Abstract
We analyzed antimicrobial resistance and virulence traits in multidrug-resistant (MDR) E. coli isolates obtained from imported shrimp using whole-genome sequences (WGSs). Antibiotic resistance profiles were determined phenotypically. WGSs identified key characteristics, including their multilocus sequence type (MLST), serotype, virulence factors, antibiotic resistance genes, and mobile elements. Most of the isolates exhibited resistance to gentamicin, streptomycin, ampicillin, chloramphenicol, nalidixic acid, ciprofloxacin, tetracycline, and trimethoprim/sulfamethoxazole. Multilocus sequence type (MLST), serotype, average nucleotide identity (ANI), and pangenome analysis showed high genomic similarity among isolates, except for EC15 and ECV01. The EC119 plasmid contained a variety of efflux pump genes, including those encoding the acid resistance transcriptional activators (gadE, gadW, and gadX), resistance-nodulation-division-type efflux pumps (mdtE and mdtF), and a metabolite, H1 symporter (MHS) family major facilitator superfamily transporter (MNZ41_23075). Virulence genes displayed diversity, particularly EC15, whose plasmids carried genes for adherence (faeA and faeC-I), invasion (ipaH and virB), and capsule (caf1A and caf1M). This comprehensive analysis illuminates antimicrobial resistance, virulence, and plasmid dynamics in E. coli from imported shrimp and has profound implications for public health, emphasizing the need for continued surveillance and research into the evolution of these important bacterial pathogens.
1. Introduction
The average U.S. seafood consumption in 2021 was 20.5 pounds per person, according to the National Fisheries Institute. Shrimp was the most popular choice, with individuals consuming 5.90 pounds per person [1]. However, much shrimp is imported from outside the U.S. In 2020, the U.S. imported approximately 6 billion pounds of seafood products, with shrimp comprising 27% of the total value of edible imports [2]. Although the Food and Drug Administration (FDA) has not approved the use of any antibiotics in U.S. shrimp aquaculture, other nations use various antibiotics in their shrimp aquaculture practices [3]. Multiple studies have demonstrated the presence of antimicrobial residues, including β-lactam, erythromycin, sulfonamide, and tetracycline, in samples of shrimp sourced from aquaculture operations in Southeast Asia [4,5].
Furthermore, several studies have indicated that shrimp is a carrier of antibiotic-resistant bacteria [6,7]. Antibiotic resistance genes present in multidrug-resistant (MDR) bacteria within shrimp can disseminate through horizontal gene transfer, potentially reaching pathogenic bacteria [8]. If a person becomes infected with these pathogenic bacteria, antibiotic treatments could be less effective. Consuming shrimp contaminated with MDR bacteria may facilitate the transfer of antibiotic resistance determinants between MDR and commensal bacteria. Elevated transfer rates of antibiotic resistance may potentially give rise to new antibiotic-resistant bacteria through the exchange of resistant genes within the human gut. This could disrupt the balance of human intestinal microbiota, posing a potential health risk [9]. Our objective in this study was to analyze antimicrobial resistance and virulence traits in MDR E. coli isolates from imported shrimp using whole-genome sequences (WGSs).
2. Materials and Methods
2.1. Isolation of E. coli from Imported Shrimp
A total of 330 frozen, imported shrimp were thawed and incubated with Luria broth (Thermo Fisher Scientific, Waltham, MA, USA) overnight. Next, the sample was streaked onto a MacConkey agar plate (Thermo Fisher Scientific), and the identity of an E. coli colony was confirmed using the Vitek GNI+ card (bioMerieux, Durham, NC, USA) and fatty acid methyl ester analysis (MIDI, Newark, DE, USA).
2.2. Phenotypic Analysis of Antibiotic Resistance
We employed the disk diffusion assay to assess the phenotypic antibiotic susceptibility of E. coli isolates [10]. It included antibiotic disks from diverse classes, such as aminoglycosides, beta-lactams, chloramphenicol, quinolones, fosfomycin, tetracycline, trimethoprim/sulfamethoxazole, and polymyxin B. Antimicrobial susceptibilities were assessed based on the criteria established by the Clinical and Laboratory Standards Institute [11].
2.3. Whole-Genome Sequencing
Genomic DNA were extracted from an overnight culture of 14 MDR E. coli using the DNeasy Blood and Tissue Kit (Qiagen, Valencia, CA, USA). DNA were fragmented using g-TUBE (Covaris, Woburn, MA, USA) to approximately 6.0 kb. We constructed libraries using the SMRTbell Express Template Prep Kit v2.0 (Pacific Biosciences, Menlo Park, CA, USA). SMRT cells of the libraries were sequenced using the PacBio Sequel II platform (Pacific Biosciences). Long-read sequencing data were assembled with the SMRT Link v10.0 Microbial Assembly application (Pacific Biosciences). Detailed information of the WGSs was reported by Alotaibi et al. [12] and sequencing data are available under the National Center for Biotechnology Information’s (NCBI) BioProject number PRJNA802087.
2.4. Identification of MLST, Serotype, Virulence, Antibiotic Resistance, and Mobile Elements
We determined multilocus sequence types (MLSTs) and serotypes of the assembled genomes by using MLST v.2.0. (https://cge.food.dtu.dk/services/MLST/ (accessed on 24 November 2023)), cgMLST v.1.2. (https://cge.food.dtu.dk/services/cgMLSTFinder/ (accessed on 5 May 2024)), and SerotypeFinder v.2.0 (https://cge.food.dtu.dk/services/SerotypeFinder/ (accessed on 25 November 2023)), available on the Center for Genomic Epidemiology (CGE) server [13]. Average nucleotide identity (ANI) values were determined by using Integrated Prokaryotes Genome and pangenome Analysis (IPGA) v.1.09 (https://nmdc.cn/ipga/ (accessed on 13 November 2023)) [14]. Antimicrobial resistance genes and point mutations were identified by using multiple databases, including the Comprehensive Antibiotic Resistance Database (CARD), the Bacterial and Viral Bioinformatics Resource Center (BV-BRC) (https://www.bv-brc.org/ (accessed on 16 January 2024)), the Bacterial Antimicrobial Resistance Reference Gene Database hosted by the NCBI as part of the National Database of Antibiotic Resistant Organisms (NDARO) (https://www.ncbi.nlm.nih.gov/pathogens/isolates/ (accessed on 20 October 2023)), and ResFinderPlus v.4.5.0. (http://genepi.food.dtu.dk/resfinder (accessed on 1 November 2023)) [15,16,17]. Mobile elements were identified by using Mobile Element Finder v.1.0.3 (https://cge.food.dtu.dk/services/MobileElementFinder/ (accessed on 27 November 2023)) on the CGE and mobileOG-db v1.1.3 of Proksee (https://proksee.ca/ (accessed on 28 November 2023)) [18,19]. Plasmid mobility and plasmid replicon were identified by using VRprofile2 (https://tool2-mml.sjtu.edu.cn/VRprofile/ (accessed on 3 December 2023)) [20]. Virulence factors were identified based on the datasets of the Virulence Factors of Pathogenic Bacteria Database (VFDB), the PATRIC curated virulence database (PATRIC_VF), and the Victors database [21,22,23].
2.5. Pangenome Analysis
We conducted pangenome analysis using CGView (https://server.gview.ca/ (accessed on 13 October 2023)) [24]. On the CGView server, Basic Local Alignment Search Tool (BLAST) analysis was performed using GenBank files of plasmid sequences, with an E value < 1 × 10−10, an alignment length cutoff value of 100, and a percent identity cutoff value of 80. The schematic maps of the complete genome structure of the plasmids were generated by using CGView.
3. Results
3.1. Antibiotic Resistance Profiles
Antibiotic resistance profiles of different E. coli isolates revealed a diverse landscape of susceptibility and resistance patterns across various antimicrobial agents (Figure 1). Notably, there was variability in resistance to a range of antibiotics among the isolates. For instance, isolates EC0002, EC110, EC119, EC120, EC123, EC126, EC331, EC335, EC338, EC339, EC1110, and EC2110 consistently exhibited resistance to gentamicin, streptomycin, ampicillin, chloramphenicol, ciprofloxacin, nalidixic acid, tetracycline, and trimethoprim/sulfamethoxazole. However, resistance to kanamycin and cefepime varied across these isolates, with some showing susceptibility or intermediate resistance.
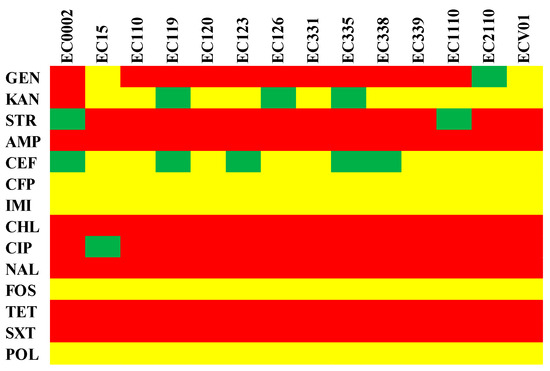
Figure 1.
Phenotypic antibiotic resistance profile of E. coli isolates. GEN: gentamicin, KAN: kanamycin, STR: streptomycin, AMP: ampicillin, CEF: cefepime, CFP: cefoperazone, IMI: imipenem, CHL: chloramphenicol, CIP: ciprofloxacin, NAL: nalidixic acid, FOS: fosfomycin, TET: tetracycline, SXT: trimethoprim/sulfamethoxazole, and POL: polymyxin B. The figure shows antimicrobial susceptibility as resistant (red), intermediate (green), or susceptible (yellow).
3.2. MLST, cgMLST, Serotype, and ANI Analysis
The MLST profiles of diverse E. coli isolates, determined by specific gene alleles, revealed both shared characteristics and unique distinctions in their genetic composition (Table 1). Notably, isolates EC0002, EC110, EC119, EC120, EC123, EC126, EC331, EC335, EC338, EC339, EC1110, and EC2110 exhibited identical alleles across all seven MLST genes, resulting in the assignment of ST93. In contrast, isolates EC15 and ECV01 displayed distinctive alleles for several MLST genes, leading to the assignment of distinct sequence types (ST1148 and ST1196, respectively). The serotype profiles of E. coli isolates unveiled a predominant pattern across the strains (Table 1). Most of the isolates consistently exhibited O7 and H4 antigen types. In contrast, EC15 and ECV01 presented distinct serotypes with O163 and H7, and O22 and H28 antigens, respectively. To assess genomic relatedness and delineate distinctions among the E. coli isolates, we computed pairwise ANI for all possible genome pairs (Figure 2). The E. coli isolates were categorized into two groups, with EC15 and ECV01 forming a distinct, independent group. The ANI values across the bacterial isolates ranged from 98.33 to 100.00%.

Table 1.
Multilocus sequence and serotyping of E. coli isolates.
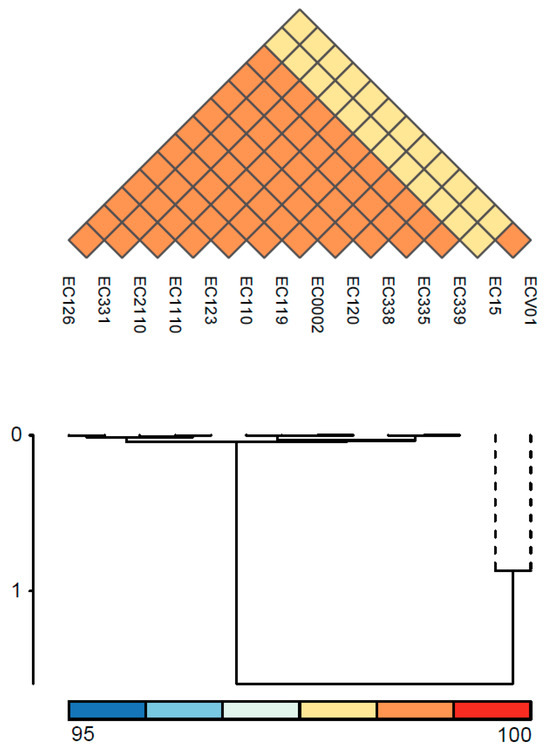
Figure 2.
ANI analysis of E. coli isolates. The color indicates the value of ANI; the value range is 95–100, with the color turning from blue to red.
3.3. Pangenome Analysis
For a more in-depth exploration of variations in the complete set of genes among E. coli strains, we conducted pangenome analysis utilizing the CGView server and its genome visualization tool. E. coli ATCC 9738 served as the reference genome, and the complete genomes of all isolates were included to generate the circular representation (Figure 3). The pangenome comparison results indicate that the genomes of the 12 isolates, excluding EC15 and ECV01, exhibited remarkably high similarity. Bacteriophage gene clusters, including portal protein, head-to-tail joining protein, lambda head decoration protein, major capsid protein, tail assembly protein, tail fiber protein, tail length tape measure protein, minor tail protein, lysin, DNA packaging protein, terminase, methyltransferase, and exonuclease, along with toxin–antitoxin system proteins and Hok/gef cell toxic protein were specifically present in the 2263–2310 kb region of EC15 (Table S1). Conversely, flagella gene clusters, encompassing cytoplasmic chaperone, motor, export apparatus, hook-filament junction, rod, ring, and flagellar type III secretion system components, were exclusively identified in the 3673–3711 kb region of ECV01 (Table S1).
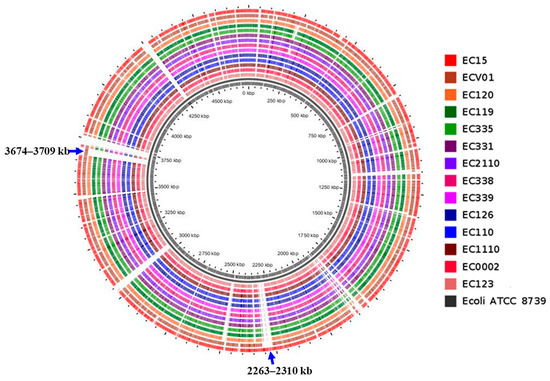
Figure 3.
Pangenome mapping of E. coli genomes. The pangenome was constructed using CGView, involving the incorporation of distinctive regions into the reference genome (E. coli ATCC 8739). Gaps in the map signify regions where genes are absent in the reference genome but are present in other genomes. Pangenome analysis revealed a high degree of similarity among most plasmids. However, a small number contained additional DNA sequences incorporating specific genes (Figure 4A,B). We observed notable differences in the sequences of the pEC15-5 and pEC331-2 plasmids compared to others (Figure 4A,B). Specifically, pEC15-5 carried DNA replication and transfer genes, including rop, mobC, mbeB, and mbeD, while pEC331-2 contained tetracycline-resistant genes, tetA and tetR. Additionally, all plasmids shared the presence of aminoglycoside- and sulfonamide-resistant genes, including aph(6)-Id, aph(3″)-Ib, and sul2 genes.
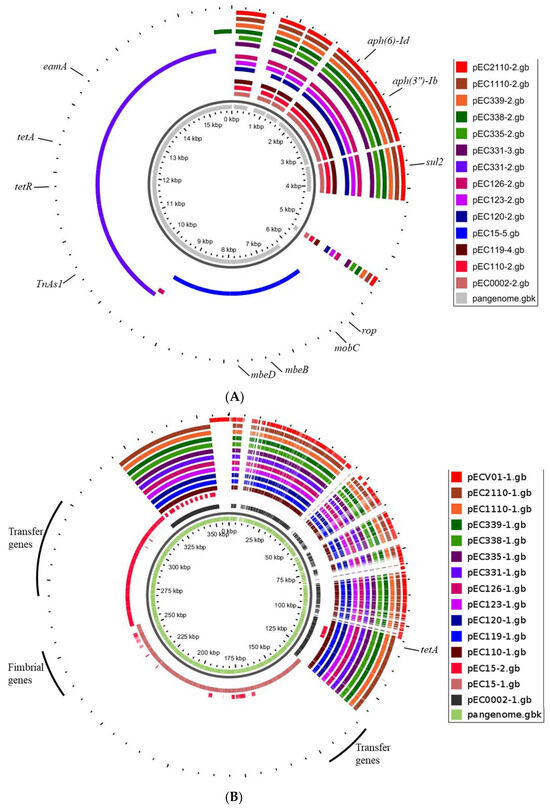
Figure 4.
Pangenome mapping of plasmids within the size range of 5.2–6.5 kb (A) and 107–124 kb (B) across E. coli isolates. The innermost circle depicts the pangenome in gray, while the outer circles represent individual plasmids.
3.4. Antimicrobial Resistance and Virulence Genes in Chromosomes
Aminoglycoside, chloramphenicol, quaternary ammonium compound, and trimethoprim/sulfamethoxazole resistance genes, including aac(3)-II,III,IV,VI,VIII,IX,X, aac(3)-IIa, aac(3)-IId, aadA2, aph(3″)-Ib, aph(6)-Id, catA2, qacE, dfrA12, sul1, and sul2, were universally detected in all isolates, with the exception of EC15 and ECV01 (Figure 5). Additionally, ECV01 exclusively harbored aadA1, cmlA1, and sul3 genes. Mutation patterns in key antibiotic resistance genes varied among isolates, with unique patterns for colistin (pmrB_Y358N in EC15 and ECV01) and quinolones (gyrA_D87N and gyrA_S83L shared; parC_E84K in all except EC15 and ECV01; parC_E84G exclusive to EC15; parC_S80I in EC15 and ECV01). The fosfomycin resistance-associated cyaA_S352T was absent in EC15 and ECV01, while glpT_E448K was universal, and uhpT_E350Q was exclusive to ECV01.

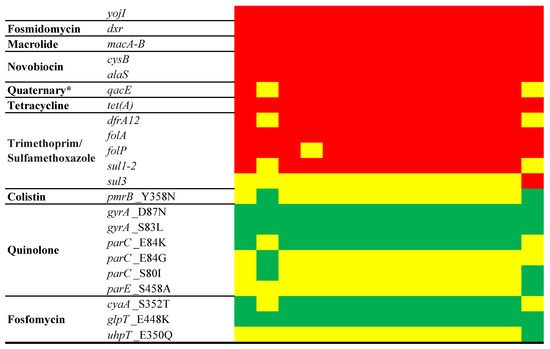
Figure 5.
Antimicrobial resistance determinants in the E. coli chromosome. The figure shows the presence (red for antimicrobial resistance genes or green for point mutations) or absence (yellow) of antimicrobial resistance genes. * Quaternary ammonium compound.
E. coli isolates exhibited varying numbers of genomic islands (GIs) and prophages. A clear pattern linking the number of GIs/prophages to the carriage of antibiotic resistance and virulence genes was not evident in Table 2. Isolate EC0002 contained 18 GIs and 9 prophages, hosting antibiotic resistance genes (aac(3)-Iid, aadA2, blaTEM-1B, dfrA12, qacE, and sul1), alongside virulence genes (csgB, csgD-G, espL1, espX1, espX4, fimB, fimE, PAAR, rfaD, rhs, tssI, and vgrG). In contrast, EC15, featuring 17 GIs and 7 prophages, displayed a repertoire of virulence genes such as acrB, cap8E, csgA-G, espL1, espX4, fimA-C, fimE, fimI, hsiB1, hsiC1, spaP-Q, and vipA-B but lacked antibiotic resistance genes. ECV01, characterized by 17 GIs and 5 prophages, exhibited a notable abundance of virulence genes without antibiotic resistance genes. Remarkably, rfaD was identified in EC0002, EC120, EC123, EC335, and ECV01, known for their association with enhanced biofilm formation and resistance to host defenses. Mobile element analysis showed commonalities (IS609, ISEc1, and MITEEc1) in all isolates except EC15 and ECV01 (Table 3). EC15 had IS3, IS609, ISEc1, ISSen1, and MITEEc1, while ECV01 had IS609, IS621, ISEc1, ISKpn8, ISSen1, and MITEEc1 exclusively.
Table 2.
Antibiotic resistance and virulence genes in genomic islands (GIs) and prophages of the chromosome. Bold: identified from prophages.
Table 3.
Mobile elements of E. coli chromosome. Bold: common in all isolates. Italics: common in all isolates except EC15 and ECV01.
A set of adhesion/invasion genes (aslA, csgB, csgD-G, cusC, ecpA-E, ecpR, fdeC, fimA-I, flgC, flgG-H, fliG, fliM, fliP-Q, focA, hcpA, pitB, ppdD, serA, ydiV, and yijP) was consistent across the chromosomes of all isolates (Figure 6). The intriguing absence of the flagellin (fliC) gene in all isolates except for EC15 and ECV01 added complexity to the observed gene profile variations among these isolates. Biofilm-related genes had consistent patterns, with gatC, gatZ, kbaY, and luxS present in all but pgaA absent in EC15 and ECV01. Capsule gene analysis revealed variable prevalence, with etk, gfcA, gfcC-E, and ymcC consistently present; however, gfcB varied, being absent in EC15 and ECV01. Iron acquisition genes entA-F, entS, fepA-D, fepG, fes, and fur were uniformly present, indicating shared iron acquisition capacity.





Figure 6.
Virulence determinants in the E. coli chromosome. The figure shows the presence (red) or absence (yellow) of virulence genes.
The prevalence of toxin genes varied, with the outer membrane efflux lipoprotein (ibeB) and outer membrane protein A (ompA) genes universally present. However, capsule polysaccharide synthesis genes were notably absent in EC15 and ECV01. Most genes associated with the type II (gspC-M) and III (espL, espX, and map) secretion systems were uniformly present, but EC15 and ECV01 lacked espL4. Genes related to the type I secretion system (nuoG) and general autotransporter pathway (gatB) genes were consistently present across all isolates.
3.5. Antimicrobial Resistance and Virulence Genes in Plasmids
Plasmids, including pEC0002-1, pEC110-1, pEC119-1, pEC120-1, pEC123-1, pEC126-1, pEC331-1, pEC335-1, pEC338-1, pEC339-1, pEC1110-1, and pEC2110-1, consistently carried resistance genes like catA2, folA, tetA, and tetC (Table 4). Conversely, plasmids such as pEC15-3, pEC15-4, and pEC15-5 appeared devoid of discernible antimicrobial resistance genes. Notably, pECV01-2 emerged as a comprehensive reservoir of antimicrobial resistance genes, featuring aadA1, aadA2, ant(3″)-Ia, blaTEM-1B, cmlA1, dfrA12, mefB, qacE, sul3, tetA, tetR, and vgaC. EC15 and ECV01 isolates exclusively possessed the streptogramin A resistance gene vgaC, distinguishing them from the remaining isolates.
Table 4.
Antibiotic resistance and virulence genes, replicon types, and sizes of E. coli plasmids.
Plasmids such as pEC15-1 and pEC15-2 showed an array of virulence genes, including caf1A, caf1M, faeA, faeC-I, finP, ipaH, and virB (Table 4); however, several plasmids exhibited no discernible virulence genes. Interestingly, pEC15-3 and pECV01-2 stood out as they possessed the virulence regulon transcriptional activator (virB) gene. The analysis of plasmid replicon types across the diverse set of E. coli plasmids revealed distinct patterns in their replication machinery (Table 4). Plasmids exceeding 123 kb in size exhibited a consistent IncFIB replicon type. Significantly, pEC15-2, pEC15-3, and pECV01-2 distinguished themselves by featuring IncI1-I, IncY, and IncR replicon types, setting them apart from the predominant IncFIB-dominated plasmids.
3.6. Mobile Elements in Plasmids
Integration/excision genes (insO1, intA, and tnpA) were present in all E. coli strains with plasmids of 123 kb in size, except for EC15 and ECV01 (Table 5). Notably, pECV01-2 showed an extensive set of integration/excision genes (int, intM, ltrA, tnp, tnp1, tnpA, and tnpR), while pEC15-3 uniquely carried ref and rdgC. Plasmids such as pEC15-1, pEC15-2, and pEC15-3 exhibited unique phage genes, including ybiI_1, exc, and nusG, and an extensive array associated with phage functions. Conversely, other plasmids shared common phage genes cor and smc. Plasmids carrying the type I restriction–modification system gene (hsdR) exhibited commonalities in their defense mechanisms.
Table 5.
Mobile elements of E. coli plasmids.
In contrast, pEC15-2 and pEC15-3 carried antirestriction protein (ardA) and toxin-antitoxin system gene (ccdA-B), respectively. Plasmids such as pEC15-1 were equipped with an extensive set of transfer genes, including finO, psiB, traA-E, traG-H, traK, traL-N, traT-U, traW-X, trbD, and trbI. Conversely, pEC15-2 featured a different repertoire, with mobilization genes such as mobB, nikB, pilI, pilK-U, psiB, sogL, traA, traC-R, traT-W, traY, trbA-C, and yggA. Notably, pEC15-5 possessed mbeB, mbeD, and mobC, further contributing to its potential for efficient plasmid transfer. pEC15-2 featured a different set of insertion sequences, including ISEc25, ISEc37, ISEc43, Tn2, TnpA_Tn3, TnpA_TnAs1, TnpR_Tn3, and TnpR_TnEc1. Plasmids like pEC331-2 exhibited specific insertion sequences such as IS15DI and TnpA_TnAs1, contributing to the genetic diversity among these elements.
We constructed circular diagrams to facilitate the comparison of plasmid backbones. The EC15 isolate exhibited a spectrum of plasmids, ranging from the large pEC15-1 (144 kb, IncFIB) to the compact pEC15-5 (5 kb, IncFIB) (Figure S1A–E). The comparison of gene profiles between two E. coli plasmids, pEC15 and pECV01, revealed both shared and distinct genetic elements within their compositions. Plasmid pEC15-1 carried virulence genes (caf1A, caf1M, and virB), fimbrial biosynthesis genes (faeC-I), plasmid replication genes (repA and repB), transfer genes (traC-G, traK, traM-N, traR, and traU-X), and various insertion sequences (IS2, IS3, IS91, and IS629). Plasmid pEC15-2, on the other hand, featured antibiotic resistance (blaTEM-1B, tetA, and tetC), mobilization (mobC and nikB), and transfer (traA-B, traE-F, traH-I, and traM-Y) genes. Plasmid pEC15-3 exhibited genes associated with phage activity (kilA), replication (repA and repL), and stability (parA). In contrast, plasmid pEC15-4 included psiB, while pEC15-5 harbored genes associated with mobilization (mbeA, mbeD, and mobC) and regulation (rop). pECV01-1 encompassed a range of genes involved in replication (dnaE and dnaQ), restriction–modification systems (hsdR), and transfer (parB, recA, and repA) (Figure S2A,B). Notably, pECV01-2 had a comprehensive collection of antibiotic resistance genes (blaTEM-1B, tetA, tetR, and cmlA1) and virulence genes (virB), indicating a potential dual role.
4. Discussion
The MLST profiles provided insights into the genetic relatedness among the E. coli isolates, with most sharing a common sequence type (ST93). The shared alleles across seven MLST genes suggest potential clonality or a common evolutionary lineage among EC0002, EC110, EC119, EC120, EC123, EC126, EC331, EC335, EC338, EC339, EC1110, and EC2110. ST93 has been sporadically identified as both avian and human extra-intestinal pathogenic or diarrheagenic E. coli in various regions, encompassing humans, animals, and globally distributed food products [25,26]. Previous studies have reported its presence in beef, veal, pork, and poultry in Switzerland [27], a pig in Laos [28], a cat and retail food in China [29], broiler chickens in Brazil [30], and patients in Finland and Uruguay [31,32]. ECV01 exhibited a unique MLST type, ST1196, which has notably been associated with MDR E. coli in livestock, pets, hospitals, and wastewater across different countries, thus playing a pivotal role in the global transmission of antimicrobial resistance genes [33,34]. cgMLST also mirrors this pattern observed with MLST. Strains EC15 and ECV01 displayed distinct cgMLST types, 32199 and 69511, respectively, whereas all other isolates belonged to cgMLST type 39996.
The serotype analysis further supports the MLST findings, with most isolates exhibiting a consistent serological identity (O7:H4). Interestingly, a recent outbreak of food poisoning in Japan occurred among individuals who consumed seaweed [35]. The culprit behind the illness was identified as O7:H4 serotype E. coli. Like our strains of E. coli, this variant also exhibited resistance to β-lactam antibiotics. Unique serotypes of EC15 (O163:H7) and ECV01 (O22:H28) underscore the diversity within this bacterial population, suggesting distinct evolutionary trajectories. ANI analysis provided additional granularity to the genomic relationships among isolates, categorizing them into two groups. EC15 and ECV01 formed a distinct group, reinforcing their genetic distinctiveness. The high ANI values within each group signify genomic similarity, while the variations between groups highlight the genetic diversity in the E. coli population.
In aquaculture, the acknowledged prophylactic and therapeutic application of tetracycline has emerged as a significant contributor to the dissemination of tetracycline resistance genes in the environment [36]. Remarkably, these resistance genes were persistent in aquaculture settings, even in the absence of continual tetracycline use [37]. Notably, all E. coli isolates in our study were found to carry both tetA and tetC genes. Consistent with our findings, earlier research has highlighted the prevalence of tetracycline resistance genes among bacteria recovered from imported shrimps originating from Asian countries [38].
Moreover, investigations have indicated the dissemination of plasmids carrying tetracycline resistance determinants between aquaculture and human environments [39]. The presence of chloramphenicol residue is a common cause for the rejection of imported shrimp in the U.S. [40], indicative of the widespread use of banned antibiotics in shrimp-exporting countries [41]. Previous investigations into E. coli isolates from fish and shrimp in Vietnam have revealed a significant number of isolates resistant to chloramphenicol [42,43]. Elevated occurrences of chloramphenicol resistance genes have also been documented in E. coli isolates from fish, shrimp, and shellfish in China [44]. Ng et al. detected E. coli isolates containing catA2 from aquaculture farms in Malaysia [45]. Notably, all E. coli isolates from imported shrimp were resistant to chloramphenicol and harbored the chloramphenicol resistance genes catA2 and cmlA1.
Sulfonamide is extensively used in animal production systems, either as a standalone drug or in conjunction with diaminopyrimidines [46]. Notably, a heightened prevalence of sulfonamide resistance has been documented in enteric bacteria isolated from both livestock and humans [47,48]. Gram-negative bacteria exhibiting resistance to sulfonamides have been reported in shrimp farms in Vietnam [49]. This resistance is frequently linked to the acquisition of sul2 [48]. The sul2 gene has predominantly been identified on small plasmids, and this pattern extends to our E. coli isolates from imported shrimp, where sul2 was also located on small plasmids. The identification of chloramphenicol (catA2), aminoglycoside- (aph(3″)-Ib and aph(6)-Id), tetracycline- (tetA and tetC), trimethoprim- (folA), and sulfonamide (sul2)-resistant genes within plasmids of the E. coli isolates from imported shrimp emphasizes their contribution to the dissemination of antibiotic resistance.
The comparative analysis of the chromosomes and plasmids of diverse E. coli isolates sheds light on the intricate interplay between intrinsic and extrachromosomal genetic elements, providing a comprehensive understanding of the factors influencing antibiotic resistance, virulence, and adaptability within this bacterial population. Examination of antimicrobial resistance genes in both chromosomes and plasmids reveals distinct patterns. While the chromosomes harbored a conserved set of resistance genes, including aac(3)-II,III,IV,VI,VIII,IX,X, aac(3)-IIa, aac(3)-IId, aadA2, aph(3″)-Ib, aph(6)-Id, catA2, pmrE, qacE, dfrA12, sul1, and sul2, the plasmids exhibited variability in resistance profiles. Plasmids such as pEC15-3, pEC15-4, pEC15-5, and pEC119-3 did not contain discernible antimicrobial resistance genes. In contrast, plasmids like pECV01-2 emerged as comprehensive reservoirs of antimicrobial resistance genes, hosting a diverse array that includes aadA1, aadA2, ant(3″)-Ia, blaTEM-1B, cmlA1, dfrA12, mefB, qacE, sul3, tetA, tetR, and vgaC. This suggests a potential reliance on plasmid determinants for antibiotic resistance in this isolate.
Antibiotic resistance ensures bacterial, and consequently plasmid, survival when the relevant resistance gene is carried on the plasmid [50]. However, the transfer of the gene to the chromosome preserves the selective advantage of resistance while alleviating the fitness cost associated with replicating large plasmids. Among the examined isolates, twelve were found to carry the genes aadA2, qacE, and dfrA12; notably, only one isolate was observed to harbor these genes within a plasmid, while the rest housed them in the chromosome. This observation aligns with the proposed evolutionary pressure for plasmid-borne antimicrobial resistance genes to move to the chromosome. On the other hand, certain determinants such as catA2, tetC, tetR, and vgaC genes were exclusively found on plasmids, highlighting the diversity in the genetic distribution of resistance determinants among bacterial populations.
Antibiotic resistance genes associated with efflux pumps are predominantly encoded on chromosomes, with E. coli efflux pump genes like acrAB-tolC, acrAD-tolC, acrEF-tolC, mdtABC-tolC, and mdtEF-tolC often integrated into the core genome, serving diverse cellular functions [51,52]. Various plasmid-borne efflux pump genes, including tetA, tetX3, tetX4, tetX5, qepA, oqxAB, acrR, tmexCD1-toprJ1, sugE, qacA/B, qacA, qacG, and qacH, have been identified in both Gram-negative and Gram-positive bacteria, contributing to antibiotic resistance [53,54,55]. However, several efflux pump genes, such as acid resistance transcriptional activators (gadE, gadW, and gadX), resistance-nodulation-division-type efflux pumps (mdtE and mdtF), and a metabolite, H1 symporter (MHS) family major facilitator superfamily transporter (MNZ41_23075), were found in the plasmid of the EC119 isolate. To our knowledge, there has been no reported instance of bacteria harboring such a diverse combination of efflux pump genes within a plasmid. The unique presence of this combination in the plasmid of EC119 underscores the need for further research that would enhance our understanding of its implications for antibiotic resistance.
Our analysis of mobile genetic elements in plasmids illuminates their remarkable potential for movement and adaptation within bacterial populations. The presence of integration/excision genes (insO1, intA, and tnpA) in the majority of plasmids suggests their capacity for mobilization. These genes function akin to molecular cut-and-paste tools, facilitating the integration of plasmid DNA into the host chromosome, potentially disseminating antimicrobial resistance and virulence genes within the bacterium [56]. Notably, plasmid pECV01-2 harbors an extensive array of integration/excision genes (int, intM, ltrA, tnp, tnp1, tnpA, and tnpR), indicating a potentially heightened role in its own mobilization compared to other plasmids. This dynamic interaction suggests active participation of the plasmid in seeking opportunities to spread its genetic payload. Distinct sets of transfer genes across plasmids imply diverse dissemination strategies. Plasmid pEC15-1 appears well equipped with a comprehensive set of conjugation-related genes like finO, traA-E, and traG-H, facilitating the direct transfer of plasmid DNA between bacterial cells. Furthermore, the presence of mbeB, mbeD, and mobC in pEC15-5 suggests their involvement in plasmid mobilization, indicating a sophisticated network of transfer mechanisms. These findings underscore the adaptability of plasmids, employing diverse strategies for successful dissemination within bacterial communities. Beyond transfer, our analysis reveals mechanisms ensuring plasmid stability within host bacteria. Notably, pEC15-3 carries a toxin–antitoxin system gene (ccdA-B), serving as a “molecular suicide switch” where a loss of the plasmid triggers host cell inactivation by the toxin [57]. This strategy confers a selective advantage to plasmid-carrying cells, promoting their survival and proliferation, thus ensuring plasmid persistence alongside host lineages. Interestingly, pEC15-2 encodes an antirestriction protein (ardA), potentially countering host or other mobile genetic element-encoded restriction enzymes [58], safeguarding plasmid integrity against degradation.
Analysis of virulence gene categories, spanning adhesin, invasion, biofilm formation, capsule, chemotaxis, iron acquisition, toxin, and secretion system genes, revealed the intricate landscape of genetic determinants within chromosomes. In the EC15 isolate, genes associated with adherence, invasion, and capsule were housed in plasmids. Unique to EC15’s plasmids were ipaH and virB genes, implicated in triggering cell death and modulating host inflammatory signals during bacterial infection [59]. The EC15 isolate also harbored K88 fimbrial biosynthesis genes on plasmids, known as major colonization factors in some enterotoxigenic E. coli strains associated with porcine neonatal and postweaning diarrhea [60]. The concurrent presence of type 1C and K88 fimbrial biosynthesis genes on chromosomes (fimA-I) and plasmids (faeA and faeC-I) suggests a collaborative role in adherence, a crucial aspect of bacterial tissue colonization. The presence of distinct capsule genes on both the chromosome (etk, gfcA-E, and ymcC) and plasmid (caf1A, caf1M) in the EC15 isolate implies a sophisticated immune evasion strategy, potentially surpassing that of other E. coli strains. The findings that we obtained from comprehensive whole-genome analyses of the E. coli isolates offer insights into the molecular basis of MDR and virulence in E. coli.
5. Conclusions
In summary, comprehensive analysis of the genetic elements in the chromosomes and plasmids of diverse E. coli isolates provides a holistic understanding of the factors influencing antibiotic resistance, pathogenicity, and adaptability. The intricate interplay between mobile elements, genomic islands, and various functional gene categories contributes to the dynamic nature of the E. coli population and has implications for public health, emphasizing the need for continued surveillance and research into the evolution of these important bacterial pathogens.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/foods13111766/s1, Figure S1: Comparative visualization of the identified plasmids. pEC15-1 (A), pEC15-2 (B), pEC15-3 (C), pEC15-4 (D), and pEC15-5 (E); Figure S2: Comparative visualization of the identified plasmids. pECV01-1 (A) and pECV01-2 (B); Table S1: Gene lists of unique regions on EC15 and ECV01.
Author Contributions
Conceptualization, K.S. and M.N.; methodology, K.S., M.N., M.P., J.C., K.A., J.R., J.A.M.; software, K.S.; validation, K.S.; formal analysis, K.S.; investigation, K.S., M.N.; resources, K.S., M.N.; data curation, K.S.; writing—original draft preparation, K.S.; writing—review and editing, K.S., M.N., M.P., J.C., K.A., S.A.K., K.A., J.R., J.A.M., A.A.K.; visualization, K.S.; supervision, K.S., M.N.; project administration, M.N.; funding acquisition, M.N. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by protocol number E0782801 from the U.S. Food and Drug Administration.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Materials, further inquiries can be directed to the corresponding author.
Acknowledgments
We thank Seong-jae Kim and Jing Han for reviewing the manuscript, and Joanne Berger, FDA Library, for editing a draft of the manuscript.
Conflicts of Interest
This manuscript reflects the views of the authors and does not necessarily reflect those of the U.S. Food and Drug Administration and the Saudi Food and Drug Authority. Any mention of commercial products is for clarification only and is not intended as approval, endorsement, or recommendation.
References
- National Fisheries Institute Media. Available online: https://aboutseafood.com/about/top-ten-list-for-seafood-consumption/ (accessed on 16 December 2023).
- National Marine Fisheries Service. Fisheries of the United States, 2020. U.S. Department of Commerce, NOAA Current Fishery Statistics No. 2020. Available online: https://www.fisheries.noaa.gov/national/sustainable-fisheries/fisheries-united-states (accessed on 16 December 2023).
- Holmström, K.; Gräslund, S.; Wahlström, A.; Poungshompoo, S.; Bengtsson, B.E.; Kautsky, N. Antibiotic use in shrimp farming and implications for environmental impacts and human health. Int. J. Food Sci. Technol. 2003, 38, 255–266. [Google Scholar] [CrossRef]
- Swapna, K.M.; Rajesh, R.; Lakshmanan, P.T. Incidence of antibiotic residues in farmed shrimps from the southern states of India. Indian. J. Geo-Mar. Sci. 2012, 41, 344–347. [Google Scholar]
- Love, D.C.; Rodman, S.; Neff, R.A.; Nachman, K.E. Veterinary Drug Residues in Seafood Inspected by the European Union, United States, Canada, and Japan from 2000 to 2009. Environ. Sci. Technol. 2011, 45, 7232–7240. [Google Scholar] [CrossRef] [PubMed]
- Boss, R.; Overesch, G.; Baumgartner, A. Antimicrobial Resistance of Escherichia coli, Enterococci, Pseudomonas aeruginosa, and Staphylococcus aureus from Raw Fish and Seafood Imported into Switzerland. J. Food Prot. 2016, 79, 1240–1246. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.A.; Islam, M.S.; Rana, M.L.; Ferdous, F.B.; Neloy, F.H.; Firdous, Z.; Hassan, J.; Rahman, M.T. Resistance Profiles and Virulence Determinants in Biofilm-Forming Enterococcus faecium Isolated from Raw Seafood in Bangladesh. Pathogens 2023, 12, 1101. [Google Scholar] [CrossRef] [PubMed]
- Heuer, O.E.; Kruse, H.; Grave, K.; Collignon, P.; Karunasagar, I.; Angulo, F.J. Human health consequences of use of antimicrobial agents in aquaculture. Clin. Infect. Dis. 2009, 49, 1248–1253. [Google Scholar] [CrossRef] [PubMed]
- Naik, O.A.; Shashidhar, R.; Rath, D.; Bandekar, J.R.; Rath, A. Characterization of multiple antibiotic resistance of culturable microorganisms and metagenomic analysis of total microbial diversity of marine fish sold in retail shops in Mumbai, India. Environ. Sci. Pollut. Res. Int. 2018, 25, 6228–6239. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; CLSI Standard: Wayne, PA, USA, 2019. [Google Scholar]
- Alotaibi, K.; Revollo, J.; Sung, K.; Miranda, J.A.; Khan, S.A.; Khan, A.A.; Nawaz, M. Draft Genome Sequences of 14 Fluoroquinolone-Resistant Escherichia coli Isolates from Imported Shrimp. Microbiol. Resour. Announc. 2023, 12, e0111622. [Google Scholar] [CrossRef] [PubMed]
- Center for Genomic Epidemiology. Available online: https://www.genomicepidemiology.org (accessed on 25 November 2023).
- Liu, D.; Zhang, Y.; Fan, G.; Sun, D.; Zhang, X.; Yu, Z.; Wang, J.; Wu, L.; Shi, W.; Ma, J. IPGA: A handy integrated prokaryotes genome and pan-genome analysis web service. iMeta 2022, 1, e55. [Google Scholar] [CrossRef]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L.; et al. The comprehensive antibiotic resistance database. Antimicrob. Agents Chemother. 2013, 57, 3348–3357. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulos, D.A.; Assaf, R.; Aziz, R.K.; Brettin, T.; Bun, C.; Conrad, N.; Davis, J.J.; Dietrich, E.M.; Disz, T.; Gerdes, S.; et al. PATRIC as a unique resource for studying antimicrobial resistance. Brief Bioinform. 2019, 20, 1094–1102. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.L.; Mullet, J.; Hindi, F.; Stoll, J.E.; Gupta, S.; Choi, M.; Keenum, I.; Vikesland, P.; Pruden, A.; Zhang, L. mobileOG-db: A Manually Curated Database of Protein Families Mediating the Life Cycle of Bacterial Mobile Genetic Elements. Appl. Environ. Microbiol. 2022, 88, e0099122. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.H.K.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Goh, Y.X.; Tai, C.; Wang, H.; Deng, Z.; Ou, H.Y. VRprofile2: Detection of antibiotic resistance-associated mobilome in bacterial pathogens. Nucleic Acids Res. 2022, 50, W768–W773. [Google Scholar] [CrossRef] [PubMed]
- Sayers, S.; Li, L.; Ong, E.; Deng, S.; Fu, G.; Lin, Y.; Yang, B.; Zhang, S.; Fa, Z.; Zhao, B.; et al. Victors: A web-based knowledge base of virulence factors in human and animal pathogens. Nucleic Acids Res. 2019, 47, D693–D700. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A reference database for bacterial virulence factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef] [PubMed]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.R.; Arantes, A.S.; Stothard, P. Comparing thousands of circular genomes using the CGView Comparison Tool. BMC Genomics 2012, 13, 202. [Google Scholar] [CrossRef] [PubMed]
- Maluta, R.P.; Logue, C.M.; Casas, M.R.; Meng, T.; Guastalli, E.A.; Rojas, T.C.; Montelli, A.C.; Sadatsune, T.; de Carvalho Ramos, M.; Nolan, L.K.; et al. Overlapped sequence types (STs) and serogroups of avian pathogenic (APEC) and human extra-intestinal pathogenic (ExPEC) Escherichia coli isolated in Brazil. PLoS ONE 2014, 9, e105016. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, X.; Zheng, S.; Yu, F.; Kong, H.; Yang, Q.; Cui, D.; Chen, N.; Lou, B.; Li, X.; et al. Serotypes, genotypes and antimicrobial resistance patterns of human diarrhoeagenic Escherichia coli isolates circulating in southeastern China. Clin. Microbiol. Infect. 2014, 20, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Vogt, D.; Overesch, G.; Endimiani, A.; Collaud, A.; Thomann, A.; Perreten, V. Occurrence and genetic characteristics of third-generation cephalosporin-resistant Escherichia coli in Swiss retail meat. Microb. Drug Resist. 2014, 20, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Thongmalayvong, B.; Akkhavong, K.; Somphavong, S.; Paboriboune, P.; Khounsy, S.; Morand, S.; Rolain, J.M. Clonal transmission of a colistin-resistant Escherichia coli from a domesticated pig to a human in Laos. J. Antimicrob. Chemother. 2015, 70, 3402–3404. [Google Scholar] [PubMed]
- Zhang, X.F.; Doi, Y.; Huang, X.; Li, H.Y.; Zhong, L.L.; Zeng, K.J.; Zhang, Y.F.; Patil, S.; Tian, G.B. Possible Transmission of mcr-1–Harboring Escherichia coli between Companion Animals and Human. Emerg. Infect. Dis. 2016, 22, 1679–1681. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.C.; Penha, R.A.C.; Andrade, L.N.; Berchieri, A.; Darini, A.L.C. Detection of chromosomal blaCTX-M-2 in diverse Escherichia coli isolates from healthy broiler chickens. Clin. Microbiol. Infect. 2014, 20, O623–O626. [Google Scholar] [CrossRef]
- Grondahl-Yli-Hannuksela, K.; Lonnqvist, E.; Kallonen, T.; Lindholm, L.; Jalava, J.; Rantakokko-Jalava, K.; Vuopio, J. The first human report of mobile colistin resistance gene, mcr-1, in Finland. APMIS 2018, 126, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Papa-Ezdra, R.; Grill Diaz, F.; Vieytes, M.; García-Fulgueiras, V.; Caiata, L.; Ávila, P.; Brasesco, M.; Christophersen, I.; Cordeiro, N.F.; Algorta, G.; et al. First three Escherichia coli isolates harbouring mcr-1 in Uruguay. J. Glob. Antimicrob. Resist. 2020, 20, 187–190. [Google Scholar] [CrossRef]
- Lima, L.S.; Proietti-Junior, A.A.; Rodrigues, Y.C.; Vieira, M.C.d.S.; Lima, L.N.G.C.; Souza, C.d.O.; Gonçalves, V.D.; Lima, M.d.O.; Rodrigues, D.d.P.; Lima, K.V.B. High Genetic Diversity and Antimicrobial Resistance in Escherichia coli Highlight Arapaima gigas (Pisces: Arapaimidae) as a Reservoir of Quinolone-Resistant Strains in Brazilian Amazon Rivers. Microorganisms 2022, 10, 808. [Google Scholar] [CrossRef]
- Delgado-Blas, J.F.; Ovejero, C.M.; David, S.; Montero, N.; Calero-Caceres, W.; Garcillan-Barcia, M.P.; de la Cruz, F.; Muniesa, M.; Aanensen, D.M.; Gonzalez-Zorn, B. Population genomics and antimicrobial resistance dynamics of Escherichia coli in wastewater and river environments. Commun. Biol. 2021, 4, 457. [Google Scholar] [CrossRef] [PubMed]
- Kashima, K.; Sato, M.; Osaka, Y.; Sakakida, N.; Kando, S.; Ohtsuka, K.; Doi, R.; Chiba, Y.; Takase, S.; Fujiwara, A.; et al. An outbreak of food poisoning due to Escherichia coli serotype O7:H4 carrying astA for enteroaggregative E. coli heat-stable enterotoxin1 (EAST1). Epidemiol. Infect. 2021, 149, e244. [Google Scholar] [CrossRef] [PubMed]
- Sivaraman, G.K.; Rajan, V.; Vijayan, A.; Elangovan, R.; Prendiville, A.; Bachmann, T.T. Antibiotic Resistance Profiles and Molecular Characteristics of Extended-Spectrum Beta-Lactamase (ESBL)-Producing Escherichia coli and Klebsiella pneumoniae Isolated From Shrimp Aquaculture Farms in Kerala, India. Front. Microbiol. 2021, 12, 622891. [Google Scholar] [CrossRef] [PubMed]
- Tamminen, M.; Karkman, A.; Lohmus, A.; Muziasari, W.I.; Takasu, H.; Wada, S.; Suzuki, S.; Virta, M. Tetracycline resistance genes persist at aquaculture farms in the absence of selection pressure. Environ. Sci. Technol. 2011, 45, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Ellis-Iversen, J.; Seyfarth, A.M.; Korsgaard, H.; Bortolaia, V.; Munck, N.; Dalsgaard, A. Antimicrobial resistant E. coli and Enterococci in pangasius fillets and prawns in Danish retail imported from Asia. Food Control 2020, 114, 106958. [Google Scholar] [CrossRef]
- Rhodes, G.; Huys, G.; Swings, J.; Mcgann, P.; Hiney, M.; Smith, P.; Pickup, R.W. Distribution of oxytetracycline resistance plasmids between aeromonads in hospital and aquaculture environments: Implication of Tn1721 in dissemination of the tetracycline resistance determinant tet A. Appl. Environ. Microbiol. 2000, 66, 3883–3890. [Google Scholar] [CrossRef]
- Food and Drug Administration (FDA). Approved Aquaculture Drugs. 2020. Available online: https://www.fda.gov/animal-veterinary/aquaculture/approved-aquaculture-drugs (accessed on 16 December 2023).
- Sharma, L.; Nagpal, R.; Jackson, C.R.; Patel, D.; Singh, P. Antibiotic-resistant bacteria and gut microbiome communities associated with wild-caught shrimp from the United States versus imported farm-raised retail shrimp. Sci. Rep. 2021, 11, 3356. [Google Scholar] [CrossRef]
- Nguyen, D.P.; Nguyen, T.A.D.; Le, T.H.; Tran, N.M.; Ngo, T.P.; Dang, V.C.; Kawai, T.; Kanki, M.; Kawahara, R.; Jinnai, M.; et al. Dissemination of extended-spectrum beta-lactamase- and AmpC beta-lactamase-producing Escherichia coli within the food distribution system of Ho Chi Minh City, Vietnam. BioMed Res. Int. 2016, 2016, 8182096. [Google Scholar] [CrossRef] [PubMed]
- Le, Q.P.; Ueda, S.; Nguyen, T.N.; Dao, T.V.; Van Hoang, T.A.; Tran, T.T.; Hirai, I.; Nakayama, T.; Kawahara, R.; Do, T.H.; et al. Characteristics of Extended-Spectrum beta-Lactamase-Producing Escherichia coli in Retail Meats and Shrimp at a Local Market in Vietnam. Foodborne Pathog. Dis. 2015, 12, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.H.; Huang, Y.B.; Yang, G.Z.; Wu, Q.P.; Zhang, J.; Wang, J.; Ding, Y.; Su, Y.; Ye, Q.H.; Wu, S.; et al. High prevalence of multidrug-resistant Escherichia coli in retail aquatic products in China and the first report of mcr-1-positive extended-spectrum β-lactamase-producing E. coli ST2705 and ST10 in fish. Int. J. Food Microbiol. 2024, 408, 110449. [Google Scholar] [CrossRef]
- Ng, K.H.; Samuel, L.; Kathleen, M.M.; Leong, S.S.; Felecia, C. Distribution and prevalence of chloramphenicol-resistance gene in Escherichia coli isolated from aquaculture and other environment. Int. Food Res. J. 2014, 21, 1321–1325. [Google Scholar]
- McEwen, S.A.; Fedorka-Cray, P.J. Antimicrobial use and resistance in animals. Clin. Infect. Dis. 2002, 34, S93–S106. [Google Scholar] [CrossRef] [PubMed]
- Enne, V.I.; Livermore, D.M.; Stephens, P.; Hall, L.M. Persistence of sulphonamide resistance in Escherichia coli in the UK despite national prescribing restriction. Lancet 2001, 357, 1325–1328. [Google Scholar] [CrossRef] [PubMed]
- Kozak, G.K.; Pearl, D.L.; Parkman, J.; Reid-Smith, R.J.; Deckert, A.; Boerlin, P. Distribution of sulfonamide resistance genes in Escherichia coli and Salmonella isolates from swine and chickens at abattoirs in Ontario and Quebec, Canada. Appl. Environ. Microbiol. 2009, 75, 5999–6001. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.T.H.; Rossi, P.; Dinh, H.D.K.; Pham, N.T.A.; Tran, P.A.; Ho, T.T.K.M.; Dinh, Q.T.; De Alencastro, L.F. Analysis of antibiotic multi-resistant bacteria and resistance genes in the effluent of an intensive shrimp farm (Long An, Vietnam). J. Environ. Manag. 2018, 214, 149–156. [Google Scholar] [CrossRef]
- Goswami, C.; Fox, S.; Holden, M.T.G.; Connor, M.; Leanord, A.; Evans, T.J. Origin, maintenance and spread of antibiotic resistance genes within plasmids and chromosomes of bloodstream isolates of Escherichia coli. Microb. Genom. 2020, 6, e000353. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Beltran, J.; Sorum, V.; Toll-Riera, M.; de la Vega, C.; Pena-Miller, R.; San Millan, A. Genetic dominance governs the evolution and spread of mobile genetic elements in bacteria. Proc. Natl. Acad. Sci. USA 2020, 117, 15755–15762. [Google Scholar] [CrossRef] [PubMed]
- Anes, J.; McCusker, M.P.; Fanning, S.; Martins, M. The ins and outs of RND efflux pumps in Escherichia coli. Front Microbiol. 2015, 6, 587. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria: An update. Drugs 2009, 69, 1555–1623. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Wan, M.; Wang, C.; Gao, X.; Yang, Q.; Partridge, S.R.; Wang, Y.; Zong, Z.; Doi, Y.; Shen, J.; et al. Emergence of a Plasmid-Encoded Resistance-Nodulation-Division Efflux Pump Conferring Resistance to Multiple Drugs, Including Tigecycline, in Klebsiella pneumoniae. mBio 2020, 11, e02930-19. [Google Scholar] [CrossRef]
- Hassanzadeh, S.; Ganjloo, S.; Pourmand, M.R.; Mashhadi, R.; Ghazvini, K. Epidemiology of efflux pumps genes mediating resistance among Staphylococcus aureus; A systematic review. Microb. Pathog. 2020, 139, 103850. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [PubMed]
- Markovski, M.; Wickner, S. Preventing bacterial suicide: A novel toxin-antitoxin strategy. Mol. Cell. 2013, 52, 611–612. [Google Scholar] [CrossRef]
- Nekrasov, S.V.; Agafonova, O.V.; Belogurova, N.G.; Delver, E.P.; Belogurov, A.A. Plasmid-encoded antirestriction protein ArdA can discriminate between type I methyltransferase and complete restriction-modification system. J. Mol. Biol. 2007, 365, 284–297. [Google Scholar] [CrossRef] [PubMed]
- Dranenko, N.O.; Tutukina, M.N.; Gelfand, M.S.; Kondrashov, F.A.; Bochkareva, O.O. Chromosome-encoded IpaH ubiquitin ligases indicate non-human enteroinvasive Escherichia. Sci. Rep. 2022, 12, 6868. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).