
Bactericidal Action of Shrimp Shell Chitooligosaccharide Conjugated with Epigallocatechin Gallate (COS-EGCG) against Listeria monocytogenes

 ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Bacterial Strains and Culture Condition
2.3. Determination of Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
2.4. Time-Kill Assay
2.5. Effect of COS-EGCG Conjugate on Extracellular Polysaccharide Production
2.6. Prevention of Biofilm Formation and Biofilm Inactivation of L. monocytogenes by COS-EGCG Conjugate
2.7. Protein Leakages of L. monocytogenes F2365 Cells Treated with COS-EGCG Conjugate
2.8. The Effect of COS-EGCG Conjugate on Bacterial Genomic DNA
2.9. Anti-Motility Testing
2.10. Scanning Electron Microscopic (SEM) Observation
2.11. Statistical Analysis
3. Results and Discussion
3.1. Antimicrobial Activity of COS and COS Conjugated with Different Phenolic Compounds against L. monocytogenes from Various Sources
3.2. Suppression of Extracellular Polysaccharide Production by COS-GECG Conjugate
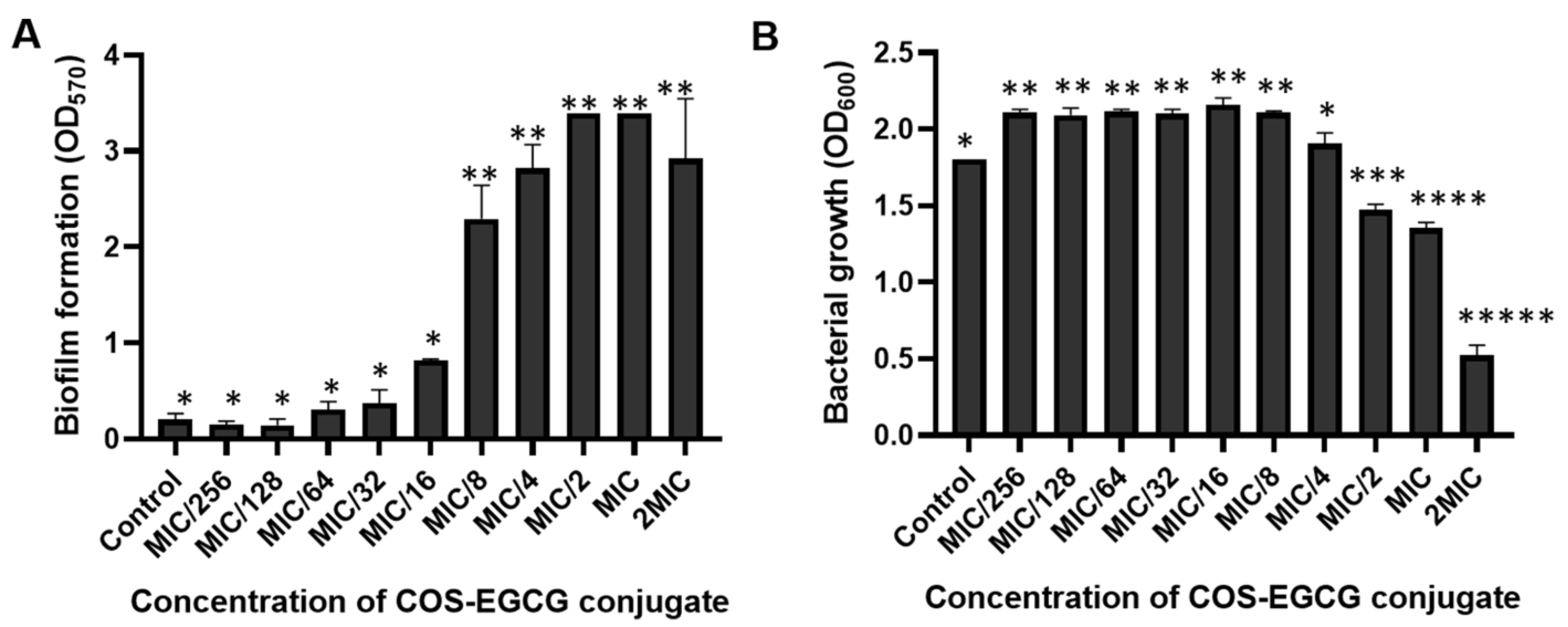
3.3. The Prevention and Inactivation of Biofilm by COS-EGCG Conjugate
3.4. Time-Kill Analysis
3.5. Protein Leakage
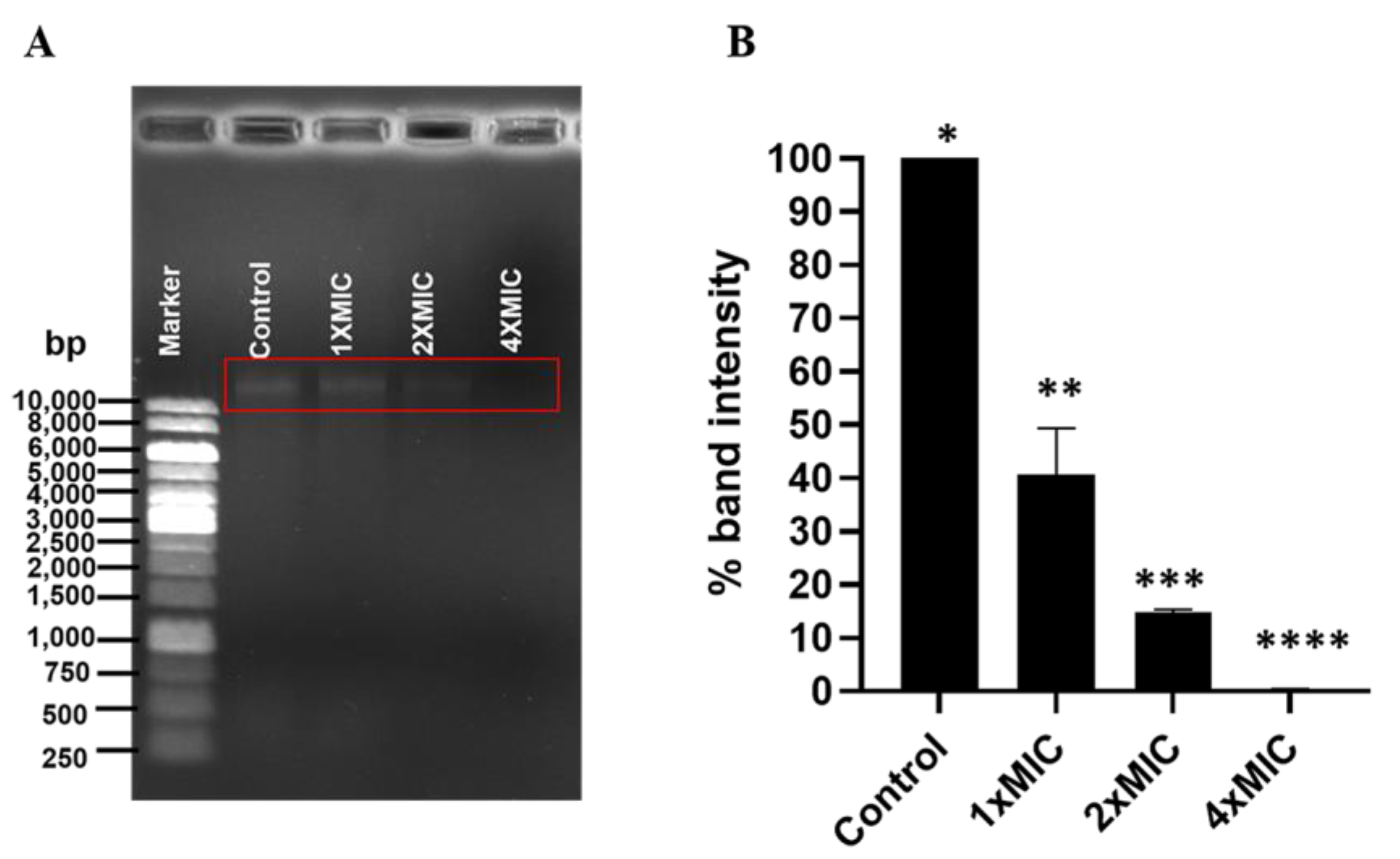
3.6. Effect of COS-EGCG Conjugate on DNA of L. monocytogenes
3.7. Effect of COS-EGCG Conjugate on L. monocytogenes Motility
3.8. Effect of COS-EGCG Conjugate on Morphological Changes of L. monocytogenes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dhama, K.; Karthik, K.; Tiwari, R.; Shabbir, M.Z.; Barbuddhe, S.; Malik, S.; Singh, R. Listeriosis in Animals, Its Public Health Significance (Food-Borne Zoonosis) and Advances in Diagnosis and Control: A Comprehensive Review. Vet. Q. 2015, 35, 211–235. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Moorhouse, E.; Monaghan, J.; Taylor, C.; Singleton, I. Sources and Survival of Listeria monocytogenes on Fresh, Leafy Produce. J. Appl. Microbiol. 2018, 125, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Kashimura, M.; Miya, S.; Kuramoto, S.; Koiso, H.; Kuda, T.; Kimura, B. Effect of Paired Antimicrobial Combinations on Listeria monocytogenes Growth Inhibition in Ready-to-Eat Seafood Products. Food Control 2012, 26, 397–400. [Google Scholar] [CrossRef]
- Lachmann, R.; Halbedel, S.; Lüth, S.; Holzer, A.; Adler, M.; Pietzka, A.; Dahouk, S.; Stark, K.; Flieger, A.; Kleta, S.; et al. Invasive Listeriosis Outbreaks and Salmon Products: A Genomic, Epidemiological Study. Emerg. Microbes Infect. 2022, 11, 1308–1315. [Google Scholar] [CrossRef]
- Beumer, R.R.; Hazeleger, W.C. Listeria monocytogenes: Diagnostic Problems. FEMS Immunol. Med. Microbiol. 2003, 35, 191–197. [Google Scholar] [CrossRef]
- Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Escámez, P.; Gironés, R.; Herman, L.; Nørrung, B.; Robertson, L.; et al. Listeria monocytogenes Contamination of Ready-to-eat Foods and the Risk for Human Health in the EU. EFSA J. 2018, 16, e05134. [Google Scholar] [CrossRef]
- Anand, S.P.; Sati, N. Artificial Preservatives and Their Harmful Effects: Looking toward Nature for Safer Alternatives. Int. J. Pharm. Sci. Res. 2013, 4, 2496–2501. [Google Scholar]
- Yarraguntla, S.R.; Kamala Kumari, P. Alternative to Artificial Preservatives. Syst. Rev. Pharm. 2019, 10, 99–102. [Google Scholar]
- Varun, T.; Senani, S.; Jayapal, N.; Chikkerur, J.; Roy, S.; Tekulapally, V.; Gautam, M.; Kumar, N. Extraction of Chitosan and Its Oligomers from Shrimp Shell Waste, Their Characterization and Antimicrobial Effect. Vet. World 2017, 10, 170–175. [Google Scholar] [CrossRef]
- Artan, M.; Karadeniz, F.; Karagozlu, M.; Kim, M.-M.; Kim, S.-K. Anti-HIV-1 Activity of Low Molecular Weight Sulfated Chitooligosaccharides. Carbohydr. Res. 2010, 345, 656–662. [Google Scholar] [CrossRef]
- Tsai, G.J.; Wu, Z.Y.; Su, W.H. Antibacterial Activity of a Chitooligosaccharide Mixture Prepared by Cellulase Digestion of Shrimp Chitosan and Its Application to Milk Preservation. J. Food Prot. 2000, 63, 747–752. [Google Scholar] [CrossRef]
- Mittal, A.; Singh, A.; Hong, H.; Benjakul, S. Chitooligosaccharides from Shrimp Shell Chitosan Prepared Using H2O2 or Ascorbic Acid/H2O2 Redox Pair Hydrolysis: Characteristics, Antioxidant and Antimicrobial Activities. Int. J. Food Sci. Technol. 2022. [Google Scholar] [CrossRef]
- Rakkhumkaew, N.; Pengsuk, C. Chitosan and Chitooligosaccharides from Shrimp Shell Waste: Characterization, Antimicrobial and Shelf Life Extension in Bread. Food Sci. Biotechnol. 2018, 27, 1201–1208. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Feng, J.; You, H.; Zhou, S.; Bai, Y.; He, J.; Cao, H.; Che, Q.; Guo, J.; Su, Z. The Microstructure, Antibacterial and Antitumor Activities of Chitosan Oligosaccharides and Derivatives. Mar. Drugs 2022, 20, 69. [Google Scholar] [CrossRef]
- Sirk, T.W.; Brown, E.F.; Sum, A.K.; Friedman, M. Molecular Dynamics Study on the Biophysical Interactions of Seven Green Tea Catechins with Lipid Bilayers of Cell Membranes. J. Agric. Food Chem. 2008, 56, 7750–7758. [Google Scholar] [CrossRef] [PubMed]
- Galdiero, S.; Falanga, A.; Cantisani, M.; Tarallo, R.; Elena Della Pepa, M.; D’Oriano, V.; Galdiero, M. Microbe-Host Interactions: Structure and Role of Gram-Negative Bacterial Porins. Curr. Protein Pept. Sci. 2012, 13, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Shimatani, K.; Ozawa, T.; Shigemune, N.; Tomiyama, D.; Yui, K.; Katsuki, M.; Ikeda, K.; Nonaka, A.; Miyamoto, T. Mechanism for the Antibacterial Action of Epigallocatechin Gallate (EGCg) on Bacillus subtilis. Biosci. Biotechnol. Biochem. 2015, 79, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Mittal, A.; Singh, A.; Zhang, B.; Visessanguan, W.; Benjakul, S. Chitooligosaccharide Conjugates Prepared Using Several Phenolic Compounds via Ascorbic Acid/H2O2 Free Radical Grafting: Characteristics, Antioxidant, Antidiabetic and Antimicrobial Activities. Foods 2022, 11, 920. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria, M45, 3rd ed.; CLSI Guideline M45; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Odedina, G.F.; Vongkamjan, K.; Voravuthikunchai, S.P. Potential Bio-Control Agent from Rhodomyrtus tomentosa against Listeria monocytogenes. Nutrients 2015, 7, 7451–7468. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Y.; Cao, J.; Jiang, H.; Yao, J.; Gong, G.; Chen, X.; Xu, W.; He, X. Antimicrobial Activity and Virulence Attenuation of Citral against the Fish Pathogen Vibrio alginolyticus. Aquaculture 2020, 515, 734578. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, Y.; Song, Q.; Wang, F.; Sun, L.; Liu, L.; Yang, X.; Yi, J.; Bao, Y.; Ma, H.; et al. Anti-Biofilm Activities from Bergenia Crassifolia Leaves against Streptococcus mutans. Front. Microbiol. 2017, 8, 1738. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Bian, L.; Zhu, Y.; Zhang, R.-R.; Shao, S.; Wu, Y.; Chen, Y.; Dang, Y.; Ding, Y.; Sun, H. Multifunctional Alkyl Ferulate Esters as Potential Food Additives: Antibacterial Activity and Mode of Action against Listeria monocytogenes and Its Application on American Sturgeon Caviar Preservation. Food Control 2018, 96, 390–402. [Google Scholar] [CrossRef]
- Avila-Novoa, M.G.; Navarrete-Sahagún, V.; González-Gómez, J.P.; Novoa-Valdovinos, C.; Guerrero-Medina, P.J.; García-Frutos, R.; Martínez-Chávez, L.; Martínez-Gonzáles, N.E.; Gutiérrez-Lomelí, M. Conditions of In Vitro Biofilm Formation by Serogroups of Listeria monocytogenes Isolated from Hass Avocados Sold at Markets in Mexico. Foods 2021, 10, 2097. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Xu, L.; Singh, A.K.; Qiu, X.; Liu, M.; Abuzeid, A.; El-Khateib, T.; Bhunia, A.K. Inactivation of Polymicrobial Biofilms of Foodborne Pathogens Using Epsilon Poly-L-Lysin Conjugated Chitosan Nanoparticles. Foods 2022, 11, 569. [Google Scholar] [CrossRef]
- Kim, G.; Xu, Y.; Zhang, J.; Sui, Z.; Corke, H. Antibacterial Activity and Multi-Targeting Mechanism of Dehydrocorydaline From Corydalis turtschaninovii Bess. Against Listeria monocytogenes. Front. Microbiol. 2022, 12, 3957. [Google Scholar] [CrossRef]
- Singh, A.; Benjakul, S.; Huda, N.; Xu, C.; Wu, P. Preparation and Characterization of Squid Pen Chitooligosaccharide-Epigallocatechin Gallate Conjugates and Their Antioxidant and Antimicrobial Activities. RSC Adv. 2020, 10, 33196–33204. [Google Scholar] [CrossRef]
- Singh, A.; Mittal, A.; Benjakul, S. Chitosan, Chitooligosaccharides and Their Polyphenol Conjugates: Preparation, Bioactivities, Functionalities and Applications in Food Systems. Food Rev. Int. 2021, 1–23. [Google Scholar] [CrossRef]
- Brandt, A.L.; Castillo, A.; Harris, K.B.; Keeton, J.T.; Hardin, M.D.; Taylor, T.M. Inhibition of Listeria monocytogenes by Food Antimicrobials Applied Singly and in Combination. J. Food Sci. 2010, 75, M557–M563. [Google Scholar] [CrossRef]
- Amankwah, S.; Abdella, K.; Kassa, T. Bacterial Biofilm Destruction: A Focused Review on the Recent Use of Phage-Based Strategies with Other Antibiofilm Agents. Nanotechnol. Sci. Appl. 2021, 14, 161. [Google Scholar] [CrossRef]
- Costa, O.Y.A.; Raaijmakers, J.M.; Kuramae, E.E. Microbial Extracellular Polymeric Substances: Ecological Function and Impact on Soil Aggregation. Front. Microbiol. 2018, 9, 1636. [Google Scholar] [CrossRef]
- Pinto, R.M.; Soares, F.A.; Reis, S.; Nunes, C.; Van Dijck, P. Innovative Strategies Toward the Disassembly of the EPS Matrix in Bacterial Biofilms. Front. Microbiol. 2020, 11, 952. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-H.; Köseoğlu, V.K.; Güvener, Z.T.; Myers-Morales, T.; Reed, J.M.; D’Orazio, S.E.F.; Miller, K.W.; Gomelsky, M. Cyclic Di-GMP-Dependent Signaling Pathways in the Pathogenic Firmicute Listeria monocytogenes. PLoS Pathog. 2014, 10, e1004301. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Jeong, M.-C.; Park, S.; Kim, S.-K.; Kim, Y.-M. Contribution of Chitooligosaccharides to Biofilm Formation, Antibiotics Resistance and Disinfectants Tolerance of Listeria monocytogenes. Microb. Pathog. 2019, 136, 103673. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.S.; Kim, Y.; Oh, S.; Jeon, W.M.; Frank, J.F.; Kim, S.H. Susceptibility of Listeria monocytogenes Biofilms and Planktonic Cultures to Hydrogen Peroxide in Food Processing Environments. Biosci. Biotechnol. Biochem. 2012, 76, 2008–2013. [Google Scholar] [CrossRef] [PubMed]
- Colagiorgi, A.; Bruini, I.; Ciccio, P.; Zanardi, E.; Ghidini, S.; Ianieri, A. Listeria monocytogenes Biofilms in the Wonderland of Food Industry. Pathogens 2017, 6, 41. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Liu, D.; Xu, L.; Tenguria, S.; Drolia, R.; Gallina, N.L.F.; Cox, A.D.; Koo, O.-K.; Bhunia, A.K. Biofilm-Isolated Listeria Monocytogenes Exhibits Reduced Systemic Dissemination at the Early (12–24 h) Stage of Infection in a Mouse Model. Npj Biofilms Microbiomes 2021, 7, 18. [Google Scholar] [CrossRef]
- Margolin, W. Bacterial Cell Wall; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1–11. ISBN 978-0-470-01590-2. [Google Scholar]
- Russell, A.D. Bacterial Outer Membrane and Cell Wall Penetration and Cell Destruction by Polluting Chemical Agents and Physical Conditions. Sci. Prog. 2003, 86, 283–312. [Google Scholar] [CrossRef]
- Fang, M.; Wang, R.; Agyekumwaa, A.K.; Yu, Y.; Xiao, X. Antibacterial Effect of Phenyllactic Acid against Vibrio parahaemolyticus and Its Application on Raw Salmon Fillets. LWT 2022, 154, 112586. [Google Scholar] [CrossRef]
- Kitichalermkiat, A.; Katsuki, M.; Sato, J.; Sonoda, T.; Masuda, Y.; Honjoh, K.; Miyamoto, T. Effect of Epigallocatechin Gallate on Gene Expression of Staphylococcus aureus. J. Glob. Antimicrob. Resist. 2020, 22, 854–859. [Google Scholar] [CrossRef]
- Guan, Z.; Feng, Q. Chitosan and Chitooligosaccharide: The Promising Non-Plant-Derived Prebiotics with Multiple Biological Activities. Int. J. Mol. Sci. 2022, 23, 6761. [Google Scholar] [CrossRef]
- Şenel, B. In Vitro Preliminary Studies of Chitooligosaccharide Coated Nanostructered Lipidic Nanoparticles for Efficient Gene Delivery. Marmara Pharm. J. 2019, 23, 671–681. [Google Scholar] [CrossRef]
- Bai, X.; Nakatsu, C.H.; Bhunia, A.K. Bacterial Biofilms and Their Implications in Pathogenesis and Food Safety. Foods 2021, 10, 2117. [Google Scholar] [CrossRef] [PubMed]
- Kearns, D.B. A Field Guide to Bacterial Swarming Motility. Nat. Rev. Microbiol. 2010, 8, 634–644. [Google Scholar] [CrossRef] [PubMed]
- Lemon Katherine, P.; Higgins Darren, E. Kolter Roberto Flagellar Motility Is Critical for Listeria monocytogenes Biofilm Formation. J. Bacteriol. 2007, 189, 4418–4424. [Google Scholar] [CrossRef]
- Li, R.; Du, W.; Yang, J.; Liu, Z.; Yousef, A.E. Control of Listeria monocytogenes Biofilm by Paenibacterin, a Natural Antimicrobial Lipopeptide. Food Control 2018, 84, 529–535. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, L.; Han, J.; Dong, P.; Luo, X.; Zhang, Y.; Zhu, L. Inhibition of Biofilm Formation and Related Gene Expression of Listeria monocytogenes in Response to Four Natural Antimicrobial Compounds and Sodium Hypochlorite. Front. Microbiol. 2021, 11, 617473. [Google Scholar] [CrossRef]
- Gao, T.; Ye, F.; Tan, Y.; Peng, M.; Yuan, F.; Liu, Z.; Zhou, D.; Yang, K.; Liu, W.; Guo, R.; et al. Metabolomics and Proteomics Analyses Revealed Mechanistic Insights on the Antimicrobial Activity of Epigallocatechin Gallate against Streptococcus suis. Front. Cell. Infect. Microbiol. 2022, 12, 1358. [Google Scholar] [CrossRef]
- Poolman, B.; Glaasker, E. Regulation of Compatible Solute Accumulation in Bacteria. Mol. Microbiol. 1998, 29, 397–407. [Google Scholar] [CrossRef]
- Garde, S.; Chodisetti, P.K.; Reddy, M. Peptidoglycan: Structure, Synthesis, and Regulation. EcoSal Plus 2021, 9. [Google Scholar] [CrossRef]
- Matica, M.A.; Aachmann, F.L.; Tøndervik, A.; Sletta, H.; Ostafe, V. Chitosan as a Wound Dressing Starting Material: Antimicrobial Properties and Mode of Action. Int. J. Mol. Sci. 2019, 20, 5889. [Google Scholar] [CrossRef]
- Shimamura, T.; Zhao, W.; Hu, Z.-Q. Mechanism of Action and Potential for Use of Tea Catechin as an Antiinfective Agent. Anti-Infect. Agents Med. Chem. Former. Curr. Med. Chem.-Anti-Infect. Agents 2007, 6, 57–62. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substances | MIC (µg/mL) | MBC (µg/mL) | MBC/MIC Ratio |
|---|---|---|---|
| COS | 2048 | 2048 | 1 |
| EGCG | 1024 | 2048 | 2 |
| COS-CAT | 256 | 2048 | 8 |
| COS-GAL | 2048 | 2048 | 1 |
| COS-CAF | 2048 | 2048 | 1 |
| COS-FER | 2048 | 2048 | 1 |
| COS-EGCG | 1024 | 1024 | 1 |
| L. monocytogenes Strains | MIC (µg/mL) | MBC (µg/mL) | MBC/MIC Ratio |
|---|---|---|---|
| LM2 | 1024 | 2048 | 2 |
| LM3 | 1024 | 2048 | 2 |
| LM9 | 1024 | 2048 | 2 |
| LM13 | 1024 | 2048 | 2 |
| LM14 | 128 | 2048 | 16 |
| EN5203 | 1024 | 2048 | 2 |
| EN5326 | 1024 | 2048 | 2 |
| EN5402 | 1024 | 2048 | 2 |
| EN5404 | 1024 | 2048 | 2 |
| EN5642 | 1024 | 2048 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buatong, J.; Mittal, A.; Mittraparp-arthorn, P.; Palamae, S.; Saetang, J.; Benjakul, S. Bactericidal Action of Shrimp Shell Chitooligosaccharide Conjugated with Epigallocatechin Gallate (COS-EGCG) against Listeria monocytogenes. Foods 2023, 12, 634. https://doi.org/10.3390/foods12030634
Buatong J, Mittal A, Mittraparp-arthorn P, Palamae S, Saetang J, Benjakul S. Bactericidal Action of Shrimp Shell Chitooligosaccharide Conjugated with Epigallocatechin Gallate (COS-EGCG) against Listeria monocytogenes. Foods. 2023; 12(3):634. https://doi.org/10.3390/foods12030634
Chicago/Turabian StyleBuatong, Jirayu, Ajay Mittal, Pimonsri Mittraparp-arthorn, Suriya Palamae, Jirakrit Saetang, and Soottawat Benjakul. 2023. "Bactericidal Action of Shrimp Shell Chitooligosaccharide Conjugated with Epigallocatechin Gallate (COS-EGCG) against Listeria monocytogenes" Foods 12, no. 3: 634. https://doi.org/10.3390/foods12030634
APA StyleBuatong, J., Mittal, A., Mittraparp-arthorn, P., Palamae, S., Saetang, J., & Benjakul, S. (2023). Bactericidal Action of Shrimp Shell Chitooligosaccharide Conjugated with Epigallocatechin Gallate (COS-EGCG) against Listeria monocytogenes. Foods, 12(3), 634. https://doi.org/10.3390/foods12030634