
Computer-Aided Screening and Revealing Action Mechanism of Green Tea Polyphenols Intervention in Alzheimer’s Disease


Abstract
1. Introduction
2. Materials and Methods
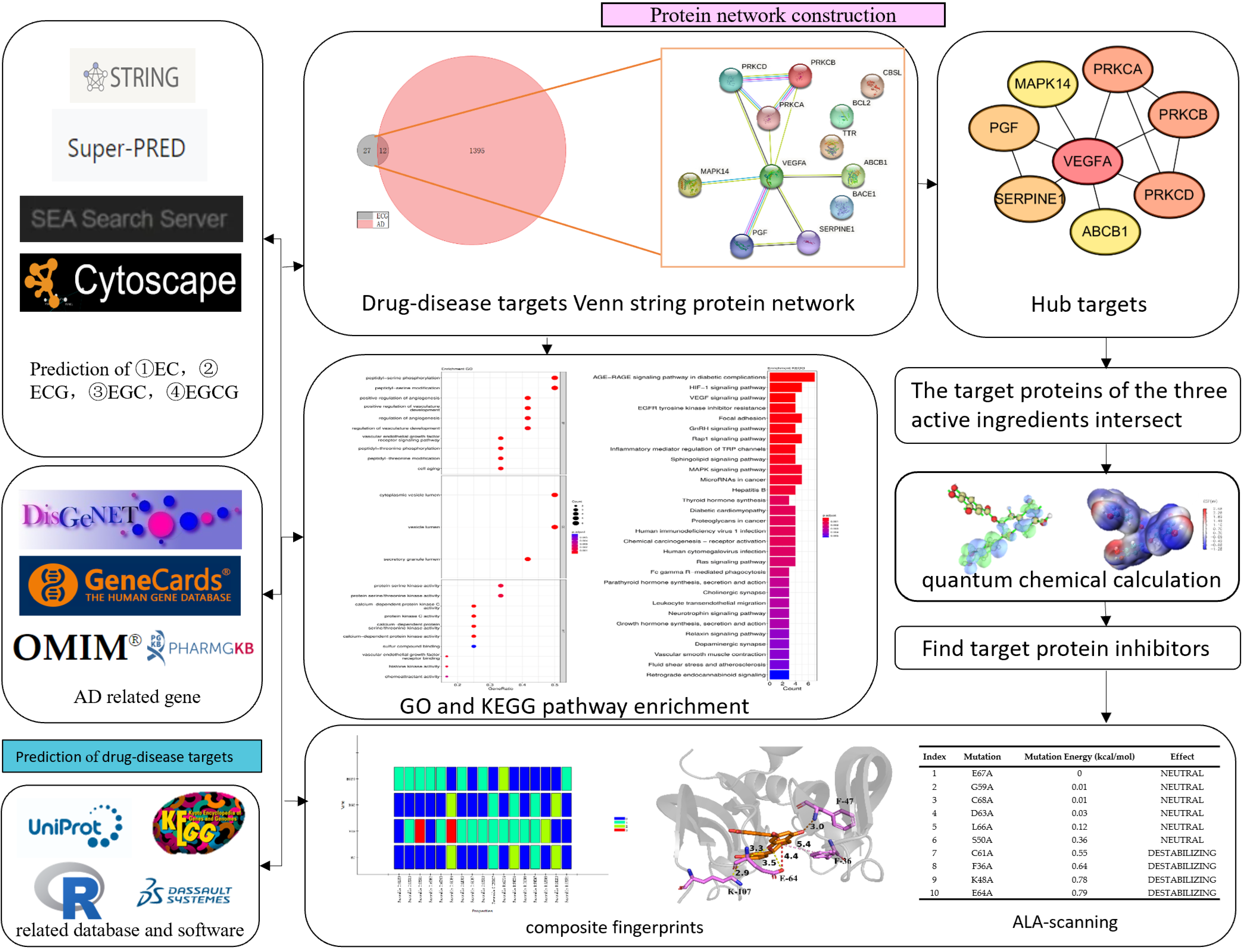
2.1. Prediction of AD-Related Targets and Tea Polyphenols Targets
2.2. Construction of the Protein–Protein Interaction (PPI) Network
2.3. GO and KEGG Enrichment Analysis
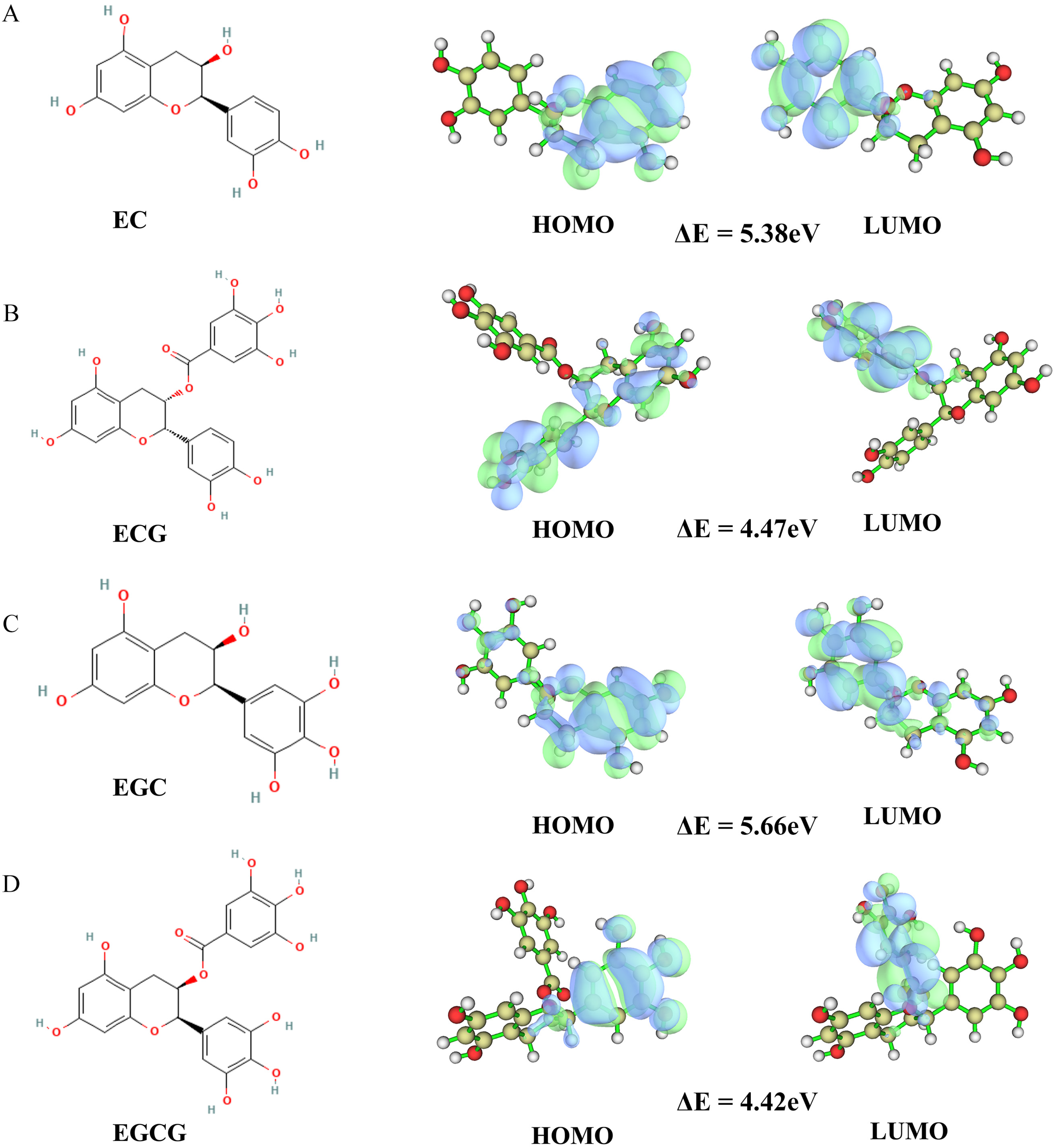
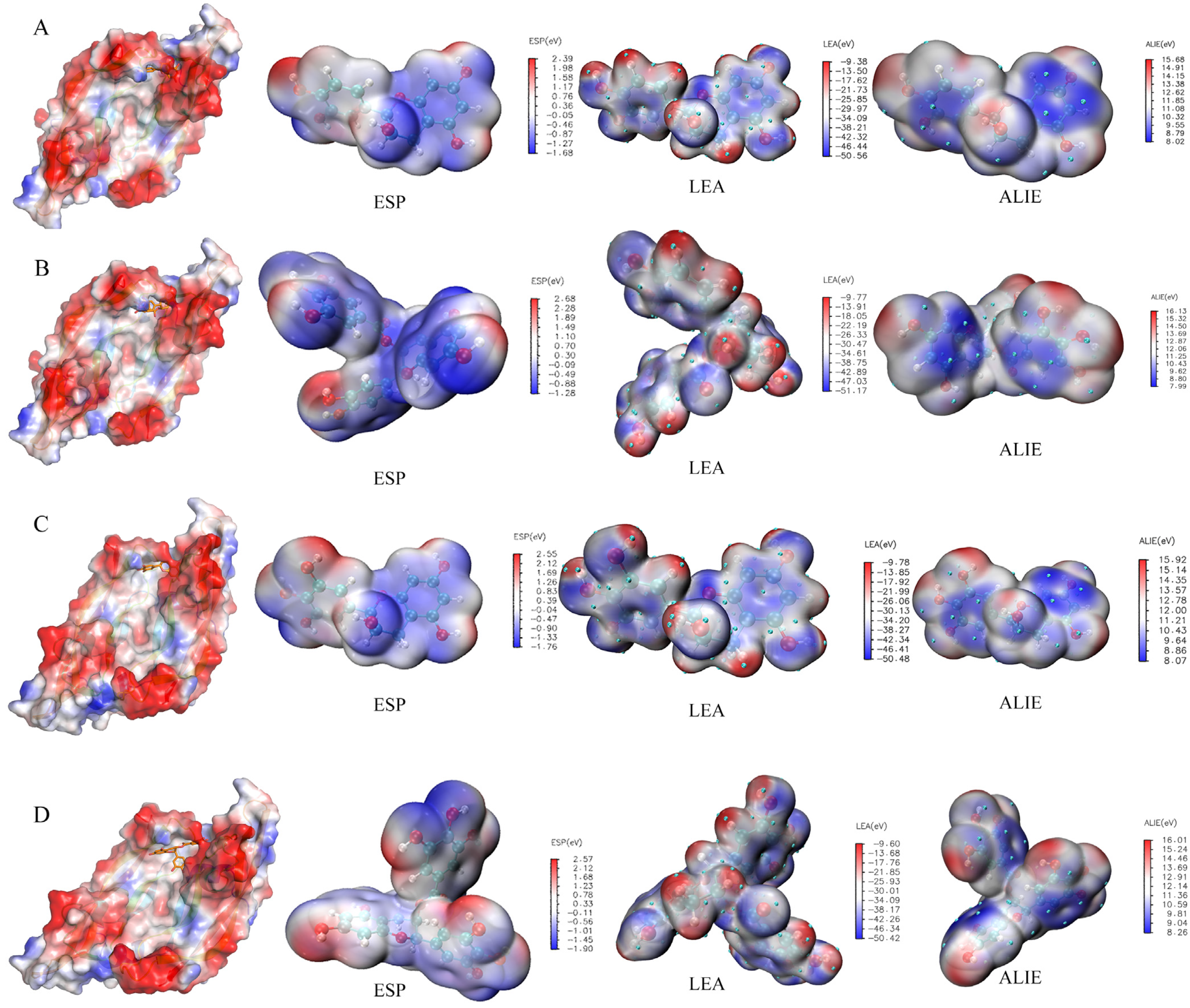
2.4. Quantum Chemical Calculation
2.5. Complex Fingerprint Analysis
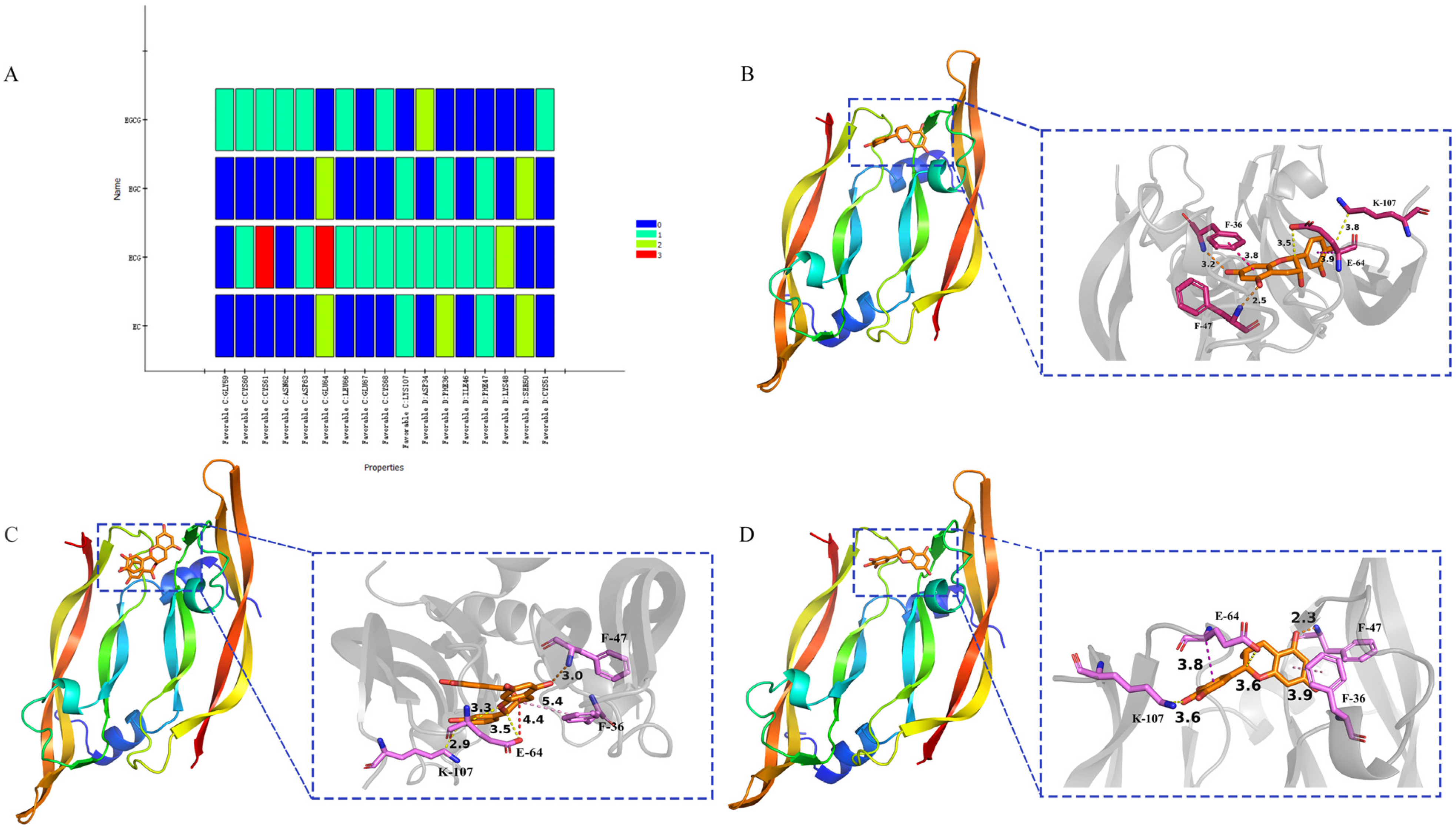
2.6. Alanine Scanning
3. Results
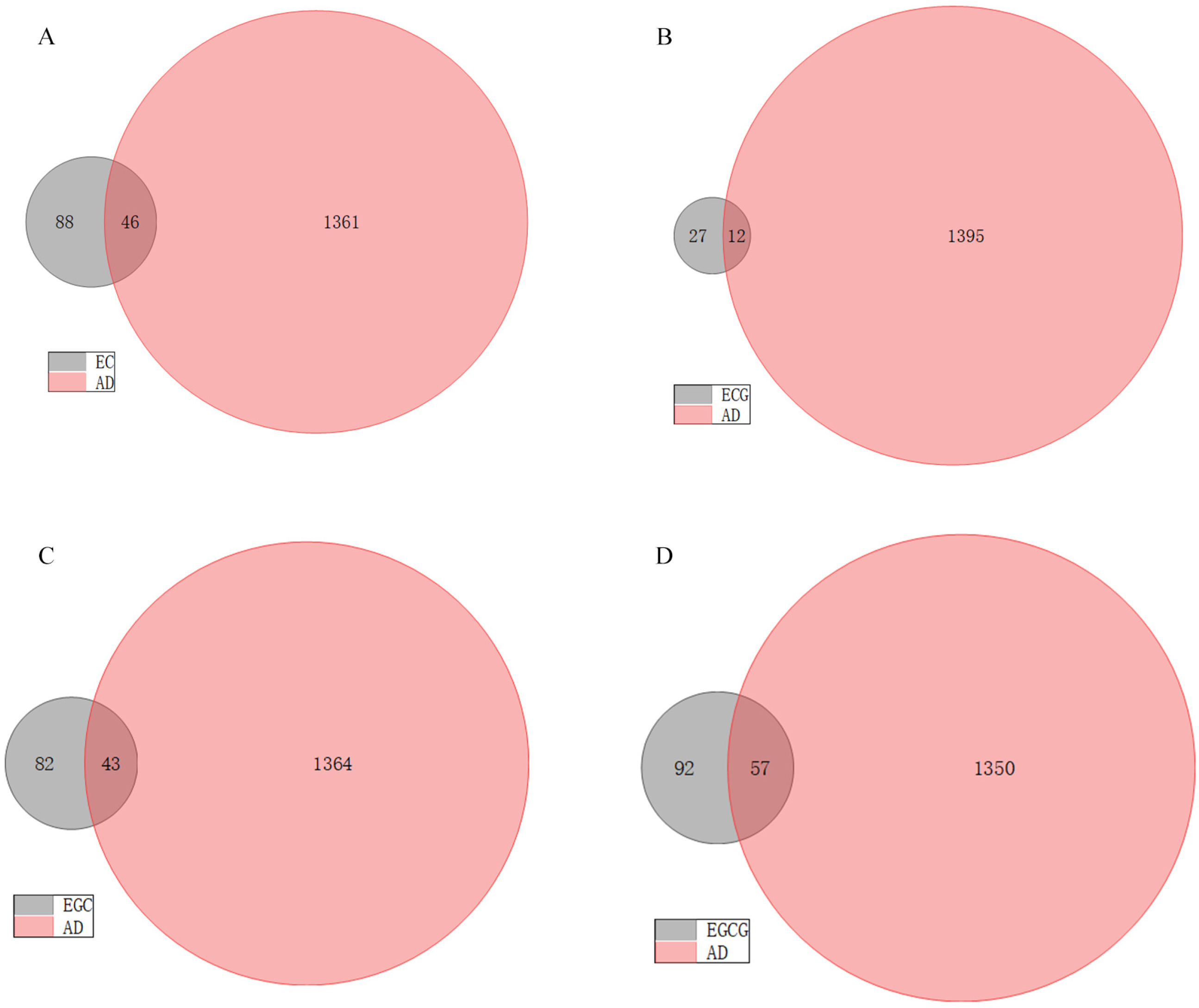
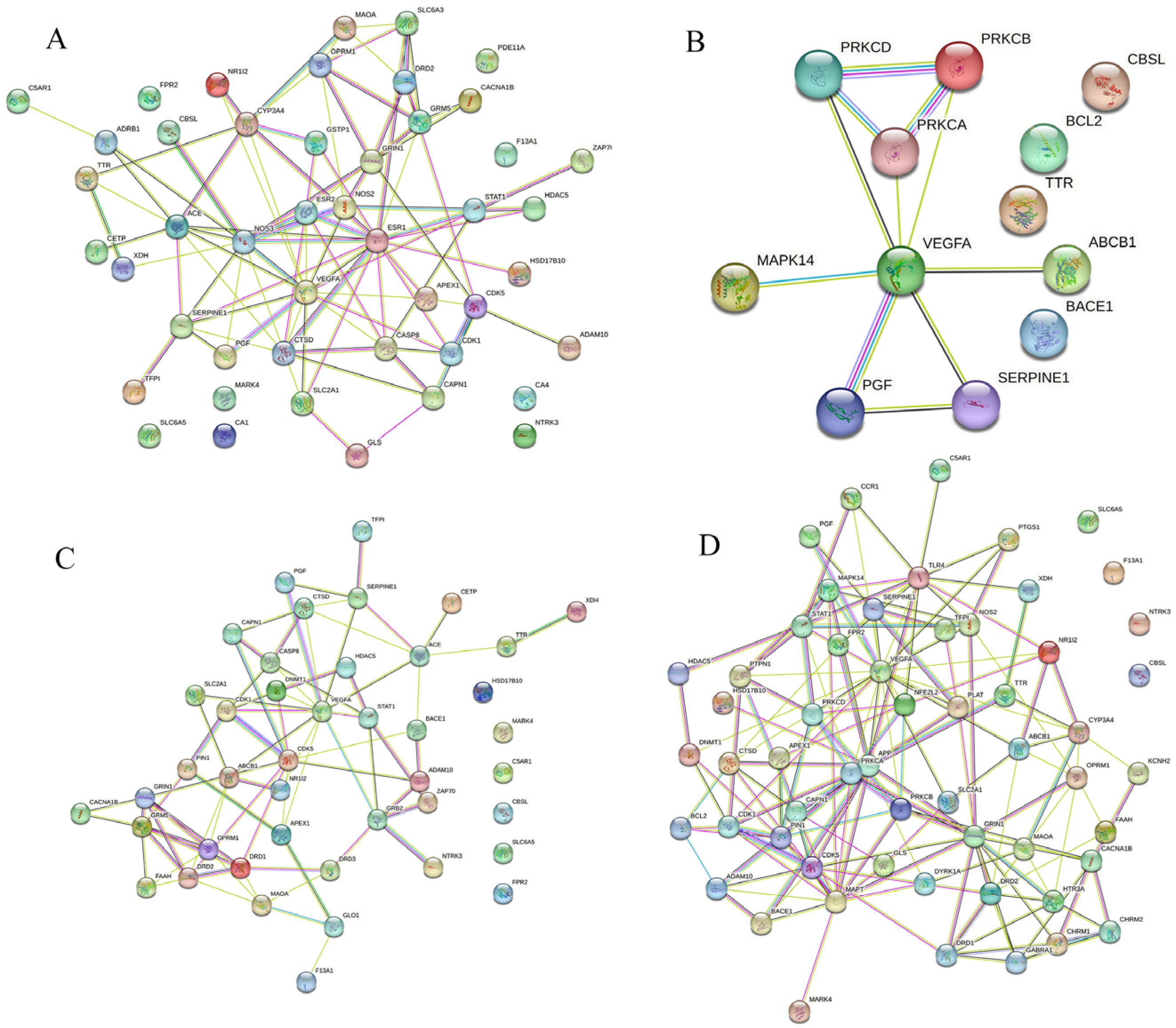
3.1. Potential Targets of AD and Tea Polyphenols
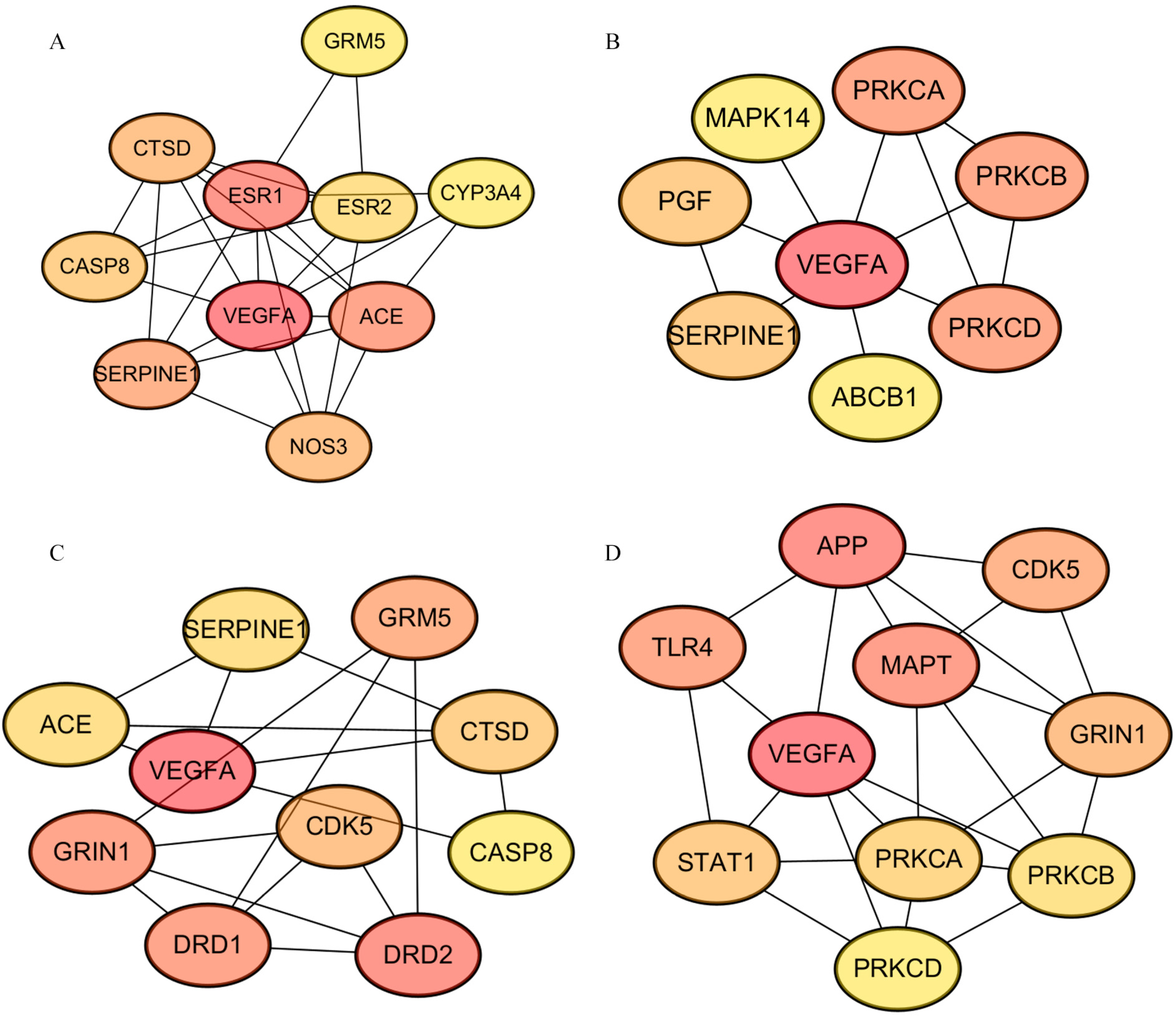
3.2. Hub Targets Collection of Tea Polyphenols Anti-AD
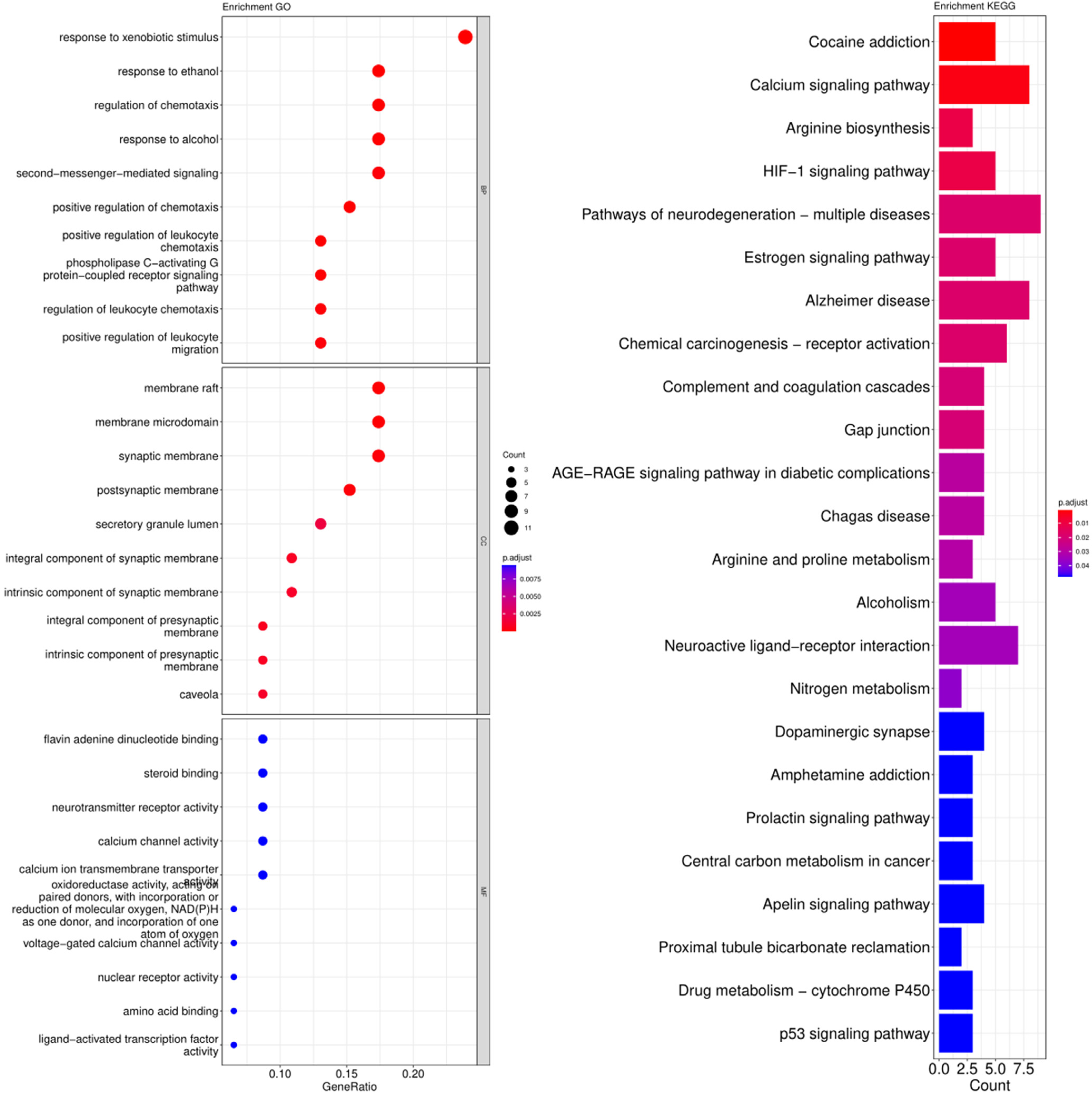
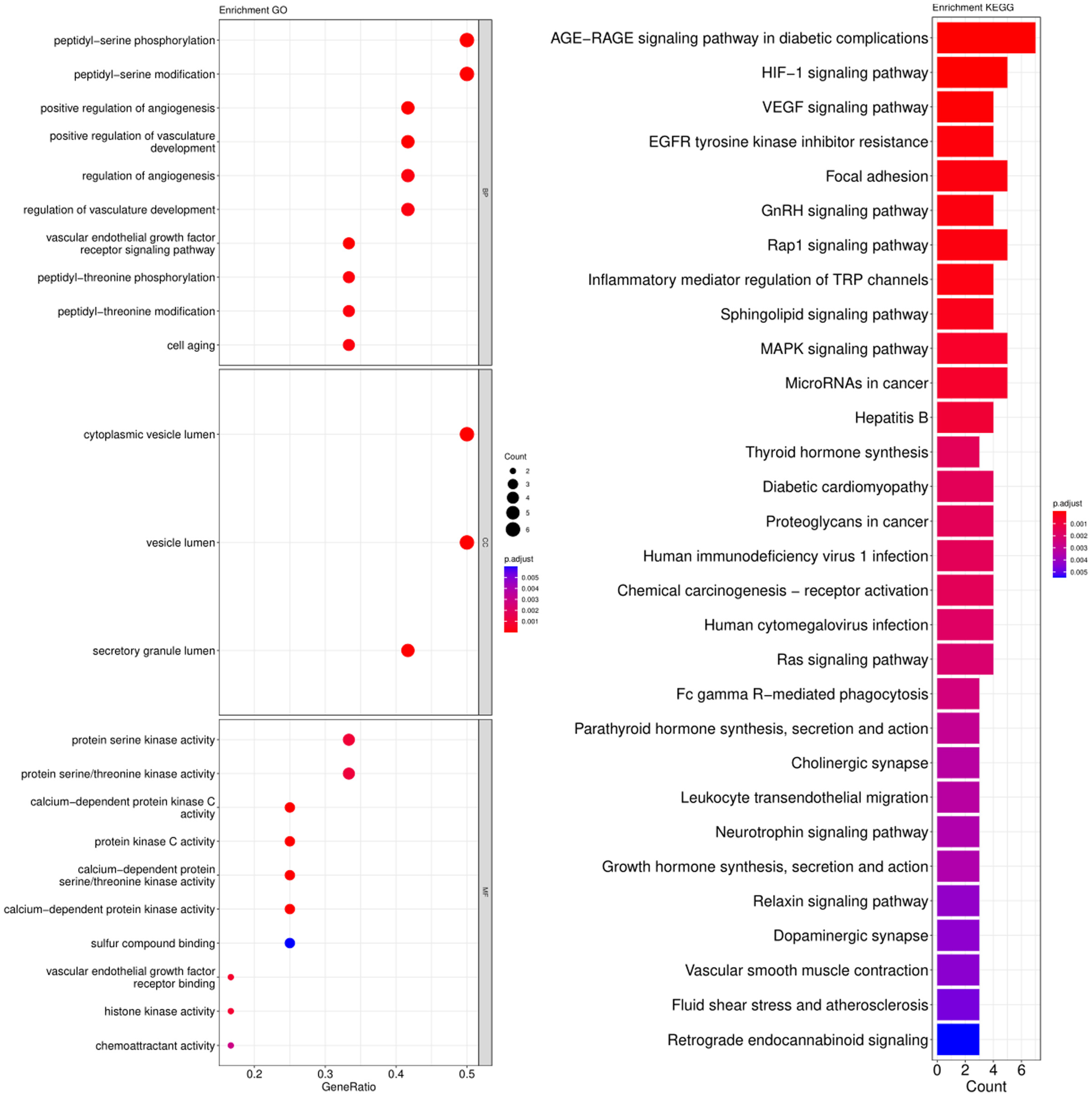
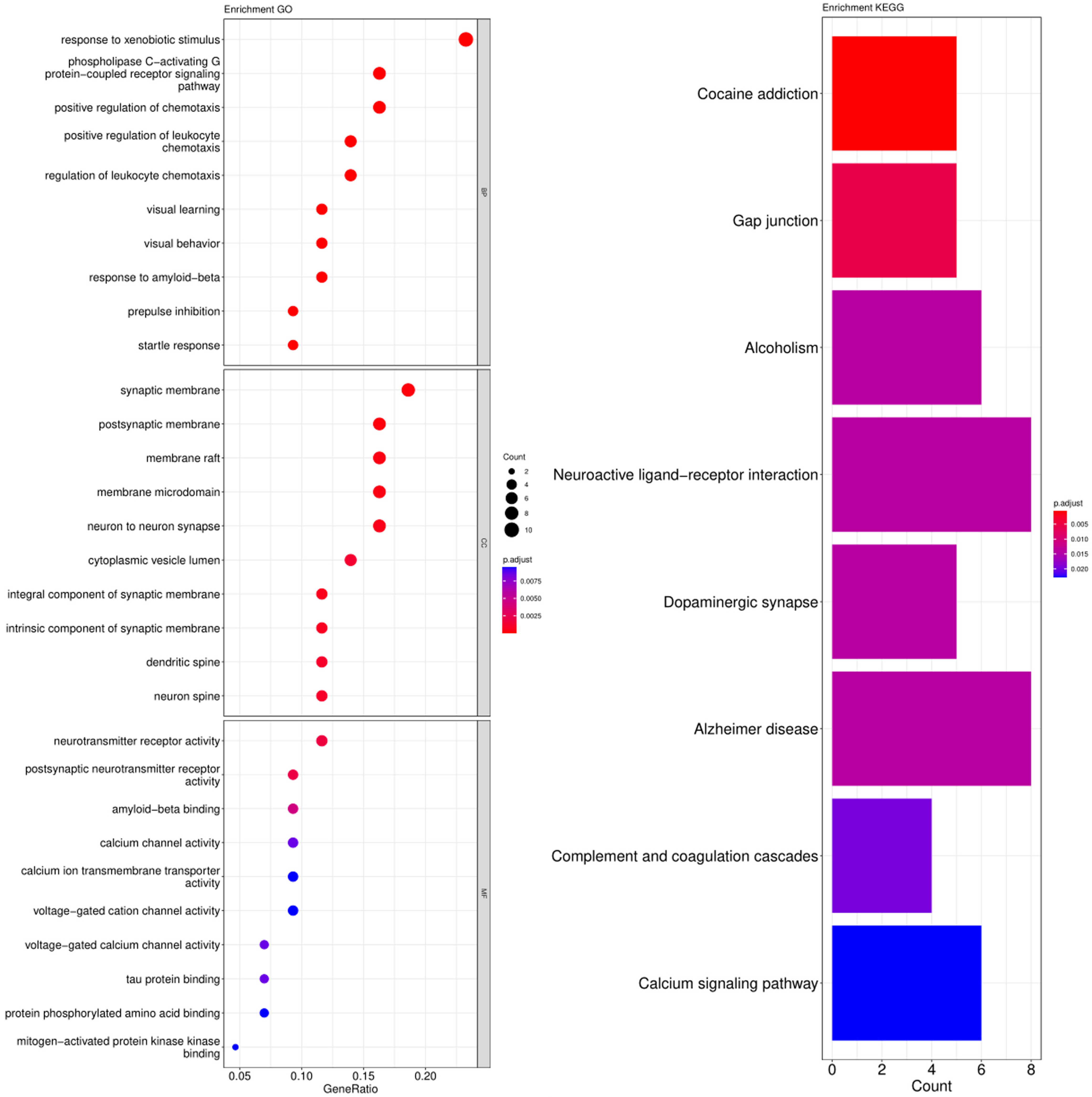
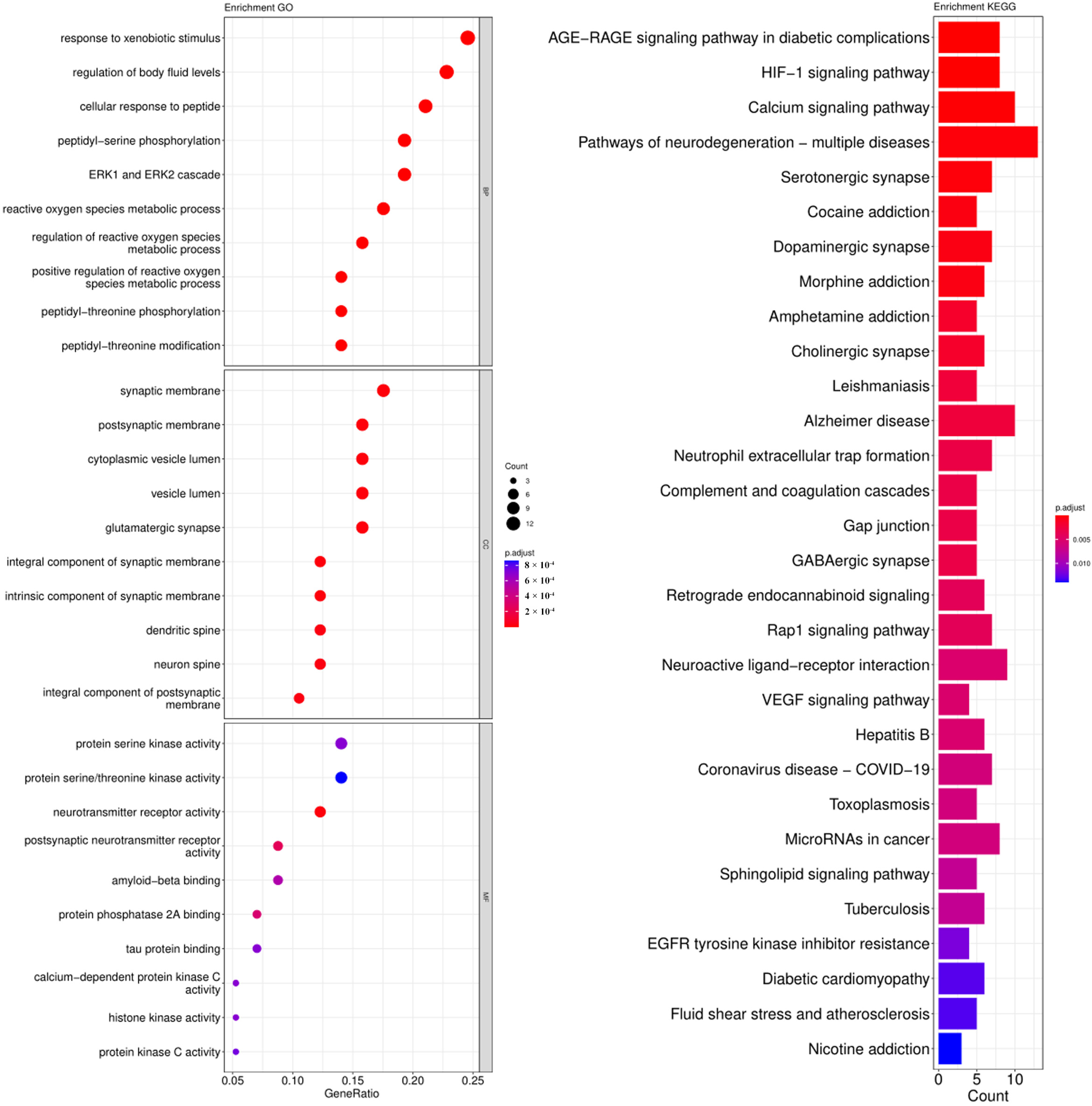
3.3. GO and KEGG Pathway Enrichment Analysis
3.4. Quantum Chemical Calculation of Four Tea Polyphenols
3.5. Binding of Four Tea Polyphenols to VEGFA
3.6. Alanine Scanning of the Binding Sites
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jeremic, D.; Jimenez-Diaz, L.; Navarro-Lopez, J.D. Past, present and future of therapeutic strategies against amyloid-beta peptides in Alzheimer’s disease: A systematic review. Ageing Res. Rev. 2021, 72, 101496. [Google Scholar] [CrossRef]
- Pleen, J.; Townley, R. Alzheimer’s disease clinical trial update 2019–2021. J. Neurol. 2022, 269, 1038–1051. [Google Scholar] [CrossRef] [PubMed]
- Tatulian, S.A. Challenges and hopes for Alzheimer’s disease. Drug Discov. Today 2022, 27, 1027–1043. [Google Scholar] [CrossRef]
- Janoutova, J.; Machaczka, O.; Zatloukalova, A.; Janout, V. Is Alzheimer’s disease a type 3 diabetes? A review. Cent. Eur. J. Public Health 2022, 30, 139–143. [Google Scholar] [CrossRef]
- Khan, S.; Barve, K.H.; Kumar, M.S. Recent Advancements in Pathogenesis, Diagnostics and Treatment of Alzheimer’s Disease. Curr. Neuropharmacol. 2020, 18, 1106–1125. [Google Scholar] [CrossRef]
- Pakdin, M.; Toutounchian, S.; Namazi, S.; Arabpour, Z.; Pouladi, A.; Afsahi, S.; Poudineh, M.; Nasab, M.M.M.; Yaghoobpoor, S.; Deravi, N. Type 2 Diabetes Mellitus and Alzheimer’s Disease: A Review of the Potential Links. Curr. Diabetes Rev. 2022, 18, e051121197760. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lu, J.; Zhao, X.; Qin, R.; Song, K.; Xu, Y.; Zhang, J.; Chen, Y. Alzheimer’s disease in elderly COVID-19 patients: Potential mechanisms and preventive measures. Neurol. Sci. 2021, 42, 4913–4920. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Singh, H.; Singh, B.; Kumar, N.; Rajput, A.; Sidhu, D.; Kaur, A.; Arora, S.; Kaur, S. A Comprehensive Review on Medicinal Herbs and Novel Formulations for the Prevention of Alzheimer’s Disease. Curr. Drug Deliv. 2022, 19, 212–228. [Google Scholar] [CrossRef]
- Lu, L.; Yu, X.; Cai, Y.; Sun, M.; Yang, H. Application of CRISPR/Cas9 in Alzheimer’s Disease. Front. Neurosci. 2021, 15, 803894. [Google Scholar] [CrossRef]
- Ouyang, F.; Jiang, Z.; Chen, X.; Chen, Y.; Wei, J.; Xing, S.; Zhang, J.; Fan, Y.; Zeng, J. Is Cerebral Amyloid-beta Deposition Related to Post-stroke Cognitive Impairment? Transl. Stroke Res. 2021, 12, 946–957. [Google Scholar] [CrossRef]
- Shi, M.; Chu, F.; Zhu, F.; Zhu, J. Impact of Anti-amyloid-beta Monoclonal Antibodies on the Pathology and Clinical Profile of Alzheimer’s Disease: A Focus on Aducanumab and Lecanemab. Front. Aging Neurosci. 2022, 14, 870517. [Google Scholar] [CrossRef]
- Chen, D.; Liu, X.; Chen, Y.; Lin, H. Amyloid peptides with antimicrobial and/or microbial agglutination activity. Appl. Microbiol. Biotechnol. 2022, 106, 7711–7720. [Google Scholar] [CrossRef] [PubMed]
- Roda, A.R.; Serra-Mir, G.; Montoliu-Gaya, L.; Tiessler, L.; Villegas, S. Amyloid-beta peptide and tau protein crosstalk in Alzheimer’s disease. Neural. Regen. Res. 2022, 17, 1666–1674. [Google Scholar] [CrossRef] [PubMed]
- Soto-Ospina, A.; Araque Marin, P.; Bedoya, G.J.; Villegas Lanau, A. Structural Predictive Model of Presenilin-2 Protein and Analysis of Structural Effects of Familial Alzheimer’s Disease Mutations. Biochem. Res. Int. 2021, 2021, 9542038. [Google Scholar] [CrossRef] [PubMed]
- Knorz, A.L.; Quante, A. Alzheimer’s Disease: Efficacy of Mono- and Combination Therapy. A Systematic Review. J. Geriatr. Psychiatry Neurol. 2022, 35, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Poudel, P.; Park, S. Recent Advances in the Treatment of Alzheimer’s Disease Using Nanoparticle-Based Drug Delivery Systems. Pharmaceutics 2022, 14, 835. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, L.; Li, Z.; Zhang, P.; Song, H.; Yao, D.A.; Cao, J.; Zhang, J.J. Green tea improves cognitive function through reducing AD-pathology and improving anti-oxidative stress capacity in Chinese middle-aged and elderly people. Front. Aging Neurosci. 2022, 14, 919766. [Google Scholar] [CrossRef]
- Amirpour, M.; Mirshekar, M.A.; Sedaghat, G.; Montazerifar, F.; Shourestani, S.; Arabmoazzen, S.; Naghizadeh, M. The effects of green tea on cognitive impairments in the rat model of Alzheimer’s disease: Protection against inflammatory and oxidative damage. Nutr. Neurosci. 2022, 25, 2659–2667. [Google Scholar] [CrossRef]
- Hou, Z.W.; Chen, C.H.; Ke, J.P.; Zhang, Y.Y.; Qi, Y.; Liu, S.Y.; Yang, Z.; Ning, J.M.; Bao, G.H. alpha-Glucosidase Inhibitory Activities and the Interaction Mechanism of Novel Spiro-Flavoalkaloids from YingDe Green Tea. J. Agric. Food Chem. 2022, 70, 136–148. [Google Scholar] [CrossRef]
- Bastianetto, S.; Yao, Z.X.; Papadopoulos, V.; Quirion, R. Neuroprotective effects of green and black teas and their catechin gallate esters against beta-amyloid-induced toxicity. Eur. J. Neurosci. 2006, 23, 55–64. [Google Scholar] [CrossRef]
- Wobst, H.J.; Sharma, A.; Diamond, M.I.; Wanker, E.E.; Bieschke, J. The green tea polyphenol (-)-epigallocatechin gallate prevents the aggregation of tau protein into toxic oligomers at substoichiometric ratios. FEBS Lett. 2015, 589, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Gour, S.; Peter, O.S.; Gandhi, S.; Goyal, P.; Pandey, J.; Harsolia, R.S.; Yadav, J.K. Effect of Green Tea Polyphenol Epigallocatechin-3-gallate on the Aggregation of alphaA(66-80) Peptide, a Major Fragment of alphaA-crystallin Involved in Cataract Development. Curr. Eye Res. 2017, 42, 1368–1377. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Yang, Y.; Zhu, S.; Lu, Y.; Zhu, L.; Wang, Y.; Wang, X. Inhibition of Abeta aggregates in Alzheimer’s disease by epigallocatechin and epicatechin-3-gallate from green tea. Bioorg. Chem. 2020, 105, 104382. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Guo, Y.; Wu, Z.; Zhao, S.; Zhang, Z.; Zheng, F.; Sun, L.; Hao, Z.; Xu, C.; Wang, T.; et al. Epicatechin gallate prevents the de novo synthesis of fatty acid and the migration of prostate cancer cells. Acta Biochim. Biophys. Sin. 2021, 53, 1662–1669. [Google Scholar] [CrossRef]
- Yong, H.; Hu, H.; Wang, Z.; Yun, D.; Kan, J.; Liu, J. Structure, stability and antioxidant activity of dialdehyde starch grafted with epicatechin, epicatechin gallate, epigallocatechin and epigallocatechin gallate. J. Sci. Food Agric. 2022, 102, 6373–6386. [Google Scholar] [CrossRef]
- Chen, W.; Zhou, C.; Chen, J.; Wang, M.; Zhou, F.; Wang, C.; Zhang, X.; Zhou, W. Design and synthesis of epigallocatechin (EGC) analogs selective to inhibit alpha-amylase over alpha-glucosidases via the incorporation of caffeine acid and its derivatives. Bioorg. Chem. 2022, 119, 105515. [Google Scholar] [CrossRef] [PubMed]
- da Silva-Junior, E.F.; Silva, L.R. Multi-target Approaches of Epigallocatechin-3-O-gallate (EGCG) and its Derivatives against Influenza Viruses. Curr. Top Med. Chem. 2022, 22, 1485–1500. [Google Scholar] [CrossRef]
- Butterfield, D.; Castegna, A.; Pocernich, C.; Drake, J.; Scapagnini, G.; Calabrese, V. Nutritional approaches to combat oxidative stress in Alzheimer’s disease. J. Nutr. Biochem. 2002, 13, 444. [Google Scholar] [CrossRef]
- Higdon, J.V.; Frei, B. Tea catechins and polyphenols: Health effects, metabolism, and antioxidant functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef]
- Weinreb, O.; Mandel, S.; Amit, T.; Youdim, M.B. Neurological mechanisms of green tea polyphenols in Alzheimer’s and Parkinson’s diseases. J. Nutr. Biochem. 2004, 15, 506–516. [Google Scholar] [CrossRef]
- Surguchov, A.; Bernal, L.; Surguchev, A.A. Phytochemicals as Regulators of Genes Involved in Synucleinopathies. Biomolecules 2021, 11, 624. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Yoshiike, Y.; Takashima, A.; Hasegawa, K.; Naiki, H.; Yamada, M. Potent anti-amyloidogenic and fibril-destabilizing effects of polyphenols in vitro: Implications for the prevention and therapeutics of Alzheimer’s disease. J. Neurochem. 2003, 87, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.S.; Gong, X.F.; Feng, J.L.; Li, H.S.; Li, X.; Deng, S.; Ren, P.Z.; Wang, J.M.; Lv, M.S.; Jin, R.F.; et al. Study on the Mechanism of Jiawei Shengjiang Powder in Improving Male Asthma-Induced Asthenospermia Based on Network Pharmacology and Bioinformatics. Drug Des. Devel. Ther. 2021, 15, 1245–1259. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Fumery, M.; Dulai, P.S.; Prokop, L.J.; Sandborn, W.J.; Murad, M.H.; Singh, S. Comparative efficacy and tolerability of pharmacological agents for management of mild to moderate ulcerative colitis: A systematic review and network meta-analyses. Lancet Gastroenterol. Hepatol. 2018, 3, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Khan, G.J.; Shang, L.J.; Peng, H.; Hu, W.C.; Zhang, J.Y.; Hua, J.; Cassandra, A.; Rashed, M.M.A.; Zhai, K.F. Computational pharmacology and bioinformatics to explore the potential mechanism of Schisandra against atherosclerosis. Food Chem. Toxicol. 2021, 150, 112058. [Google Scholar] [CrossRef]
- Pinero, J.; Ramirez-Anguita, J.M.; Sauch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1.30.1–1.30.33. [Google Scholar] [CrossRef]
- Whirl-Carrillo, M.; Huddart, R.; Gong, L.; Sangkuhl, K.; Thorn, C.F.; Whaley, R.; Klein, T.E. An Evidence-Based Framework for Evaluating Pharmacogenomics Knowledge for Personalized Medicine. Clin. Pharmacol. Ther. 2021, 110, 563–572. [Google Scholar] [CrossRef]
- UniProt, C. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Keiser, M.J.; Roth, B.L.; Armbruster, B.N.; Ernsberger, P.; Irwin, J.J.; Shoichet, B.K. Relating protein pharmacology by ligand chemistry. Nat. Biotechnol. 2007, 25, 197–206. [Google Scholar] [CrossRef]
- Nickel, J.; Gohlke, B.O.; Erehman, J.; Banerjee, P.; Rong, W.W.; Goede, A.; Dunkel, M.; Preissner, R. SuperPred: Update on drug classification and target prediction. Nucleic Acids Res. 2014, 42, W26–W31. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- The Gene Ontology Consortium. The Gene Ontology resource: Enriching a GOld mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.; Ketabi, S.; Hasheminasab, G. QSAR study of antituberculosis activity of oxadiazole derivatives using DFT calculations. J. Recept Signal Transduct Res. 2022, 42, 503–511. [Google Scholar] [CrossRef]
- Lu, T.; Chen, F. Multiwfn: A multifunctional wavefunction analyzer. J. Comput. Chem. 2012, 33, 580–592. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Brooks, B.R.; Brooks, C.L., 3rd; Mackerell, A.D., Jr.; Nilsson, L.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.N.; Head, M.S.; Kulkarni, A.; LaLonde, J.M. Validation studies of the site-directed docking program LibDock. J. Chem. Inf. Model 2007, 47, 2159–2171. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Liu, Y.; Zheng, P.; Zhang, T.; Ye, X.; Liu, M.; Huang, M.; Wan, Q.; Zhang, J. Based on UPLC-Q-TOF-MS/MS, Systematic Network Pharmacology, and Molecular Docking to Explore the Potential Mechanism of Fructus Aurantii for Major Depression Disorder. Evid. Based Complement. Alternat. Med. 2021, 2021, 6486287. [Google Scholar] [CrossRef] [PubMed]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J Comput Aided Mol Des 2010, 24, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.; Duan, M.; Zhang, C.; Wang, K.; Feng, L.; Song, L.; Wu, S.; Chen, X. Identification of Potential Biomarkers Associated with Acute Myocardial Infarction by Weighted Gene Coexpression Network Analysis. Oxid. Med. Cell. Longev. 2021, 2021, 5553811. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yu, S.Z.; Liang, X.; Li, Y.; Lai, K.P. Vitamin C exerts anti-cadmium induced fracture functions/targets: Bioinformatic and biostructural findings. Food Sci. Hum. Wellness 2022, 11, 1384–1391. [Google Scholar] [CrossRef]
- Gabrilovich, D.; Ishida, T.; Oyama, T.; Ran, S.; Kravtsov, V.; Nadaf, S.; Carbone, D.P. Vascular endothelial growth factor inhibits the development of dendritic cells and dramatically affects the differentiation of multiple hematopoietic lineages in vivo. Blood 1998, 92, 4150–4166. [Google Scholar] [CrossRef]
- Alfaro, C.; Suarez, N.; Gonzalez, A.; Solano, S.; Erro, L.; Dubrot, J.; Palazon, A.; Hervas-Stubbs, S.; Gurpide, A.; Lopez-Picazo, J.M.; et al. Influence of bevacizumab, sunitinib and sorafenib as single agents or in combination on the inhibitory effects of VEGF on human dendritic cell differentiation from monocytes. Br. J. Cancer 2009, 100, 1111–1119. [Google Scholar] [CrossRef]
- Huang, L.; Jia, J.; Liu, R. Decreased serum levels of the angiogenic factors VEGF and TGF-beta1 in Alzheimer’s disease and amnestic mild cognitive impairment. Neurosci. Lett. 2013, 550, 60–63. [Google Scholar] [CrossRef]
- Hohman, T.J.; Bell, S.P.; Jefferson, A.L.; Alzheimer’s Disease Neuroimaging, I. The role of vascular endothelial growth factor in neurodegeneration and cognitive decline: Exploring interactions with biomarkers of Alzheimer disease. JAMA Neurol. 2015, 72, 520–529. [Google Scholar] [CrossRef]
- Minnelli, C.; Cianfruglia, L.; Laudadio, E.; Mobbili, G.; Galeazzi, R.; Armeni, T. Effect of Epigallocatechin-3-Gallate on EGFR Signaling and Migration in Non-Small Cell Lung Cancer. Int. J. Mol. Sci. 2021, 22, 11833. [Google Scholar] [CrossRef]
- Heber, D.; Zhang, Y.; Yang, J.; Ma, J.E.; Henning, S.M.; Li, Z. Green tea, black tea, and oolong tea polyphenols reduce visceral fat and inflammation in mice fed high-fat, high-sucrose obesogenic diets. J. Nutr. 2014, 144, 1385–1393. [Google Scholar] [CrossRef] [PubMed]
- Ando, K.; Kanazawa, S.; Tetsuka, T.; Ohta, S.; Jiang, X.; Tada, T.; Kobayashi, M.; Matsui, N.; Okamoto, T. Induction of Notch signaling by tumor necrosis factor in rheumatoid synovial fibroblasts. Oncogene 2003, 22, 7796–7803. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Chen, J.; Lo, C.Z.; Guo, Q.; Consortium, Z.I.B.; Feng, J.; Zhao, X.M. Impaired type I interferon signaling activity implicated in the peripheral blood transcriptome of preclinical Alzheimer’s disease. EBioMedicine 2022, 82, 104175. [Google Scholar] [CrossRef] [PubMed]
- Ghosh-Roy, A.; Wu, Z.; Goncharov, A.; Jin, Y.; Chisholm, A.D. Calcium and cyclic AMP promote axonal regeneration in Caenorhabditis elegans and require DLK-1 kinase. J. Neurosci. 2010, 30, 3175–3183. [Google Scholar] [CrossRef]
- Chou, C.W.; Huang, W.J.; Tien, L.T.; Wang, S.J. (-)-Epigallocatechin gallate, the most active polyphenolic catechin in green tea, presynaptically facilitates Ca2+-dependent glutamate release via activation of protein kinase C in rat cerebral cortex. Synapse 2007, 61, 889–902. [Google Scholar] [CrossRef]
- Schmid, T.; Zhou, J.; Brune, B. HIF-1 and p53: Communication of transcription factors under hypoxia. J. Cell. Mol. Med. 2004, 8, 423–431. [Google Scholar] [CrossRef]
- Xu, J.; Tu, Y.; Wang, Y.; Xu, X.; Sun, X.; Xie, L.; Zhao, Q.; Guo, Y.; Gu, Y.; Du, J.; et al. Prodrug of epigallocatechin-3-gallate alleviates choroidal neovascularization via down-regulating HIF-1alpha/VEGF/VEGFR2 pathway and M1 type macrophage/microglia polarization. Biomed. Pharmacother. 2020, 121, 109606. [Google Scholar] [CrossRef]
- Li, Y.; Ying, C.; Zuo, X.; Yi, H.; Yi, W.; Meng, Y.; Ikeda, K.; Ye, X.; Yamori, Y.; Sun, X. Green tea polyphenols down-regulate caveolin-1 expression via ERK1/2 and p38MAPK in endothelial cells. J. Nutr. Biochem. 2009, 20, 1021–1027. [Google Scholar] [CrossRef]
- Chen, Y.J.; Chan, D.C.; Chiang, C.K.; Wang, C.C.; Yang, T.H.; Lan, K.C.; Chao, S.C.; Tsai, K.S.; Yang, R.S.; Liu, S.H. Advanced glycation end-products induced VEGF production and inflammatory responses in human synoviocytes via RAGE-NF-kappaB pathway activation. J. Orthop. Res. 2016, 34, 791–800. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Mutation | Mutation Energy (kcal/mol) | Effect |
|---|---|---|---|
| 1 | S50A | −0.23 | NEUTRAL |
| 2 | C61A | −0.01 | NEUTRAL |
| 3 | E67A | 0.02 | NEUTRAL |
| 4 | D63A | 0.03 | NEUTRAL |
| 5 | L66A | 0.05 | NEUTRAL |
| 6 | I35A | 0.23 | NEUTRAL |
| 7 | F47A | 0.36 | NEUTRAL |
| 8 | D34A | 0.41 | NEUTRAL |
| 9 | E64A | 0.61 | DESTABILIZING |
| 10 | F36A | 1.2 | DESTABILIZING |
| Index | Mutation | Mutation Energy (kcal/mol) | Effect |
|---|---|---|---|
| 1 | E67A | 0 | NEUTRAL |
| 2 | G59A | 0.01 | NEUTRAL |
| 3 | C68A | 0.01 | NEUTRAL |
| 4 | D63A | 0.03 | NEUTRAL |
| 5 | L66A | 0.12 | NEUTRAL |
| 6 | S50A | 0.36 | NEUTRAL |
| 7 | C61A | 0.55 | DESTABILIZING |
| 8 | F36A | 0.64 | DESTABILIZING |
| 9 | K48A | 0.78 | DESTABILIZING |
| 10 | E64A | 0.79 | DESTABILIZING |
| Index | Mutation | Mutation Energy (kcal/mol) | Effect |
|---|---|---|---|
| 1 | S50A | −0.58 | STABILIZING |
| 2 | C61A | −0.19 | NEUTRAL |
| 3 | D63A | −0.06 | NEUTRAL |
| 4 | N62A | −0.04 | NEUTRAL |
| 5 | I46A | 0 | NEUTRAL |
| 6 | E67A | 0.18 | NEUTRAL |
| 7 | F47A | 0.28 | NEUTRAL |
| 8 | D34A | 0.37 | NEUTRAL |
| 9 | E64A | 0.53 | DESTABILIZING |
| 10 | F36A | 0.98 | DESTABILIZING |
| Index | Mutation | Mutation Energy (kcal/mol) | Effect |
|---|---|---|---|
| 1 | C61A | −0.32 | NEUTRAL |
| 2 | L66A | −0.12 | NEUTRAL |
| 3 | G59A | −0.08 | NEUTRAL |
| 4 | F47A | −0.04 | NEUTRAL |
| 5 | D63A | 0.02 | NEUTRAL |
| 6 | I46A | 0.36 | NEUTRAL |
| 7 | S50A | 0.45 | NEUTRAL |
| 8 | D34A | 0.46 | NEUTRAL |
| 9 | E64A | 0.61 | DESTABILIZING |
| 10 | K48A | 0.84 | DESTABILIZING |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Yang, X.; Gao, Y.; Han, W. Computer-Aided Screening and Revealing Action Mechanism of Green Tea Polyphenols Intervention in Alzheimer’s Disease. Foods 2023, 12, 635. https://doi.org/10.3390/foods12030635
Wang M, Yang X, Gao Y, Han W. Computer-Aided Screening and Revealing Action Mechanism of Green Tea Polyphenols Intervention in Alzheimer’s Disease. Foods. 2023; 12(3):635. https://doi.org/10.3390/foods12030635
Chicago/Turabian StyleWang, Min, Xiaotang Yang, Yilin Gao, and Weiwei Han. 2023. "Computer-Aided Screening and Revealing Action Mechanism of Green Tea Polyphenols Intervention in Alzheimer’s Disease" Foods 12, no. 3: 635. https://doi.org/10.3390/foods12030635
APA StyleWang, M., Yang, X., Gao, Y., & Han, W. (2023). Computer-Aided Screening and Revealing Action Mechanism of Green Tea Polyphenols Intervention in Alzheimer’s Disease. Foods, 12(3), 635. https://doi.org/10.3390/foods12030635