
A Quantitative Risk Assessment Model for Listeria monocytogenes in Ready-to-Eat Cantaloupe

 ,
,  , ,
, ,  , ,
, ,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Exposure Assessment
2.1.1. Preharvest Module
- Data and assumptions:

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Module | Stage | Microbial Process | Assumptions | Sources | Function in R |
|---|---|---|---|---|---|
| Preharvest | Soil and irrigation water contamination | Contamination | The function assumes cantaloupe contamination through soil and irrigation water, with soil and water contamination characteristics such as the prevalence (pSoil and pIrrig) and distributions of L. monocytogenes concentration in soil and water as inputs. Risk factors such as irrigation prior to harvest, the use of organic fertilizer, or the use of a soil barrier (mulch) are included. It makes use of the outputs of caIrrig2rind and caSoil2rind sub-functions (see below). | [38,53,54,55] | caPrimaryProduction |
| Irrigation water to rind contamination | Contamination | The function evaluates the contamination of cantaloupes through irrigation water only. It considers water contamination characteristics: prevalence (pIrrig) and concentration (cIrrig), which have to be chosen by the user based on existing data. cIrrig is conditional to water sources contaminated with L. monocytogenes. | [38,53,54,55] | caIrrig2rind | |
| Soil-to-rind contamination | Contamination | The function evaluates the contamination of cantaloupes through soil only. It considers soil contamination characteristics such as prevalence (pSoil) and concentration (cSoil) and the quantity of soil deposited on the cantaloupe rind. pSoil is conditional to risk factors such as irrigation before harvesting and the use of organic fertilizer, affecting pSoil with associated odds ratios (F_irrig_rain and fManure). It also takes into account the proportion of fields grown with a barrier (pFoil) and the reduction fraction of the quantity of soil transferred to the rind when a barrier is used (rFoil). | [18,56] | caSoil2rind | |
| Harvest | Contamination during harvest | Cross-contamination | The function simulates cross-contamination that might occur at the moment of harvesting from elements such as conveyors, crates, or plastic surfaces. It can be used for cantaloupes harvested in farms (intended for RTE and sold as whole cantaloupe in formal retail). Parameters such as the probability of cross-contamination (probCCH) and the variability in the transfer coefficient (trMean, trSd) help to assess the transfer. | caHarvestCC | |
| Holding time post-harvest | Survival | The function simulates the survival of L. monocytogenes on cantaloupe rind during post-harvest holding time or during any short storage before cantaloupes are washed in the packinghouse. It calculates the decline in L. monocytogenes, assuming no growth on surfaces because any injury to the rind that would promote growth would be recent and the holding time too short for significant growth. The function allows defining a probability that the lot is kept at cold temperatures (4–10 °C). | caHoldingTime | ||
| Pre-processing | Brushing cantaloupes | Removal | The function caBrush() models the removal of bacteria during the brushing or scrubbing step of cleaning cantaloupes. It requires the mean log10 reduction due to brushing as an input, which quantifies how effectively bacteria are removed from the rind during this cleaning step. | caBrush | |
| Processing | Flume tank cross-contamination | Cross-contamination | The caFlumeTankCC function simulates the potential contamination of cantaloupe when in direct contact with contaminated water in a flume tank. It accounts for four possible scenarios: cross-contamination in lots already contaminated, re-contamination in lots not previously contaminated, and scenarios where no cross-contamination occurs, regardless of initial lot status. | caFlumeTankCC | |
| Dicing of cantaloupe | Cross-contamination | The caDicing function simulates the transfer of L. monocytogenes from rind to flesh during the dicing of cantaloupe in a processing environment. It is assumed that each cantaloupe is separately diced, and if contaminated on the rind, a fraction of L. monocytogenes cells is transferred to the diced pieces. The function does not consider cross-contamination from dicing machines or knives. | [29,57] | caDicing | |
| Partitioning | Cross-contamination | The caPartitioningCC() function simulates the potential cross-contamination of cantaloupes during dicing and partitioning into packed units. It accounts for four possible scenarios of contamination involving sublots already contaminated or not. The algorithm also models the random distribution of L. monocytogenes from a contaminated sublot of diced cantaloupe into packed units using a dispersion factor, indicating the heterogeneity in the distribution of cells among pack units. | [58,59] | caPartitioningCC | |
| Microbiological lot testing | Microbiological testing of RTE cantaloupe | Removal | The caTesting() function simulates the microbiological testing of RTE cantaloupe samples from a lot or sublot. It models sampling and testing based on a defined sampling plan (two-class or three-class). The algorithm uses bootstrapping to estimate the probability of detecting contaminated lots and returns the output matrix either in the original lot or sublot arrangement, depending on the user’s choice. | caTesting | |
| Cold Chain Storage | All stages after processing | Growth | Bacterial growth is estimated using the primary growth model of with a lag phase Baranyi and Roberts [60], taking into account temperature conditions and the initial physiological state of cells (q0). | A range of research and studies, including [31,61], and several others, provide the data and parameters used to calibrate the growth model under temperatures below 30 °C. | caGrowthBaranyi |
| Transport from processing to retail | Growth | The caTrans2RetRTE() function simulates the growth of L. monocytogenes in RTE diced cantaloupe during cold transport to retail, utilizing the caGrowthBaranyi() function. It assumes uniform initial conditions for all RTE diced cantaloupe packs from each lot, including the same initial q0, transport temperature, and time. | [62,63] | caTrans2RetRTE | |
| Display of RTE diced cantaloupe packs at retail | Growth | The caRetRTE() function simulates the growth of L. monocytogenes in RTE diced cantaloupe during display at retail, using the caGrowthBaranyi() function. It assumes uniform retail conditions for all RTE diced cantaloupe packs, including the same lnQt (from the previous logistic stage), retail temperature, and sampled retail time. The Pert distributions represent the variability in retail time and temperature. | [64] | caRetRTE | |
| Transport of RTE diced cantaloupe packs from retail to home | Growth | The caRet2HomeRTE() function simulates the growth of L. monocytogenes in RTE diced cantaloupe during transport from retail to home, using the caGrowthBaranyi() function. The transportation time and temperature are sampled at the unit level to reflect the variability depending on the consumer. The algorithm uses a gamma distribution for the variability in transport time and a Pert distribution for transport temperature. | - | caRet2HomeRTE | |
| Consumer handling | Storage of RTE diced cantaloupe packs at home | Growth | The caHomeRTE() function simulates the growth of L. monocytogenes in RTE diced cantaloupe during home storage. It samples home storage time and temperature at the unit level, depending on consumer practices. The input data includes lot-specific values of EGR5 and unit-specific values of lnQt from the previous stage. Pert distributions are used to represent the variability in home storage time and temperature. | [62,65,66] | caHomeRTE |
| Country | Characteristics | Positive/Total (%Prevalence) | Source |
|---|---|---|---|
| Canada | Cultivated fields, 7 fields fertilized with animal manure in addition to inorganic fertilizer | 1/13 (7.7) | [38] |
| Malaysia | Vegetable fields in traditional farming | 4/21 (19.0) | [67] |
| USA | Organic/Irrigate/Manure/Compost Farm 1: no/no/yes/yes Farm 2: yes/yes/yes/yes Farm 3: no/yes/yes/no Farm 4: no/yes/no/no Farm 5: no/no/no/no (data broken down by farm not available) | 16/178 (8.9) | [55] |
| France | Cultivated soils from France | 9/53 (17.0) | [39] |
| Poland | Lands fertilized with manure Lands fertilized with artificial fertilizers Garden plots intensively fertilized with manure Wastelands | 2/173 (1.2) 0/173 (0.0) 5/47 (10.6) 0/120 (0.0) | [40] |
| Austria | Soil types (humus, sand, and clay) | 28/467 (6.0) | [68] |
| USA | Soil samples from spinach fields Low-risk fields High-risk fields | 24/546 (4.4) 62/546 (11.4) | [18] |
| Source | Risk Factor | Description | OR | 95% CI | p-Value |
|---|---|---|---|---|---|
| [55] | Manure | Last time manure was applied | |||
| Within 365 days | 7.0 | [3.1–15.4] | <0.001 | ||
| Over 365 days | 0.6 | [0.2–1.7] | 0.381 | ||
| Not applied | 1.0 | ||||
| Irrigation | Last time field was irrigated | ||||
| Within 3 days | 6.0 | [2.0–18.1] | 0.010 | ||
| 4–7 days | 1.2 | [0.3–4.5] | 0.793 | ||
| 8–14 days | 0.4 | [0.1–2.0] | 0.288 | ||
| Over 14 days/not irrigated | 1.0 | ||||
| Soil cultivation | Last time soil was cultivated | ||||
| Within 7 days | 2.9 | [1.1–8.6] | 0.050 | ||
| 8–14 days | 1.4 | [0.4–5.1] | 0.660 | ||
| 15–30 days | 0.4 | [0.1–1.7] | 0.224 | ||
| Over 30 days | 1.0 | ||||
| [18] | Irrigation/rain | Time since irrigation/rain occurred | |||
| 24 h | 25 | [5.7–99] | 0.010 | ||
| 48 h | 2.5 | [0.49–12] | 0.27 | ||
| 72 h | 3.4 | [0.74–15] | 0.11 | ||
| 144–192 h | 1.0 | ||||
| Amount of irrigation water (mm) applied to field 2 days before sample collection 1 | 1.2 | [1.1–1.3] | 0.010 | ||
| [48] | Areas within 37.5 m of surface water | 3.0 | [2.0–4.6] | <0.001 | |
| Areas within 62.5 m of pasture | 2.9 | [1.4–6.0] | 0.005 | ||
| Country | Type of Water | Positive/Total (% Prevalence) | Source |
|---|---|---|---|
| Austria | River and pond | 0/68 (0.0) | [68] |
| USA | Engineered water | 0/28 (0.0) | [56] |
| USA | Engineered water | 0/14 (0.0) | [55] |
| Malaysia | Irrigation water of vegetable farms | 0/15 (0.0) | [67] |
| India | River water | 8/100 (8.0) | [69] |
| Switzerland | River, stream, inland canal | 25/191 (13.1) | [53] |
| Canada | Rural and urban watersheds | 56/329 (17.0) | [70] |
| USA | Pond, river used for irrigation | 2/9 (22.2) | [56] |
| South Africa | Roof-harvested rain water | 72/297 (22.0) | [71] |
| Canada | Surface (river) | 32/134 (23.9) | [72] |
| USA | Surface water | 48/146 (33.0) | [55] |
| USA | Lake, stream, river, pond | 605/1405 (43.1) | [73] |
| South Africa | Irrigation canal and river | 19/36 (52.8) | [74] |
| USA | Surface water for irrigation | 33/52 (63.5) | [18] |
- The R functions:
2.1.2. Harvest of Cantaloupes
- Data and assumptions:
- The R functions:
2.1.3. Pre-Processing: Cleaning and Washing
- Data and assumptions:
- The R functions:
2.1.4. Processing
- Data and assumptions:
- The R function:
2.1.5. Microbiological Lot Testing
- Data and assumptions:
- The R function:
2.1.6. Cold Chain During Transport to Retail
- The R functions:
2.1.7. Consumer Handling
- Data and assumptions:
- The R function:
2.2. Risk Characterization
2.3. QRA Model’s Ouputs
2.4. QRA Model’s Implementation
3. Results and Discussion
3.1. Use of the Model for Risk Management Scenarios
3.2. Validation and Sensitivity Analysis
3.3. Perspectives and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chan, Y.-W.; Hoban, A.; Moore, H.; Greig, D.R.; Painset, A.; Jorgensen, F.; Chattaway, M.A.; Jenkins, C.; Balasegaram, S.; McCormick, J. Two outbreaks of foodborne gastrointestinal infection linked to consumption of imported melons, United Kingdom, March to August 2021. J. Food Prot. 2023, 86, 100027. [Google Scholar] [CrossRef]
- Jenkins, E.; Gardenhire, I.; Whitney, B.M.; Martin, K.B.; Schwensohn, C.; Gieraltowski, L.; Leeper, M.M.; McCurdy, V.; McClure, M.; Wellman, A. An investigation of an outbreak of Salmonella Newport infections linked to melons–United States, 2020. Food Control 2023, 152, 109833. [Google Scholar] [CrossRef]
- McGeoch, L.J.; Hoban, A.; Sawyer, C.; Rabie, H.; Painset, A.; Browning, L.; Brown, D.; McCarthy, C.; Nelson, A.; Firme, A. Salmonella Saintpaul outbreak associated with cantaloupe consumption, the United Kingdom and Portugal, September to November 2023. Epidemiol. Infect. 2024, 152, e78. [Google Scholar] [CrossRef]
- Federman, S.S.; Jenkins, E.; Wilson, C.; DeLaGarza, A.; Schwensohn, C.; Schneider, B.; Nsubuga, J.; Literman, R.; Wellman, A.; Whitney, B.M. An investigation of an outbreak of Salmonella Typhimurium infections linked to cantaloupe–United States, 2022. Food Control 2024, 166, 110733. [Google Scholar] [CrossRef]
- Walsh, K.A.; Bennett, S.D.; Mahovic, M.; Gould, L.H. Outbreaks associated with cantaloupe, watermelon, and honeydew in the United States, 1973–2011. Foodborne Pathog. Dis. 2014, 11, 945–952. [Google Scholar] [CrossRef]
- McCollum, J.T.; Cronquist, A.B.; Silk, B.J.; Jackson, K.A.; O’Connor, K.A.; Cosgrove, S.; Gossack, J.P.; Parachini, S.S.; Jain, N.S.; Ettestad, P. Multistate outbreak of listeriosis associated with cantaloupe. N. Engl. J. Med. 2013, 369, 944–953. [Google Scholar] [CrossRef]
- NSW. Listeria Outbreak Investigation. In Summary Report for the Melon Industry, October 2018; NSW Department of Primary Industries, Biosecurity & Food Safety: Orange, Australia, 2018; p. 12. ISBN 978-1-76058-267-8. [Google Scholar]
- EFSA Panel on Biological Hazards; Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernández Escámez, P.S.; Girones, R.; Herman, L.; Koutsoumanis, K. Listeria monocytogenes contamination of ready-to-eat foods and the risk for human health in the EU. EFSA J. 2018, 16, e05134. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, A.; Kooh, P.; Augustin, J.-C.; Guillier, L.; Thébault, A.; Cadavez, V.; Gonzales-Barron, U.; Sanaa, M. Risk factors for sporadic listeriosis: A systematic review and meta-analysis. Microb. Risk Anal. 2021, 17, 100128. [Google Scholar] [CrossRef]
- Locatelli, A.; Spor, A.; Jolivet, C.; Piveteau, P.; Hartmann, A. Biotic and abiotic soil properties influence survival of Listeria monocytogenes in soil. PLoS ONE 2013, 8, e75969. [Google Scholar] [CrossRef]
- Vivant, A.-L.; Garmyn, D.; Piveteau, P. Listeria monocytogenes, a down-to-earth pathogen. Front. Cell. Infect. Microbiol. 2013, 3, 87. [Google Scholar] [CrossRef] [PubMed]
- Kljujev, I.; Raicevic, V.; Jovicic-Petrovic, J.; Vujovic, B.; Mirkovic, M.; Rothballer, M. Listeria monocytogenes–Danger for health safety vegetable production. Microb. Pathog. 2018, 120, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Beuchat, L.R. Listeria monocytogenes: Incidence on vegetables. Food Control 1996, 7, 223–228. [Google Scholar] [CrossRef]
- Domenichini, G.; Fogliazza, D.; Pagani, M. Studies on the transmission of Listeria by means of arthropods. Ital. J. Food Sci. 1992, 4, 269–278. [Google Scholar]
- Lapanje, A.; Zrimec, A.; Drobne, D.; Rupnik, M. Long-term Hg pollution-induced structural shifts of bacterial community in the terrestrial isopod (Porcellio scaber) gut. Environ. Pollut. 2010, 158, 3186–3193. [Google Scholar] [CrossRef] [PubMed]
- Townsend, A.; Strawn, L.K.; Chapman, B.J.; Dunn, L.L. A systematic review of Listeria species and Listeria monocytogenes prevalence, persistence, and diversity throughout the fresh produce supply chain. Foods 2021, 10, 1427. [Google Scholar] [CrossRef]
- Lagarde, J.; Feurer, C.; Denis, M.; Douarre, P.-E.; Piveteau, P.; Roussel, S. Listeria monocytogenes prevalence and genomic diversity along the pig and pork production chain. Food Microbiol. 2023, 119, 104430. [Google Scholar] [CrossRef] [PubMed]
- Weller, D.; Wiedmann, M.; Strawn, L.K. Spatial and temporal factors associated with an increased prevalence of Listeria monocytogenes in spinach fields in New York State. Appl. Environ. Microbiol. 2015, 81, 6059–6069. [Google Scholar] [CrossRef]
- Mahmoud, B.; Vaidya, N.; Corvalan, C.; Linton, R. Inactivation kinetics of inoculated Escherichia coli O157: H7, Listeria monocytogenes and Salmonella Poona on whole cantaloupe by chlorine dioxide gas. Food Microbiol. 2008, 25, 857–865. [Google Scholar] [CrossRef]
- Ukuku, D.O.; Fett, W.F. Relationship of cell surface charge and hydrophobicity to strength of attachment of bacteria to cantaloupe rind. J. Food Prot. 2002, 65, 1093–1099. [Google Scholar] [CrossRef]
- Ferreira, V.; Wiedmann, M.; Teixeira, P.; Stasiewicz, M. Listeria monocytogenes persistence in food-associated environments: Epidemiology, strain characteristics, and implications for public health. J. Food Prot. 2014, 77, 150–170. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, B.; Cerf, O. Persistence of Listeria monocytogenes in food industry equipment and premises. Int. J. Food Microbiol. 2011, 145, 1–8. [Google Scholar] [CrossRef] [PubMed]
- U.S. FDA. Environmental Assessment: Factors Potentially Contributing to the Contamination of Fresh Whole Cantaloupe Implicated in a Multi-State Outbreak of Listeriosis. 2011. Available online: https://www.fda.gov/food/outbreaks-foodborne-illness/outbreak-investigation-reports (accessed on 8 June 2025).
- U.S. FDA. External Summary Report: FY 2013 Inspection, Environmental Sampling and Sample Collection (Pre and Post-Process) at Cantaloupe Packinghouses Assignment. 2015. Available online: https://www.fda.gov/food/foodborne-pathogens/external-summary-report-fy-2013-inspection-environmental-sampling-and-sample-collection-pre-and-post#_ftnref1 (accessed on 8 June 2025).
- Dunn, L.L.; Friedrich, L.M.; Strawn, L.K.; Danyluk, M.D. Prevalence of Listeria monocytogenes and indicator microorganisms in Florida cantaloupe packinghouses, 2013–2014. Food Microbiol. 2022, 104, 103970. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards; Koutsoumanis, K.; Ordóñez, A.A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; De Cesare, A.; Herman, L.; Hilbert, F.; Lindqvist, R.; et al. Microbiological hazards associated with the use of water in the post-harvest handling and processing operations of fresh and frozen fruits, vegetables and herbs (ffFVHs). Part 1 (outbreak data analysis, literature review and stakeholder questionnaire). EFSA J. 2023, 21, e08332. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.I.; Truchado, P.; Tudela, J.A.; Allende, A. Environmental monitoring of three fresh-cut processing facilities reveals harborage sites for Listeria monocytogenes. Food Control 2024, 155, 110093. [Google Scholar] [CrossRef]
- Sullivan, G.; Wiedmann, M. Detection and prevalence of Listeria in US produce packinghouses and fresh-cut facilities. J. Food Prot. 2020, 83, 1656–1666. [Google Scholar] [CrossRef]
- Ukuku, D.O.; Olanya, M.; Geveke, D.J.; Sommers, C.H. Effect of native microflora, waiting period, and storage temperature on Listeria monocytogenes serovars transferred from cantaloupe rind to fresh-cut pieces during preparation. J. Food Prot. 2012, 75, 1912–1919. [Google Scholar] [CrossRef]
- Bassett, J.; McClure, P. A risk assessment approach for fresh fruits. J. Appl. Microbiol. 2008, 104, 925–943. [Google Scholar] [CrossRef] [PubMed]
- Danyluk, M.D.; Friedrich, L.M.; Schaffner, D.W. Modeling the growth of Listeria monocytogenes on cut cantaloupe, honeydew and watermelon. Food Microbiol. 2014, 38, 52–55. [Google Scholar] [CrossRef]
- Li, D.; Friedrich, L.M.; Danyluk, M.D.; Harris, L.J.; Schaffner, D.W. Development and validation of a mathematical model for growth of pathogens in cut melons. J. Food Prot. 2013, 76, 953–958. [Google Scholar] [CrossRef]
- Gonzales-Barron, U.; Cadavez, V.; De Oliveira Mota, J.; Guillier, L.; Sanaa, M. A critical review of risk assessment models for Listeria monocytogenes in produce. Foods 2024, 13, 1111. [Google Scholar] [CrossRef]
- Guzel, M. Quantitative Risk Assessment for Listeria monocytogenes on Fresh-Cut Lettuce and Fresh-Cut Cantaloupe; Texas A&M University: College Station, TX, USA, 2015. [Google Scholar]
- FAO; WHO. Summary and Conclusions. In Proceedings of the Joint FAO/WHO Expert Meeting on Microbiological Risk Assessment of Listeria monocytogenes in Foods, Rome, Italy, 24–28 October 2022. [Google Scholar]
- FAO; WHO. Part 2: Risk Assessment Models. Summary and Conclusions. In Proceedings of the Joint FAO/WHO Expert Meeting on Microbiological Risk Assessment of Listeria monocytogenes in Foods, Rome, Italy, 29 May–2 June 2023. [Google Scholar]
- Nauta, M.J. Modelling bacterial growth in quantitative microbiological risk assessment: Is it possible? Int. J. Food Microbiol. 2002, 73, 297–304. [Google Scholar] [CrossRef]
- Dowe, M.J.; Jackson, E.D.; Mori, J.G.; Bell, C.R. Listeria monocytogenes survival in soil and incidence in agricultural soils. J. Food Prot. 1997, 60, 1201–1207. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, A.; Depret, G.; Jolivet, C.; Henry, S.; Dequiedt, S.; Piveteau, P.; Hartmann, A. Nation-wide study of the occurrence of Listeria monocytogenes in French soils using culture-based and molecular detection methods. J. Microbiol. Methods 2013, 93, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Szymczak, B.; Szymczak, M.; Sawicki, W.; Dąbrowski, W. Anthropogenic impact on the presence of L. monocytogenes in soil, fruits, and vegetables. Folia Microbiol. 2014, 59, 23–29. [Google Scholar] [CrossRef]
- Girardin, H.; Morris, C.E.; Albagnac, C.; Dreux, N.; Glaux, C.; Nguyen-The, C. Behaviour of the pathogen surrogates Listeria innocua and Clostridium sporogenes during production of parsley in fields fertilized with contaminated amendments. FEMS Microbiol. Ecol. 2005, 54, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Miceli, A.; Settanni, L. Influence of agronomic practices and pre-harvest conditions on the attachment and development of Listeria monocytogenes in vegetables. Ann. Microbiol. 2019, 69, 185–199. [Google Scholar] [CrossRef]
- Oliveira, M.; Usall, J.; Viñas, I.; Solsona, C.; Abadias, M. Transfer of Listeria innocua from contaminated compost and irrigation water to lettuce leaves. Food Microbiol. 2011, 28, 590–596. [Google Scholar] [CrossRef]
- McLaughlin, H.P.; Casey, P.G.; Cotter, J.; Gahan, C.G.; Hill, C. Factors affecting survival of Listeria monocytogenes and Listeria innocua in soil samples. Arch. Microbiol. 2011, 193, 775–785. [Google Scholar] [CrossRef]
- Nicholson, F.A.; Groves, S.J.; Chambers, B.J. Pathogen survival during livestock manure storage and following land application. Bioresour. Technol. 2005, 96, 135–143. [Google Scholar] [CrossRef]
- Allende, A.; Monaghan, J. Irrigation water quality for leafy crops: A perspective of risks and potential solutions. Int. J. Environ. Res. Public Health 2015, 12, 7457–7477. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on the risk posed by pathogens in food of non-animal origin. Part 2 (Salmonella and Norovirus in leafy greens eaten raw as salads). EFSA J. 2014, 12, 3600. [Google Scholar]
- Weller, D.; Shiwakoti, S.; Bergholz, P.; Grohn, Y.; Wiedmann, M.; Strawn, L.K. Validation of a previously developed geospatial model that predicts the prevalence of Listeria monocytogenes in New York State produce fields. Appl. Environ. Microbiol. 2016, 82, 797–807. [Google Scholar] [CrossRef]
- Nyarko, E.; Kniel, K.E.; Millner, P.D.; Luo, Y.; Handy, E.T.; Reynnells, R.; East, C.; Sharma, M. Survival and growth of Listeria monocytogenes on whole cantaloupes is dependent on site of contamination and storage temperature. Int. J. Food Microbiol. 2016, 234, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Handy, E.T.; East, C.L.; Kim, S.; Jiang, C.; Callahan, M.T.; Allard, S.M.; Micallef, S.; Craighead, S.; Anderson-Coughlin, B. Prevalence of Salmonella and Listeria monocytogenes in non-traditional irrigation waters in the Mid-Atlantic United States is affected by water type, season, and recovery method. PLoS ONE 2020, 15, e0229365. [Google Scholar] [CrossRef] [PubMed]
- Acheamfour, C.L.; Parveen, S.; Hashem, F.; Sharma, M.; Gerdes, M.E.; May, E.B.; Rogers, K.; Haymaker, J.; Duncan, R.; Foust, D. Levels of Salmonella enterica and Listeria monocytogenes in alternative irrigation water vary based on water source on the Eastern Shore of Maryland. Microbiol. Spectr. 2021, 9, e00669-21. [Google Scholar] [CrossRef]
- Iwu, C.D.; Iwu-Jaja, C.J.; Elhadi, R.; Semerjian, L.; Okoh, A.I. Modelling the potential risk of infection associated with Listeria monocytogenes in irrigation water and agricultural soil in two district municipalities in South Africa. Microorganisms 2022, 10, 181. [Google Scholar] [CrossRef] [PubMed]
- Raschle, S.; Stephan, R.; Stevens, M.J.; Cernela, N.; Zurfluh, K.; Muchaamba, F.; Nüesch-Inderbinen, M. Environmental dissemination of pathogenic Listeria monocytogenes in flowing surface waters in Switzerland. Sci. Rep. 2021, 11, 9066. [Google Scholar] [CrossRef]
- Richards, G.M.; Beuchat, L.R. Attachment of Salmonella Poona to cantaloupe rind and stem scar tissues as affected by temperature of fruit and inoculum. J. Food Prot. 2004, 67, 1359–1364. [Google Scholar] [CrossRef]
- Strawn, L.K.; Gröhn, Y.T.; Warchocki, S.; Worobo, R.W.; Bihn, E.A.; Wiedmann, M. Risk factors associated with Salmonella and Listeria monocytogenes contamination of produce fields. Appl. Environ. Microbiol. 2013, 79, 7618–7627. [Google Scholar] [CrossRef]
- Strawn, L.K.; Fortes, E.D.; Bihn, E.A.; Nightingale, K.K.; Gröhn, Y.T.; Worobo, R.W.; Wiedmann, M.; Bergholz, P.W. Landscape and meteorological factors affecting prevalence of three food-borne pathogens in fruit and vegetable farms. Appl. Environ. Microbiol. 2013, 79, 588–600. [Google Scholar] [CrossRef]
- Ukuku, D.O.; Fett, W. Behavior of Listeria monocytogenes inoculated on cantaloupe surfaces and efficacy of washing treatments to reduce transfer from rind to fresh-cut pieces. J. Food Prot. 2002, 65, 924–930. [Google Scholar] [CrossRef]
- Hoelzer, K.; Pouillot, R.; Gallagher, D.; Silverman, M.B.; Kause, J.; Dennis, S. Estimation of Listeria monocytogenes transfer coefficients and efficacy of bacterial removal through cleaning and sanitation. Int. J. Food Microbiol. 2012, 157, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Nauta, M.J. Microbiological risk assessment models for partitioning and mixing during food handling. Int. J. Food Microbiol. 2005, 100, 311–322. [Google Scholar] [CrossRef]
- Baranyi, J.; Roberts, T.A. A dynamic approach to predicting bacterial growth in food. Int. J. Food Microbiol. 1994, 23, 277–294. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Liu, Y.; Huang, L. Growth kinetics of Listeria monocytogenes and spoilage microorganisms in fresh-cut cantaloupe. Food Microbiol. 2013, 34, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Iwahori, J.i.; Kasuga, F.; Wang, J.; Forghani, F.; Park, M.-S.; Oh, D.-H. Risk assessment for Listeria monocytogenes on lettuce from farm to table in Korea. Food Control 2013, 30, 190–199. [Google Scholar] [CrossRef]
- Sant’Ana, A.S.; Franco, B.D.; Schaffner, D.W. Risk of infection with Salmonella and Listeria monocytogenes due to consumption of ready-to-eat leafy vegetables in Brazil. Food Control 2014, 42, 1–8. [Google Scholar] [CrossRef]
- Derens-Bertheau, E.; Osswald, V.; Laguerre, O.; Alvarez, G. Cold chain of chilled food in France. Int. J. Refrig. 2015, 52, 161–167. [Google Scholar] [CrossRef]
- Carrasco, E.; Pérez-Rodríguez, F.; Valero, A.; García-Gimeno, R.M.; Zurera, G. Risk assessment and management of Listeria monocytogenes in ready-to-eat lettuce salads. Compr. Rev. Food Sci. Food Saf. 2010, 9, 498–512. [Google Scholar] [CrossRef]
- Nauta, M.J.; Litman, S.; Barker, G.C.; Carlin, F. A retail and consumer phase model for exposure assessment of Bacillus cereus. Int. J. Food Microbiol. 2003, 83, 205–218. [Google Scholar] [CrossRef]
- Jeyaletchumi, P.; Tunung, R.; Selina, P.M.; Chai, L.; Radu, S.; Farinazleen, M.; Cheah, Y.; Mitsuaki, N.; Yoshitsugu, N.; Kumar, M.P. Evaluation of Listeria spp. and Listeria monocytogenes in selected vegetable farms. J. Trop. Agric. Food Sci. 2011, 39, 255–266. [Google Scholar]
- Linke, K.; Rückerl, I.; Brugger, K.; Karpiskova, R.; Walland, J.; Muri-Klinger, S.; Tichy, A.; Wagner, M.; Stessl, B. Reservoirs of Listeria species in three environmental ecosystems. Appl. Environ. Microbiol. 2014, 80, 5583–5592. [Google Scholar] [CrossRef]
- Soni, D.K.; Singh, R.K.; Singh, D.V.; Dubey, S.K. Characterization of Listeria monocytogenes isolated from Ganges water, human clinical and milk samples at Varanasi, India. Infect. Genet. Evol. 2013, 14, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Stea, E.C.; Purdue, L.M.; Jamieson, R.C.; Yost, C.K.; Truelstrup Hansen, L. Comparison of the prevalences and diversities of Listeria species and Listeria monocytogenes in an urban and a rural agricultural watershed. Appl. Environ. Microbiol. 2015, 81, 3812–3822. [Google Scholar] [CrossRef] [PubMed]
- Jongman, M.; Korsten, L. Assessment of irrigation water quality and microbiological safety of leafy greens in different production systems. J. Food Saf. 2017, 37, e12324. [Google Scholar] [CrossRef]
- Lyautey, E.; Lapen, D.R.; Wilkes, G.; McCleary, K.; Pagotto, F.; Tyler, K.; Hartmann, A.; Piveteau, P.; Rieu, A.; Robertson, W.J. Distribution and characteristics of Listeria monocytogenes isolates from surface waters of the South Nation River watershed, Ontario, Canada. Appl. Environ. Microbiol. 2007, 73, 5401–5410. [Google Scholar] [CrossRef]
- Cooley, M.B.; Quiñones, B.; Oryang, D.; Mandrell, R.E.; Gorski, L. Prevalence of shiga toxin producing Escherichia coli, Salmonella enterica, and Listeria monocytogenes at public access watershed sites in a California Central Coast agricultural region. Front. Cell. Infect. Microbiol. 2014, 4, 30. [Google Scholar] [CrossRef]
- Ijabadeniyi, O.A.; Debusho, L.K.; Vanderlinde, M.; Buys, E.M. Irrigation water as a potential preharvest source of bacterial contamination of vegetables. J. Food Saf. 2011, 31, 452–461. [Google Scholar] [CrossRef]
- Fu, Y.; Bhunia, A.K.; Yao, Y. Abrasive brushing reduces pathogen biofilms at cantaloupe rind surface. Int. J. Food Microbiol. 2020, 329, 108685. [Google Scholar] [CrossRef]
- Guzel, M.; Moreira, R.G.; Omac, B.; Castell-Perez, M.E. Quantifying the effectiveness of washing treatments on the microbial quality of fresh-cut romaine lettuce and cantaloupe. LWT 2017, 86, 270–276. [Google Scholar] [CrossRef]
- Ukuku, D.O.; Bari, M.; Kawamoto, S.; Isshiki, K. Use of hydrogen peroxide in combination with nisin, sodium lactate and citric acid for reducing transfer of bacterial pathogens from whole melon surfaces to fresh-cut pieces. Int. J. Food Microbiol. 2005, 104, 225–233. [Google Scholar] [CrossRef] [PubMed]
- FAO; WHO. Prevention and Control of Microbiological Hazards in Fresh Fruits and Vegetables: Part 4: Specific Commodities: Meeting Report; FAO: Rome, Italy; WHO: Geneva, Switzerland, 2023. [Google Scholar] [CrossRef]
- FDA. FDA-iRISK 4.2 Food Safety Modeling Tool: Technical Document; U.S. Food and Drug Administration: Silver Spring, MD, USA, 2021. Available online: https://irisk.foodrisk.org/Documents/FDAiRISKTechnicalDocumentation.pdf (accessed on 8 June 2025).
- Selma, M.V.; Ibáñez, A.M.; Allende, A.; Cantwell, M.; Suslow, T. Effect of gaseous ozone and hot water on microbial and sensory quality of cantaloupe and potential transference of Escherichia coli O157: H7 during cutting. Food Microbiol. 2008, 25, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Patil, R.D. Transfer of Listeria monocytogenes During Cutting, Slicing, Dicing, and Subsequent Storage of Cantaloupe and Honeydew Melons. Master’s Thesis, Michigan State University, East Lansing, MI, USA, 2017. [Google Scholar]
- Pouillot, R.; Chen, Y.; Van Doren, J.M. Elucidating the influence of the lower and upper microbiological limits: When is a 3-class sampling plan useful to test for pathogens in food? Food Control 2024, 163, 110544. [Google Scholar] [CrossRef]
- Mercier, S.; Villeneuve, S.; Mondor, M.; Uysal, I. Time–temperature management along the food cold chain: A review of recent developments. Compr. Rev. Food Sci. Food Saf. 2017, 16, 647–667. [Google Scholar] [CrossRef] [PubMed]
- McKellar, R.C.; LeBlanc, D.I.; Lu, J.; Delaquis, P. Simulation of Escherichia coli O157: H7 behavior in fresh-cut lettuce under dynamic temperature conditions during distribution from processing to retail. Foodborne Pathog. Dis. 2012, 9, 239–244. [Google Scholar] [CrossRef]
- Scolforo, C.Z.; Bairros, J.V.; Rezende, A.C.B.; Silva, B.S.; Alves, R.B.; Costa, D.S.; Andrade, N.J.; Sant’Ana, A.S.; Pena, W.E. Modeling the fate of Listeria monocytogenes and Salmonella enterica in the pulp and on the outer rind of Canary melons (Cucumis melo (Indorus Group)). LWT 2017, 77, 290–297. [Google Scholar] [CrossRef]
- Farber, J.; Wang, S.; Cai, Y.; Zhang, S. Changes in populations of Listeria monocytogenes inoculated on packaged fresh-cut vegetables. J. Food Prot. 1998, 61, 192–195. [Google Scholar] [CrossRef]
- Hong, Y.K.; Yoon, W.B.; Huang, L.; Yuk, H.G. Predictive Modeling for Growth of Non-and Cold-adapted Listeria monocytogenes on Fresh-cut Cantaloupe at Different Storage Temperatures. J. Food Sci. 2014, 79, M1168–M1174. [Google Scholar] [CrossRef]
- Moreira, J.; Mera, E.; Singh Chhetri, V.; King, J.M.; Gentimis, T.; Adhikari, A. Effect of storage temperature and produce type on the survival or growth of Listeria monocytogenes on peeled rinds and fresh-cut produce. Front. Microbiol. 2023, 14, 1151819. [Google Scholar] [CrossRef]
- FSA. National Microbiological Survey and Consumer Habits in relation to Frozen Vegetables, Fruits and Herbs (19NS6); Monitoring & Surveillance Series; Food Safety Authority of Ireland, safeFood: Dublin, Ireland, 2022; p. 62. ISBN 978-1-910348-54-3. [Google Scholar]
- Pouillot, R.; Miconnet, N.; Afchain, A.-L.; Delignette-Muller, M.L.; Beaufort, A.; Rosso, L.; Denis, J.-B.; Cornu, M. Quantitative Risk Assessment of Listeria monocytogenes in French Cold-Smoked Salmon: I. Quantitative Exposure Assessment. Risk Anal. 2007, 27, 683–700. [Google Scholar] [CrossRef]
- O’Connor-Shaw, R.; Roberts, R.; Ford, A.; Nottingham, S. Shelf life of minimally processed honeydew, kiwifruit, papaya, pineapple and cantaloupe. J. Food Sci. 1994, 59, 1202–1206. [Google Scholar] [CrossRef]
- Bonanno, L.; Bergis, H.; Gnanou-Besse, N.; Asséré, A.; Danan, C. Which domestic refrigerator temperatures in Europe?-Focus on shelf-life studies regarding Listeria monocytogenes (Lm) in ready-to-eat (RTE) foods. Food Microbiol. 2024, 123, 104595. [Google Scholar] [CrossRef] [PubMed]
- Roccato, A.; Uyttendaele, M.; Membré, J.-M. Analysis of domestic refrigerator temperatures and home storage time distributions for shelf-life studies and food safety risk assessment. Food Res. Int. 2017, 96, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.; Celidonio, F.; Oliveira, K. Verification of the temperature of domestic refrigerators to minimize the deterioration and possible foodborne illnesses. Hig. Aliment. 2008, 22, 42–45. [Google Scholar]
- Amaro, A.L.; Spadafora, N.D.; Pereira, M.J.; Dhorajiwala, R.; Herbert, R.J.; Müller, C.T.; Rogers, H.J.; Pintado, M. Multitrait analysis of fresh-cut cantaloupe melon enables discrimination between storage times and temperatures and identifies potential markers for quality assessments. Food Chem. 2018, 241, 222–231. [Google Scholar] [CrossRef]
- Luciano, W.A.; Griffin, S.; Targino de Souza Pedrosa, G.; Alvarenga, V.; Valdramidis, V.; Magnani, M. Growth behavior of low populations of Listeria monocytogenes on fresh-cut mango, melon and papaya under different storage temperatures. Food Microbiol. 2022, 102, 103930. [Google Scholar] [CrossRef]
- Pouillot, R.; Kiermeier, A.; Guillier, L.; Cadavez, V.; Sanaa, M. Updated Parameters for Listeria monocytogenes Dose–Response Model Considering Pathogen Virulence and Age and Sex of Consumer. Foods 2024, 13, 751. [Google Scholar] [CrossRef]
- FAO; WHO. Risk Assessment of Listeria monocytogenes in Ready-to-Eat Foods: Technical Report; World Health Organization and Food and Agriculture Organization of the United Nations: Geneve, Switzerland, 2004; pp. 1–269. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 8 June 2025.).
- Johnston, L.M.; Jaykus, L.-A.; Moll, D.; Martinez, M.C.; Anciso, J.; Mora, B.; Moe, C.L. A field study of the microbiological quality of fresh produce. J. Food Prot. 2005, 68, 1840–1847. [Google Scholar] [CrossRef]
- Zhang, H.; Yamamoto, E.; Murphy, J.; Locas, A. Microbiological safety of ready-to-eat fresh-cut fruits and vegetables sold on the Canadian retail market. Int. J. Food Microbiol. 2020, 335, 108855. [Google Scholar] [CrossRef]
- Félix, B.; Sevellec, Y.; Palma, F.; Douarre, P.E.; Felten, A.; Radomski, N.; Mallet, L.; Blanchard, Y.; Leroux, A.; Soumet, C.; et al. A European-wide dataset to uncover adaptive traits of Listeria monocytogenes to diverse ecological niches. Sci. Data 2022, 9, 190. [Google Scholar] [CrossRef]
- Jorgensen, J.; Bland, R.; Waite-Cusic, J.; Kovacevic, J. Diversity and antimicrobial resistance of Listeria spp. and L. monocytogenes clones from produce handling and processing facilities in the Pacific Northwest. Food Control 2021, 123, 107665. [Google Scholar] [CrossRef]
- Sullivan, G.; Orsi, R.H.; Estrada, E.; Strawn, L.; Wiedmann, M. Whole-genome sequencing-based characterization of Listeria isolates from produce packinghouses and fresh-cut facilities suggests both persistence and reintroduction of fully virulent L. monocytogenes. Appl. Environ. Microbiol. 2022, 88, e01177-22. [Google Scholar] [CrossRef] [PubMed]





| Source | Inoculation | Temp. (°C) | Time (Day) | Counts |
|---|---|---|---|---|
| Spot-inoculated (6 log10) on drawn circles (11 cm2) of cantaloupe, then allowed to dry for 1 h. | log10 CFU/cantaloupe (Athena/Rocky Ford Cultivars) | |||
| 4 | 0 | 5.77/5.77 | ||
| 1 | 4.96/5.57 | |||
| 3 | 3.65/4.35 | |||
| 5 | 2.78/3.48 | |||
| 7 | 2.87/3.39 | |||
| 9 | 0.78/2.00 | |||
| 15 | 1.75/2.96 | |||
| 10 | 0 | 5.80/5.80 | ||
| 1 | 5.59/5.52 | |||
| 3 | 4.83/4.35 | |||
| 5 | 4.14/4.62 | |||
| 7 | 3.73/4.28 | |||
| 9 | 2.76/4.00 | |||
| 25 | 0 | 5.85/5.85 | ||
| 1 | 5.10/5.10 | |||
| 3 | 5.10/2.03 | |||
| 5 | 3.30/2.10 | |||
| 7 | 3.23/3.23 | |||
| Inoculated by immersion in 3 L suspension at 8 log10 CFU/mL, then allowed to dry for 1 h. | log10 CFU/cm2 | |||
| 4 | 0 | 3.47 | ||
| 1 | 3.47 | |||
| 3 | 3.08 | |||
| 6 | 2.93 | |||
| 9 | 2.77 | |||
| 15 | 2.46 | |||
| 20 | 0 | 3.47 | ||
| 1 | 3.47 | |||
| 3 | 3.08 | |||
| 6 | 2.70 | |||
| 9 | 2.31 | |||
| 15 | 1.50 |
| Treatment | Study | Concentration (%) | Exposure Time (min) | Temperature (°C) | Reduction (log10 CFU/cm2) | St. Error (Reduction) | n 1 |
|---|---|---|---|---|---|---|---|
| Tap | [76] | - | 3 | 25 | 0.18 | 0.053 | 8 |
| water | [29] | 5 | 25 | 0.20 | - | 3 | |
| [77] 2 | |||||||
| Stored at 5 °C x 0 day | 5 | 20 | 0.20 | - | 3 | ||
| Stored at 5 °C x 7 days | 5 | 20 | 0.15 | 3 | |||
| [57] 2 | |||||||
| Stored at 4 °C x 1 day | - | 2 | 25 | 0.30 | - | 3 | |
| Stored at 4 °C x 5 days | - | 2 | 25 | 0.22 | - | 3 | |
| Stored at 4 °C x 15 days | - | 2 | 25 | 0.22 | - | 3 | |
| ClO2 gas | [19] | 0.00005 | 2 | 25 | 1.2 | - | 6 |
| 0.0001 | 2 | 25 | 1.8 | - | 6 | ||
| 0.00015 | 2 | 25 | 2.1 | - | 6 | ||
| 0.0003 | 2 | 25 | 2.1 | - | 6 | ||
| 0.0005 | 2 | 25 | 2.2 | - | 6 | ||
| 0.00005 | 10 | 25 | 3.3 | - | 6 | ||
| 0.0001 | 10 | 25 | 3.2 | - | 6 | ||
| 0.00015 | 10 | 25 | 3.7 | 6 | |||
| 0.0003 | 10 | 25 | 3.8 | - | 6 | ||
| 0.0005 | 10 | 25 | 4.3 | - | 6 | ||
| SH 3 | 0.020 | 5 | 25 | 0.57 | 0.250 | 8 | |
| Stored at 4 °C x 1 day | 0.100 | 2 | 25 | >3.0 | - | 3 | |
| Stored at 4 °C x 5 days | 0.100 | 2 | 25 | >3.0 | - | 3 | |
| Stored at 4 °C x 15 days | 0.100 | 2 | 25 | >3.0 | - | 3 | |
| H2O2 | [29] | 2.5 | 5 | 25 | 2.8 | 3 | |
| [77] 2 | |||||||
| Stored at 5 °C x 0 day | 2.5 | 5 | 20 | 2.3 | - | 3 | |
| Stored at 5 °C x 7 days | 2.5 | 5 | 20 | 2.8 | 3 | ||
| [57] 2 | |||||||
| Stored at 4 °C x 1 day | 5.0 | 2 | 25 | >3.0 | - | 3 | |
| Stored at 4 °C x 5 days | 5.0 | 2 | 25 | >3.0 | - | 3 | |
| Stored at 4 °C x 15 days | 5.0 | 2 | 25 | >3.0 | - | 3 | |
| HPLNC 4 | [77] 2 | ||||||
| Stored at 5 °C x 0 day | - | 5 | 20 | >4.0 | - | 3 | |
| Stored at 5 °C x 7 days | - | 5 | 20 | >4.0 | - | 3 |
| Source | L. monocytogenes on the Rind (log10 CFU/cm2) | L. monocytogenes in Cantaloupe Flesh | Transfer Rate (%) (10flesh/10rind) × 100 | |
|---|---|---|---|---|
| To fresh-cut (diced pieces) | (log10 CFU/g) | |||
| [20] | 2.16 | 0.23 | 1.175 | |
| 3.26 | 0.54 | 0.191 | ||
| 3.98 | 1.31 | 0.214 | ||
| 4.52 | 1.46 | 0.087 | ||
| [29] | 4.60 | 2.60 | 1.000 | |
| 4.40 | 2.20 | 0.631 | ||
| To slices (flesh surface) | (log10 CFU/cm2) | |||
| [81] | 4 °C | 5.94 | 2.45 | 0.032 |
| 5.44 | 1.42 | 0.010 | ||
| 30 °C | 5.22 | 1.64 | 0.026 | |
| 5.44 | 1.17 | 0.005 | ||
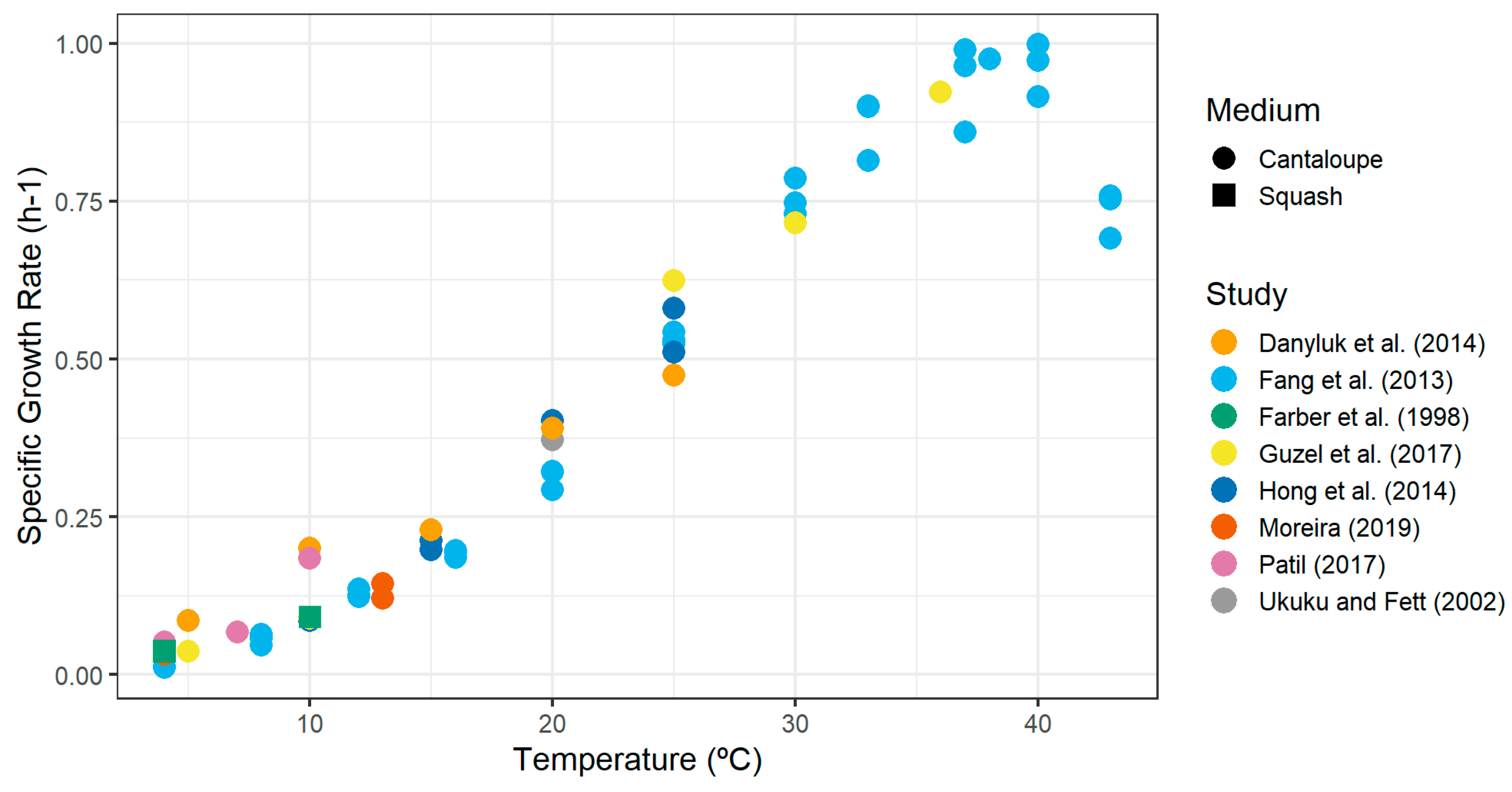
| Study | Medium | Strain | Stressed | Temperature (°C) | Specific GR (h−1) |
|---|---|---|---|---|---|
| [61] | Cantaloupe | F2365, H7858, ATCC19115 | Stressed | 4.0 | 0.0120 |
| (rifampicin-resistant and cold-resistant) | 8.0 | 0.0470 | |||
| 12 | 0.1260 | ||||
| 16 | 0.1860 | ||||
| 20 | 0.2930 | ||||
| 25 | 0.5250 | ||||
| 30 | 0.7300 | ||||
| 33 | 0.8150 | ||||
| 40 | 0.9160 | ||||
| 37 | 0.8600 | ||||
| 43 | 0.6920 | ||||
| F4260 | Stressed (rifampicin-resistant and cold-resistant) | 4.0 | 0.0110 | ||
| 8.0 | 0.0580 | ||||
| 12 | 0.1230 | ||||
| 16 | 0.1940 | ||||
| 20 | 0.3210 | ||||
| 25 | 0.5300 | ||||
| 30 | 0.7470 | ||||
| 33 | 0.9000 | ||||
| 37 | 0.9900 | ||||
| 40 | 0.9730 | ||||
| 43 | 0.7590 | ||||
| 38 | 0.9750 | ||||
| V7 | Stressed (rifampicin-resistant and cold-resistant) | 43 | 0.7540 | ||
| 40 | 0.9980 | ||||
| 33 | 0.9010 | ||||
| 37 | 0.9640 | ||||
| 30 | 0.7860 | ||||
| 25 | 0.5430 | ||||
| 20 | 0.3220 | ||||
| 16 | 0.1970 | ||||
| 12 | 0.1350 | ||||
| 8.0 | 0.0640 | ||||
| [87] | Cantaloupe | ATCC BAA839, ATCC BAA839, ATCC 19111, ATCC 13932 | Stressed (rifampicin-resistant and cold-resistant) | 10 | 0.0852 |
| 15 | 0.1983 | ||||
| 20 | 0.4030 | ||||
| 25 | 0.5803 | ||||
| 10 | 0.0852 | ||||
| 15 | 0.2118 | ||||
| 20 | 0.4030 | ||||
| 25 | 0.5112 | ||||
| Diced cantaloupe | Scott A, H7778, ATCC-15313, CCR1LG | Not stressed | 20 | 0.3720 | |
| Fresh-cut cantaloupe | LCDC 81-861, Scott A, 101M, V7 | Not stressed | 5.0 | 0.0850 | |
| 10 | 0.2000 | ||||
| 15 | 0.2300 | ||||
| 20 | 0.3900 | ||||
| 25 | 0.4745 | ||||
| Fresh-cut cantaloupe | NRCC B33076 | Not stressed | 5.0 | 0.0368 | |
| 10 | 0.0898 | ||||
| 25 | 0.6240 | ||||
| 30 | 0.7161 | ||||
| 36 | 0.9233 | ||||
| [81] | Diced cantaloupe | J22F, J29H, M3 | Not stressed | 4.0 | 0.0520 |
| 7.0 | 0.0670 | ||||
| 10 | 0.1840 | ||||
| [88] | Fresh-cut watermelon | LCDC 81-861, V7, 101M, Scott A | Not stressed | 4.0 | 0.0318 |
| 13 | 0.1213 | ||||
| 13 | 0.1438 | ||||
| [86] | Squash | Not stated | Not stressed | 4.0 | 0.0370 |
| 10 | 0.0910 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guillier, L.; Gonzales-Barron, U.; Pouillot, R.; De Oliveira Mota, J.; Allende, A.; Kovacevic, J.; Guldimann, C.; Fazil, A.; Al-Qadiri, H.; Dong, Q.; et al. A Quantitative Risk Assessment Model for Listeria monocytogenes in Ready-to-Eat Cantaloupe. Foods 2025, 14, 2212. https://doi.org/10.3390/foods14132212
Guillier L, Gonzales-Barron U, Pouillot R, De Oliveira Mota J, Allende A, Kovacevic J, Guldimann C, Fazil A, Al-Qadiri H, Dong Q, et al. A Quantitative Risk Assessment Model for Listeria monocytogenes in Ready-to-Eat Cantaloupe. Foods. 2025; 14(13):2212. https://doi.org/10.3390/foods14132212
Chicago/Turabian StyleGuillier, Laurent, Ursula Gonzales-Barron, Régis Pouillot, Juliana De Oliveira Mota, Ana Allende, Jovana Kovacevic, Claudia Guldimann, Aamir Fazil, Hamzah Al-Qadiri, Qingli Dong, and et al. 2025. "A Quantitative Risk Assessment Model for Listeria monocytogenes in Ready-to-Eat Cantaloupe" Foods 14, no. 13: 2212. https://doi.org/10.3390/foods14132212
APA StyleGuillier, L., Gonzales-Barron, U., Pouillot, R., De Oliveira Mota, J., Allende, A., Kovacevic, J., Guldimann, C., Fazil, A., Al-Qadiri, H., Dong, Q., Hasegawa, A., Cadavez, V., & Sanaa, M. (2025). A Quantitative Risk Assessment Model for Listeria monocytogenes in Ready-to-Eat Cantaloupe. Foods, 14(13), 2212. https://doi.org/10.3390/foods14132212