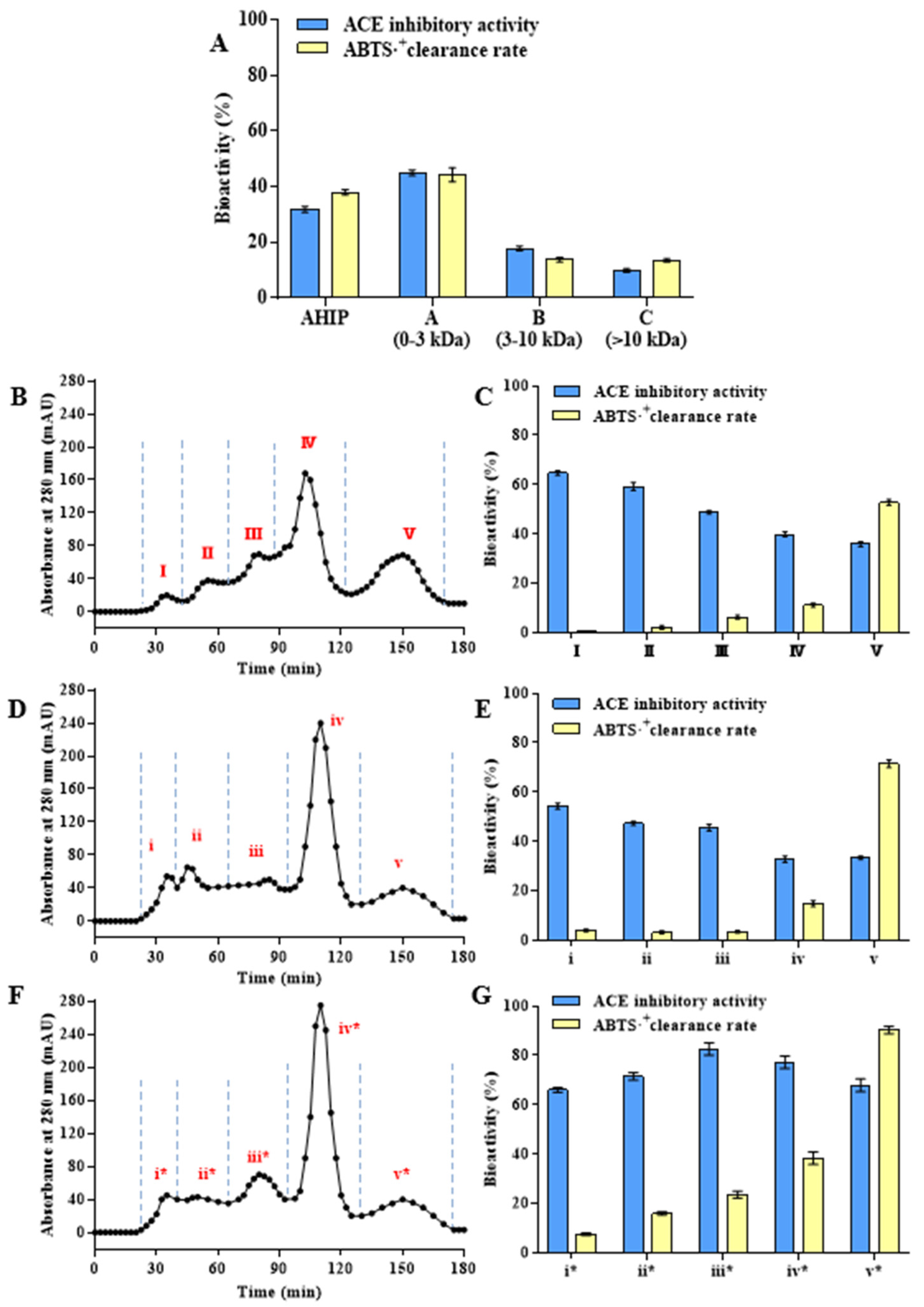
3.2. Separation and Analysis of Bioactive Abalone Peptides
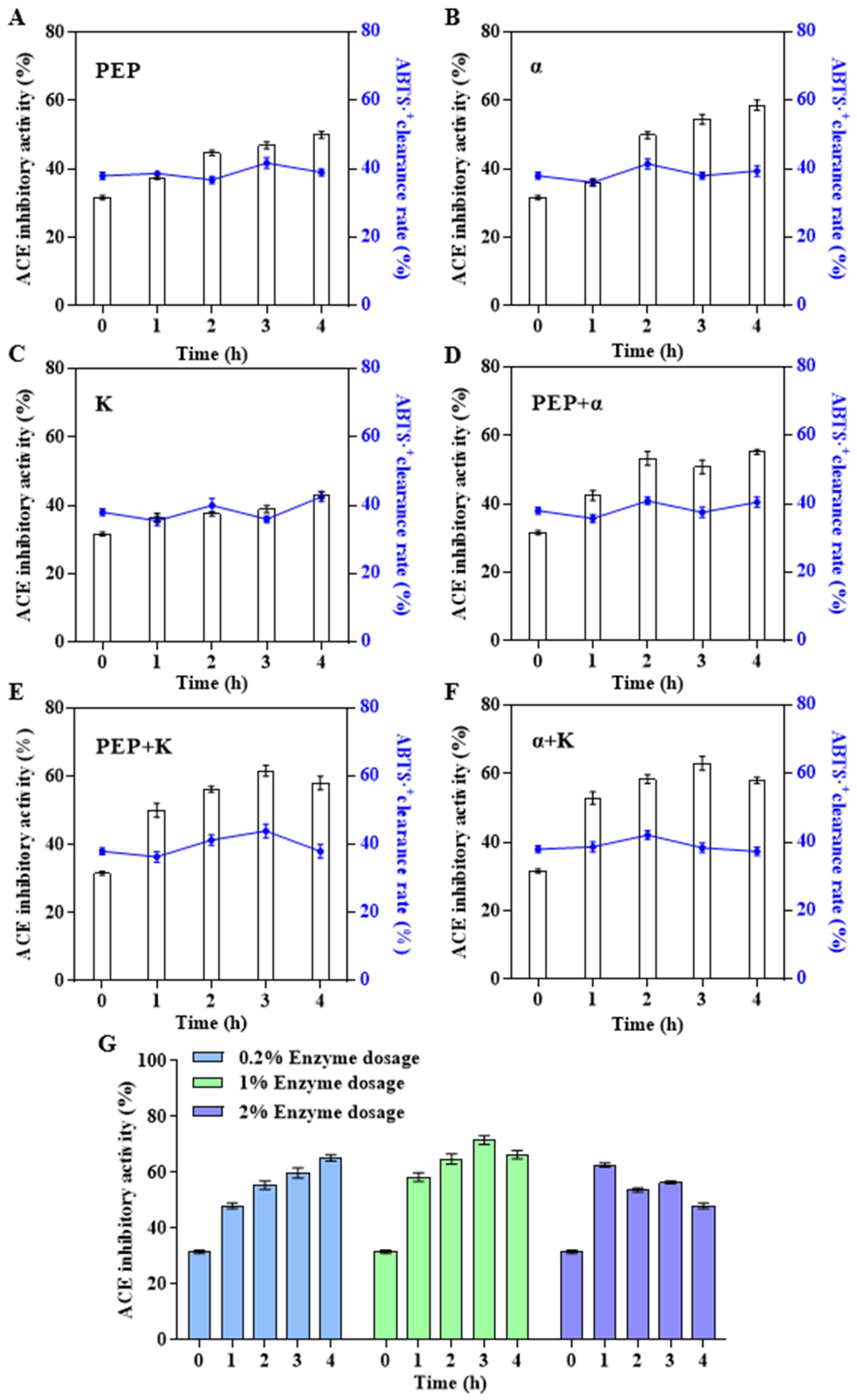
3.3. Improvement of Bioactivity of Abalone Polypeptides by Secondary Enzymatic Hydrolysis with Specific Proteases
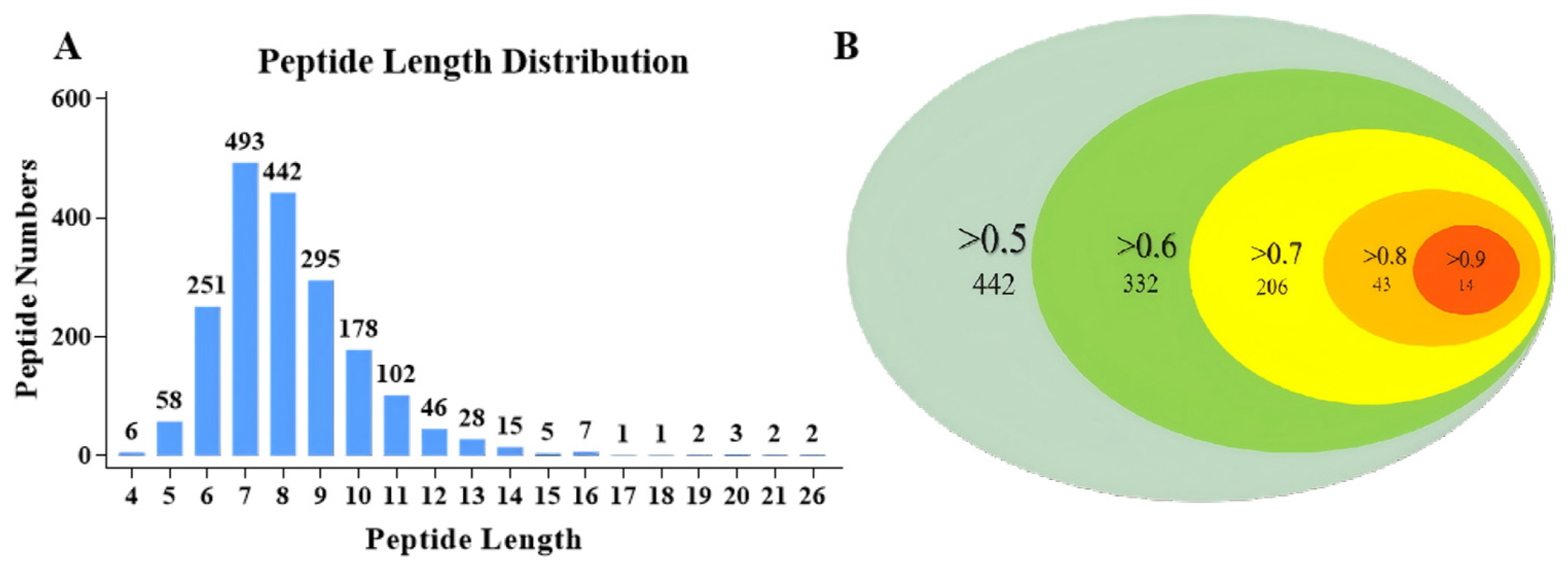
3.4. Peptidomic Analysis of APSH-iii* and APSH-iv*
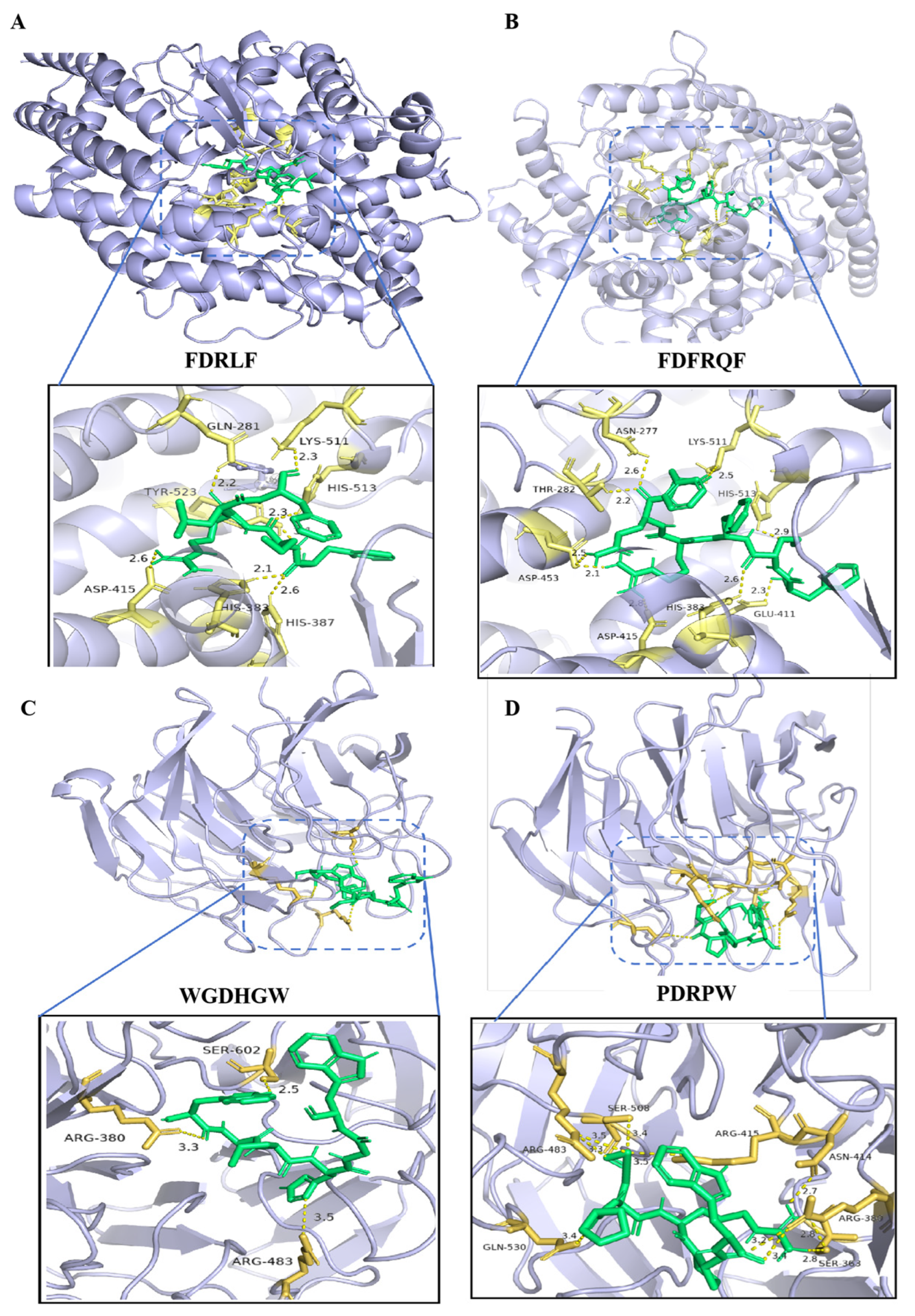
3.5. Virtual Screening of Bioactive Peptides Using Bioinformatics Tools and Molecular Docking
3.6. Peptide Synthesis and Bioactivity Verification by Wet Experiment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghosh, S.; Sarkar, T.; Pati, S.; Kari, Z.A.; Edinur, H.A.; Chakraborty, R. Novel Bioactive Compounds from Marine Sources as a Tool for Functional Food Development. Front. Mar. Sci. 2022, 9, 832957. [Google Scholar] [CrossRef]
- Yan, H.; Chu, Y.; Li, W.; Zhang, D.; Yang, Q.; Wei, Z.; Wang, Z.; Wang, Y.; Zhu, L. Advances in Research on Biological and Functional Properties of Bioactive Peptides Derived from Marine Sources. Food Sci. 2023, 44, 18–28. [Google Scholar] [CrossRef]
- Suo, S.K.; Zhao, Y.Q.; Wang, Y.M.; Pan, X.Y.; Chi, C.F.; Wang, B. Seventeen novel angiotensin converting enzyme (ACE) inhibitory peptides from the protein hydrolysate of Mytilus edulis: Isolation, identification, molecular docking study, and protective function on HUVECs. Food Funct. 2022, 13, 7831–7846. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, X.; Li, F.; Wang, W.; Leng, K.; Li, Y.; Guo, X. Optimization of Enzymatic Hydrolysis Preparation Process and Stability Evaluation of ACE Inhibitory Peptides from Antarctic Krill (Euphausia superba). Sci. Technol. Food Ind. 2023, 21, 243–250. [Google Scholar] [CrossRef]
- Cunha, S.A.; Pintado, M.E. Bioactive peptides derived from marine sources: Biological and functional properties. Trends Food Sci. Technol. 2022, 119, 348–370. [Google Scholar] [CrossRef]
- Lv, R.Z.; Dong, Y.F.; Bao, Z.J.; Zhang, S.M.; Lin, S.Y.; Sun, N. Advances in the activity evaluation and cellular regulation pathways of food-derived antioxidant peptides. Trends Food Sci. Technol. 2022, 122, 171–186. [Google Scholar] [CrossRef]
- Liu, X.; Chen, S.; Li, L.; Yang, X.; Huang, H. Nutritional analysis and quality evaluation of four kinds of abalone muscle. Food Ferment. Ind. 2018, 44, 227–231. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Masci, P.P.; Gobe, G.C.; Osborne, S.A. Therapeutic potential of abalone and status of bioactive molecules: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2017, 57, 1742–1748. [Google Scholar] [CrossRef]
- He, Z.; Zhang, Y.; Zeng, J.; Lu, C.; Li, P.; Du, B. Research Progress on the Nutritional Composition, Functional Activities and Processing of Abalone. Sci. Technol. Food Ind. 2023, 15, 440–450. [Google Scholar] [CrossRef]
- Guo, S.; Wang, J.; He, C.B.; Wei, H.C.; Ma, Y.; Xiong, H.J. Preparation and antioxidant activities of polysaccharides obtained from abalone viscera by combination of enzymolysis and multiple separation methods. J. Food Sci. 2020, 85, 4260–4270. [Google Scholar] [CrossRef]
- Chen, S.Y.; Xiao, J.B.; Su, J.Q. Preliminary study on immune function of polysaccharide extract from abalone viscera. Light Text. Ind. Fujian 2023, 2, 7–12. [Google Scholar] [CrossRef]
- Xiao, J.B.; Chen, S.Y.; Chen, Y.Q.; Su, J.Q. Preliminary study on enhancing immune function of abalone visceral lipids. Light Text. Ind. Fujian 2022, 2, 7–11. [Google Scholar] [CrossRef]
- Yang, Z.; Zhao, X.; Ye, J.; Xu, H.; Chen, J. Study on extraction and purification process and antioxidant characteristics of abalone visceral phospholipids. J. Henan Univ. Technol. Nat. Sci. Ed. 2023, 44, 11–17. [Google Scholar] [CrossRef]
- Li, G.Y.; Zuo, X.; Luo, X.L.; Chen, Z.Q.; Cao, W.H.; Lin, H.S.; Qin, X.M.; Wu, L.Y.; Zheng, H.A. Functional, physicochemical, and structural properties of the hydrolysates derived from the abalone (Haliotis discus subsp hannai Ino) foot muscle proteins. Food Chem. X 2023, 19, 12. [Google Scholar] [CrossRef] [PubMed]
- Toldrá, F.; Reig, M.; Aristoy, M.C.; Mora, L. Generation of bioactive peptides during food processing. Food Chem. 2018, 267, 395–404. [Google Scholar] [CrossRef]
- Su, Y.J.; Liao, H.J.; Yang, J.I. Purification and Identification of an ACE-Inhibitory Peptide from Gracilaria tenuistipitata Protein Hydrolysates. Processes 2022, 10, 1128. [Google Scholar] [CrossRef]
- Tang, P.; Han, Q.; Bao, B.; Bao, C.; Wu, W. Research Progress on Preparation, Separation, Purification and Structure-Function Relationship of Marine Bioactive Peptides. Sci. Technol. Food Ind. 2022, 24, 488–495. [Google Scholar] [CrossRef]
- Zheng, S.L.; Luo, Q.B.; Suo, S.K.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Preparation, Identification, Molecular Docking Study and Protective Function on HUVECs of Novel ACE Inhibitory Peptides from Protein Hydrolysate of Skipjack Tuna Muscle. Mar. Drugs 2022, 20, 21. [Google Scholar] [CrossRef]
- Gu, Y.X.; Zhang, J.C.; Niu, Y.J.; Sun, B.G.; Liu, Z.Y.; Mao, X.Z.; Zhang, Y.Y. Screening and Characterization of novel umami peptides in Cheddar cheese using peptidomics and bioinformatics approaches. LWT-Food Sci. Technol. 2024, 194, 9. [Google Scholar] [CrossRef]
- Nielsen, P.M.; Petersen, D.; Dambmann, C. Improved method for determining food protein degree of hydrolysis. J. Food Sci. 2001, 66, 642–646. [Google Scholar] [CrossRef]
- Wei, M.R.; Liu, K.Z.; Yang, X.M.; Li, J.H.; Zhang, G.Q.; Du, G.C. Application of protein glutaminase in the enzymatic preparation of whey protein peptides. Food Ferment. Ind. 2023, 1, 16–24. [Google Scholar] [CrossRef]
- Mæhre, H.K.; Dalheim, L.; Edvinsen, G.K.; Elvevoll, E.O.; Jensen, I.J. Protein DeterminationMethod Matters. Foods 2018, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.F.; Luo, J.Y.; Lin, J.R.; Meng, C.; Hong, J. Preparation of angiotensin I-converting enzyme (ACE) inhibitory peptides from Tie Guanyin tea residue protein using two-step enzymatic hydrolysis. Food Sci. Technol. 2022, 42, 13. [Google Scholar] [CrossRef]
- Wang, L.Y.; Ma, M.T.; Yu, Z.P.; Du, S.K. Preparation and identification of antioxidant peptides from cottonseed proteins. Food Chem. 2021, 352, 9. [Google Scholar] [CrossRef]
- Wei, G.Q.; Zhao, Q.; Wang, D.D.; Fan, Y.Z.; Shi, Y.A.; Huang, A.X. Novel ACE inhibitory, antioxidant and a-glucosidase inhibitory peptides identified from fermented rubing cheese through peptidomic and molecular docking. LWT-Food Sci. Technol. 2022, 159, 11. [Google Scholar] [CrossRef]
- Montone, C.M.; Capriotti, A.L.; Cavaliere, C.; La Barbera, G.; Piovesana, S.; Chiozzi, R.Z.; Laganà, A. Peptidomic strategy for purification and identification of potential ACE-inhibitory and antioxidant peptides in Tetradesmus obliquus microalgae. Anal. Bioanal. Chem. 2018, 410, 3573–3586. [Google Scholar] [CrossRef]
- de Castro, R.J.S.; Sato, H.H. Biologically active peptides: Processes for their generation, purification and identification and applications as natural additives in the food and pharmaceutical industries. Food Res. Int. 2015, 74, 185–198. [Google Scholar] [CrossRef]
- Sitanggang, A.B.; Putri, J.E.; Palupi, N.S.; Hatzakis, E.; Syamsir, E.; Budijanto, S. Enzymatic Preparation of Bioactive Peptides Exhibiting ACE Inhibitory Activity from Soybean and Velvet Bean: A Systematic Review. Molecules. 2021, 26, 3822. [Google Scholar] [CrossRef]
- Angulo, M.; Márquez, M.C. A Green Technology Approach Using Enzymatic Hydrolysis to Valorize Meat Waste as a Way to Achieve a Circular Economy. Appl. Sci. 2023, 13, 8763. [Google Scholar] [CrossRef]
- Sandra, M.P.; Diego, B. Enzymatic concentration in leaves, bark and latex of papaya and its softening effect in bovine meat. Novasinergi. 2018, 1, 72–79. [Google Scholar] [CrossRef]
- Liu, Q.; Yao, Y.S.; Ibrahim, M.A.A.; El Halawany, A.M.; Yang, L.; Zhang, X.W. Production of Dual Inhibitory Hydrolysate by Enzymatic Hydrolysis of Squid Processing By-product. Mar. Biotechnol. 2022, 24, 293–302. [Google Scholar] [CrossRef]
- Wang, Y.M.; Li, X.Y.; Wang, J.; He, Y.; Chi, C.F.; Wang, B. Antioxidant peptides from protein hydrolysate of skipjack tuna milt: Purification, identification, and cytoprotection on H2O2 damaged human umbilical vein endothelial cells. Process Biochem. 2022, 113, 258–269. [Google Scholar] [CrossRef]
- Yao, Y.X.; Xie, X.; Yan, S.Z.; Guan, Y.; Ma, H.F.; Chen, H.; Sun, S.K. Effect of Enzymatic Hydrolysis Duration on Structure and Functional Properties of Soybean Protein Isolate. J. Food Sci. Biotechnol. 2024, 43, 135–143. [Google Scholar] [CrossRef]
- Wang, Q.; Shi, C.Y.; Qian, Y.M.; Li, H.X.; Chen, H.L.; Chen, X.X.; Cao, W.G. Effects of hydrolysis time on antioxidant activity of tilapia skin collagen proteolytic. J. Hunan Univ. Arts Sci. 2021, 33, 36–41. [Google Scholar] [CrossRef]
- Zhao, M.; Wu, C.Y. Optimization of enzymatic hydrolysis of tilapia waste by using response surface methodology. Food Res. Dev. 2007, 1, 48–53. [Google Scholar] [CrossRef]
- Ahmed, I.; Asgher, M.; Sher, F.; Hussain, S.M.; Nazish, N.; Joshi, N.; Sharma, A.; Parra-Saldívar, R.; Bilal, M.; Iqbal, H.M.N. Exploring Marine as a Rich Source of Bioactive Peptides: Challenges and Opportunities from Marine Pharmacology. Mar. Drugs 2022, 20, 208. [Google Scholar] [CrossRef]
- Wang, J.; Wu, Y.; Yang, X. Review on research status and application prospect of bioactive peptides from marine shellfish. Sci. Technol. Food Ind. 2013, 34, 346–349. [Google Scholar] [CrossRef]
- Zaky, A.A.; Simal-Gandara, J.; Eun, J.B.; Shim, J.H.; Abd El-Aty, A.M. Bioactivities, Applications, Safety, and Health Benefits of Bioactive Peptides From Food and By-Products: A Review. Front. Nutr. 2022, 8, 815640. [Google Scholar] [CrossRef]
- Dong, X.M.; Suo, S.K.; Wang, Y.M.; Zeng, Y.H.; Chi, C.F.; Wang, B. High Fischer ratio oligopeptides from Antarctic krill: Ameliorating function and mechanism to alcoholic liver injury through regulating AMPK/Nrf2/IκBα pathways. J. Funct. Foods 2024, 122, 12. [Google Scholar] [CrossRef]
- Mao, Z.J.; Jiang, H.; Sun, J.A.; Zhao, Y.H.; Gao, X.; Mao, X.Z. Research progress in the preparation and structure-activity relationship of bioactive peptides derived from aquatic foods. Trends Food Sci. Technol. 2024, 147, 14. [Google Scholar] [CrossRef]
- Tu, M.L.; Wang, C.; Chen, C.; Zhang, R.Y.; Liu, H.X.; Lu, W.H.; Jiang, L.Z.; Du, M. Identification of a novel ACE-inhibitory peptide from casein and evaluation of the inhibitory mechanisms. Food Chem. 2018, 256, 98–104. [Google Scholar] [CrossRef]
- García-Horsman, J.A. The role of prolyl oligopeptidase, understanding the puzzle. Ann. Transl. Med. 2020, 8, 3. [Google Scholar] [CrossRef]
- Ma, M.; Feng, Y.; Miao, Y.; Shen, Q.; Tang, S.; Dong, J.; Zhang, J.Z.H.; Zhang, L. Revealing the Sequence Characteristics and Molecular Mechanisms of ACE Inhibitory Peptides by Comprehensive Characterization of 160,000 Tetrapeptides. Foods 2023, 12, 1573. [Google Scholar] [CrossRef]
- Ulug, S.K.; Jahandideh, F.; Wu, J.P. Novel technologies for the production of bioactive peptides. Trends Food Sci. Technol. 2021, 108, 27–39. [Google Scholar] [CrossRef]
- Ferreira, R.; Amado, F.; Vitorino, R. Empowering peptidomics: Utilizing computational tools and approaches. Bioanalysis 2023, 15, 1315–1325. [Google Scholar] [CrossRef] [PubMed]
- Bianca, d.F.G.; Márcio, d.B.; Thaís, d.S.R. Bioactive peptides from beans with the potential to decrease the risk of developing noncommunicable chronic diseases. Crit. Rev. Food Sci. Nutr. 2020, 61, 2003–2021. [Google Scholar] [CrossRef]
- Longarela, N.R.; Ramos, M.P.; Vilariño, J.M.L. Bioinformatics tools for the study of bioactive peptides from vegetal sources: Evolution and future perspectives. Crit. Rev. Food Sci. Nutr. 2024, 1, 20. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef]
- Zamyatnin, A.A.; Borchikov, A.S.; Vladimirov, M.G.; Voronina, O.L. The EROP-Moscow oligopeptide database. Nucleic Acids Res. 2006, 34, D261–D266. [Google Scholar] [CrossRef]
- Du, Z.J.; Comer, J.; Li, Y.H. Bioinformatics approaches to discovering food-derived bioactive peptides: Reviews and perspectives. TrAC Trends Anal. Chem. 2023, 162, 29. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Levels | Factors | |||
|---|---|---|---|---|
| Hydrolysis Time/(h) | Hydrolysis Temperature /(°C) | Solid–Liquid Ratio | Enzyme Dosage /(%) | |
| 1 | 3 | 37 | 1:10 | 1 |
| 2 | 4 | 46 | 1:20 | 2 |
| 3 | 5 | 55 | 1:30 | 3 |
| Experimental Serial Number | Factors | DH/% | Polypeptide Content/% | Comprehensive Score /(Points) | |||
|---|---|---|---|---|---|---|---|
| Hydrolysis Time/(h) | Hydrolysis Temperature/(°C) | Solid–Liquid Ratio | Enzyme Dosage /(%) | ||||
| 1 | 1 | 1 | 1 | 1 | 11.05 | 78.92 | 66.15 |
| 2 | 2 | 2 | 2 | 1 | 15.82 | 81.59 | 75.81 |
| 3 | 3 | 3 | 3 | 1 | 20.35 | 83.07 | 84.34 |
| 4 | 3 | 2 | 1 | 2 | 20.78 | 70.93 | 77.76 |
| 5 | 1 | 3 | 2 | 2 | 19.94 | 76.52 | 79.71 |
| 6 | 2 | 1 | 3 | 2 | 16.66 | 82.01 | 77.48 |
| 7 | 2 | 3 | 1 | 3 | 29.63 | 63.40 | 88.16 |
| 8 | 3 | 1 | 2 | 3 | 19.99 | 75.52 | 79.19 |
| 9 | 1 | 2 | 3 | 3 | 21.58 | 78.54 | 83.69 |
| K1 | 76.517 | 74.273 | 77.357 | 75.433 | |||
| K2 | 80.483 | 79.087 | 78.237 | 78.317 | |||
| K3 | 80.430 | 84.070 | 81.837 | 83.880 | |||
| R | 3.966 | 9.797 | 4.48 | 8.447 | |||
| Validation Experiment Number | DH/% | Polypeptide Content/% |
|---|---|---|
| 1 | 21.13 | 70.72 |
| 2 | 21.61 | 73.44 |
| 3 | 22.53 | 73.19 |
| RSD/% | 3.27 | 2.08 |
| AA Species | AA Content/[g·(100 g)−1] | |
|---|---|---|
| A-I (2–3 kDa) | A-V (0–1 kDa) | |
| Asp | 18.64 | 13.21 |
| Glu | 17.10 | 9.49 |
| Ser | 10.44 | 5.88 |
| His | 4.87 | 5.00 |
| Gly | 17.98 | 5.03 |
| Thr | 8.60 | 4.82 |
| Arg | 4.64 | 2.00 |
| Ala | 7.29 | 1.82 |
| Tyr | 4.20 | 19.94 |
| Cys-s | 1.76 | 13.11 |
| Val | 5.76 | 2.49 |
| Met | 3.51 | 0.79 |
| Phe | 2.90 | 23.30 |
| Ile | 3.03 | 1.94 |
| Leu | 5.94 | 3.05 |
| Lys | 6.45 | 1.01 |
| Pro | 13.67 | 3.23 |
| Trp | 3.83 | 0.58 |
| Peptide | Mass/ (Da) | PeptideRanker | Binding Energy with ACE Enzyme /(kcal·mol−1) | Binding Energy with Keap1 Protein /(kcal·mol−1) | ACEi IC50/ (mg·mL−1) | ABTS+ EC50/ (mg·mL−1) |
|---|---|---|---|---|---|---|
| FDRLF | 696 | 0.964558 | −8.5 | −6.8 | 0.05 | - |
| SPPFFDGMTR | 1154 | 0.951409 | −8.1 | −6.7 | 0.18 | - |
| SYPPLGRF | 935 | 0.947818 | −8.6 | −6.2 | 0.20 | 0.17 |
| FDFRQF | 858 | 0.945513 | −7.6 | −6.4 | 0.12 | - |
| GFDFRQF | 915 | 0.942461 | −6.2 | −6.5 | 0.29 | - |
| WGDHGW | 756 | 0.937728 | −7.2 | −7.0 | 0.54 | 0.14 |
| PEHFPF | 772 | 0.937014 | −9.7 | −7.8 | 0.40 | - |
| SSYPPLGRF | 1023 | 0.932763 | −8.6 | −6.8 | 0.15 | 0.16 |
| WPNRPP | 765 | 0.9292 | −5.8 | −6.8 | 0.29 | 0.18 |
| WSDRIPF | 919 | 0.921708 | −7.3 | −6.8 | 0.17 | 0.23 |
| GWDKFWPE | 1063 | 0.920339 | −7.3 | −7.1 | 0.33 | 0.42 |
| PDRPW | 669 | 0.917602 | −7.6 | −7.6 | 0.33 | 0.12 |
| FYDHIF | 840 | 0.917509 | −8.9 | −7.1 | 0.37 | 1.08 |
| ADWDFLPAK | 1062 | 0.910647 | −8.2 | −7.6 | 0.23 | 0.34 |
| SP [18] | 202 | - | - | - | 0.06 | - |
| VDRYF [18] | 699 | - | - | - | 0.28 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, K.; Pang, C.; Li, Q.; Li, J.; Du, G.; Zhang, G. Step Enzymatic Hydrolysis and In Silico Screening-Assisted Preparation of Bioactive Peptides from Abalone. Foods 2025, 14, 1209. https://doi.org/10.3390/foods14071209
Liu K, Pang C, Li Q, Li J, Du G, Zhang G. Step Enzymatic Hydrolysis and In Silico Screening-Assisted Preparation of Bioactive Peptides from Abalone. Foods. 2025; 14(7):1209. https://doi.org/10.3390/foods14071209
Chicago/Turabian StyleLiu, Kanzhen, Cuiping Pang, Qinghua Li, Jianghua Li, Guocheng Du, and Guoqiang Zhang. 2025. "Step Enzymatic Hydrolysis and In Silico Screening-Assisted Preparation of Bioactive Peptides from Abalone" Foods 14, no. 7: 1209. https://doi.org/10.3390/foods14071209
APA StyleLiu, K., Pang, C., Li, Q., Li, J., Du, G., & Zhang, G. (2025). Step Enzymatic Hydrolysis and In Silico Screening-Assisted Preparation of Bioactive Peptides from Abalone. Foods, 14(7), 1209. https://doi.org/10.3390/foods14071209