
Partial Removal of Phenolics Coupled with Alkaline pH Shift Improves Canola Protein Interfacial Properties and Emulsion in In Vitro Digestibility



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Removal of Phenolic Compounds
2.3. Preparation of Protein Isolate and pH12 Shift Treatment
2.4. Determination of Total Phenolic Content (TPC)
2.5. UPLC-PAD-QTOF-MS Analysis of Extracted Phenolic Compounds
2.6. Protein Solubility
2.7. Electrophoresis
2.8. Dynamic Light Scattering (DLS)
2.9. Structure Characterization
2.9.1. Surface Hydrophobicity
2.9.2. Far-UV Circular Dichroism (CD) Spectroscopy
2.10. Interfacial Properties
2.11. Emulsification and Emulsion Properties
2.12. In Vitro Emulsion Digestion and Characterization
2.13. Statistical Analysis
3. Results and Discussion
3.1. Phenolics Removal and Color Improvement
3.2. Protein Solubility and Polypeptide Profile
3.3. Particle Size and Distribution
3.4. Structural Characterization
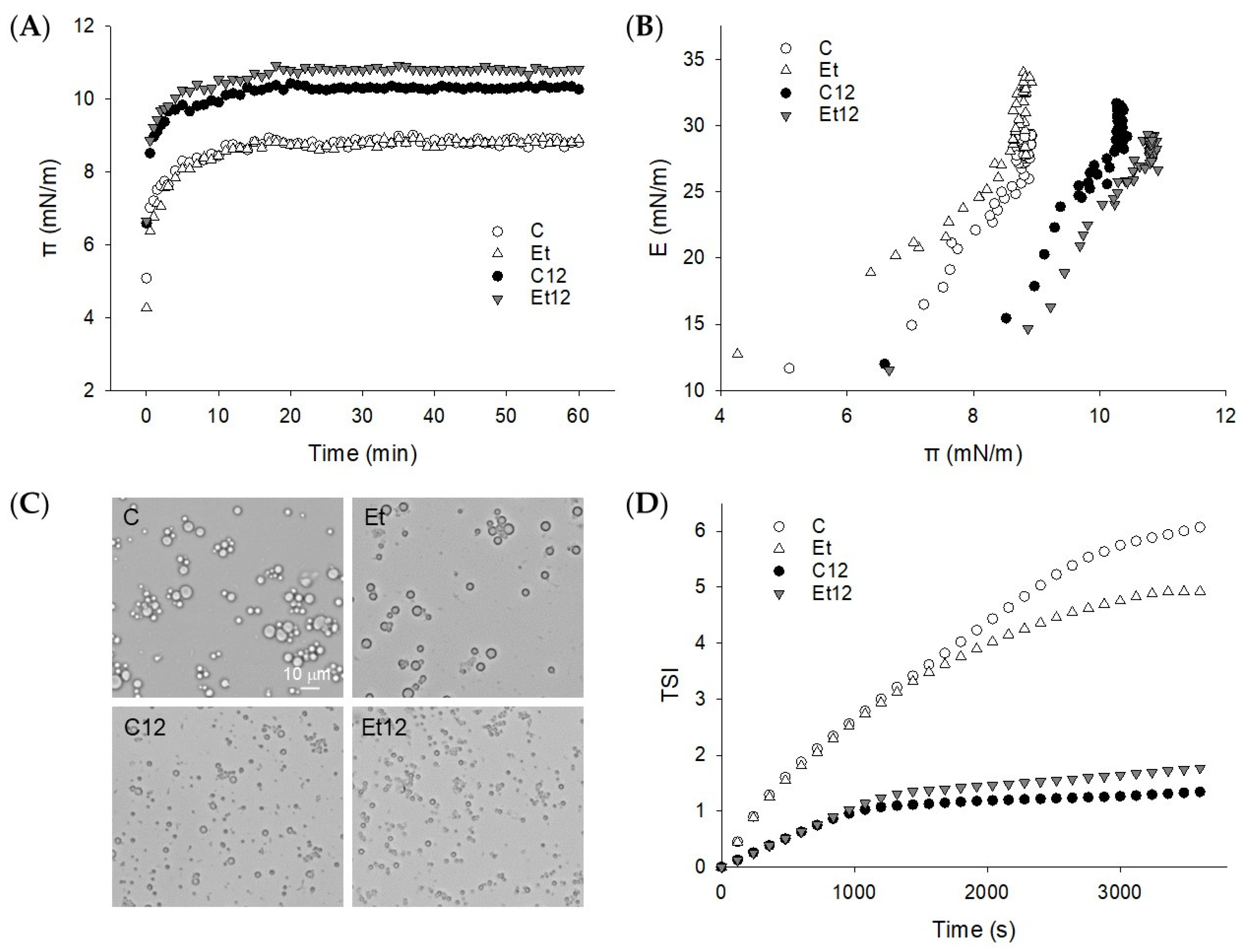
3.5. Interface Properties
3.6. Emulsion Characterization
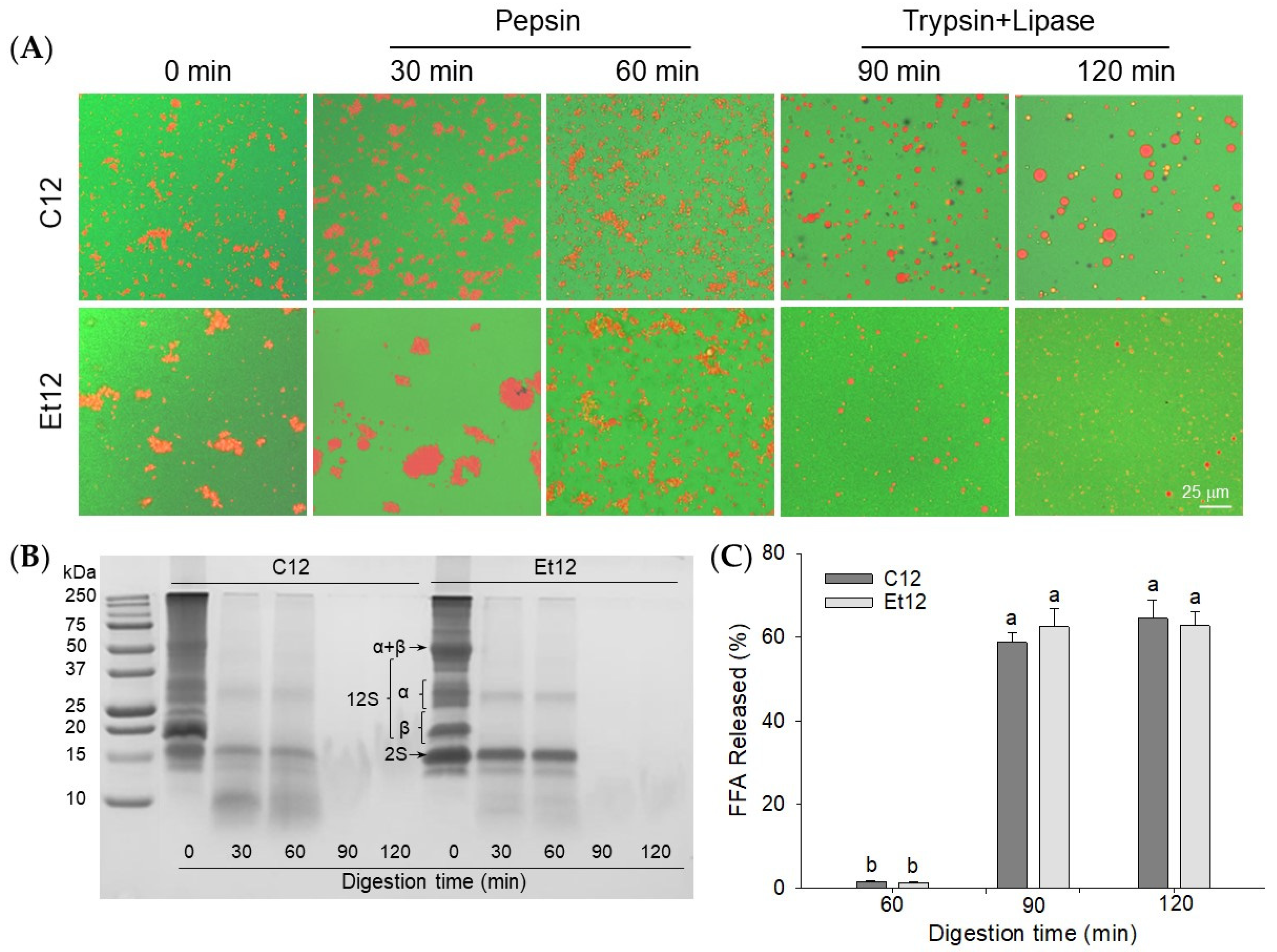
3.7. Emulsion Digestion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, M.; Jiang, T.S.; Pan, J.L. Antioxidant Activities of Rapeseed Protein Hydrolysates. Food Bioprocess Technol. 2009, 4, 1144–1152. [Google Scholar] [CrossRef]
- Akbari, A.; Wu, J. An integrated method of isolating napin and cruciferin from defatted canola meal. LWT 2015, 64, 308–315. [Google Scholar] [CrossRef]
- Sarwar, G.; Blair, R.; Friedman, M.; Gumbmann, M.R.; Hackler, L.R.; Pellett, P.L.; Smith, T.K. Inter-and intra-laboratory var-iability in rat growth assays for estimating protein quality of foods. J. Assoc. Off. Anal. Chem. 1984, 67, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Perera, S.P.; McIntosh, T.C.; Wanasundara, J.P.D. Structural Properties of Cruciferin and Napin of Brassica napus (Canola) Show Distinct Responses to Changes in pH and Temperature. Plants 2016, 5, 36. [Google Scholar] [CrossRef]
- Campbell, L.; Rempel, C.B.; Wanasundara, J.P.D. Canola/rapeseed protein: Future opportunities and directions-workshop pro-ceedings of IRC 2015. Plants 2016, 5, 17. [Google Scholar] [CrossRef]
- Wanasundara, J.P.; McIntosh, T.C.; Perera, S.P.; Withana-Gamage, T.S.; Mitra, P. Canola/rapeseed protein-functionality and nutrition. OCL 2016, 23, D407. [Google Scholar] [CrossRef]
- Tan, S.H.; Mailer, R.J.; Blanchard, C.L.; Agboola, S.O. Canola Proteins for Human Consumption: Extraction, Profile, and Functional Properties. J. Food Sci. 2010, 76, R16–R28. [Google Scholar] [CrossRef]
- Rękas, A.; Ścibisz, I.; Siger, A.; Wroniak, M. The effect of microwave pretreatment of seeds on the stability and degradation kinetics of phenolic compounds in rapeseed oil during long-term storage. Food Chem. 2017, 222, 43–52. [Google Scholar] [CrossRef]
- Kato, A.; Nakai, S. Hydrophobicity determined by a fluorescence probe method and its correlation with surface properties of proteins. Biochim. Biophys. Acta 1980, 624, 13–20. [Google Scholar] [CrossRef]
- Ozdal, T.; Capanoglu, E.; Altay, F. A review on protein–phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- Gruener, L.; Ismond, M. Effects of acetylation and succinylation on the functional properties of the canola 12S globulin. Food Chem. 1997, 60, 513–520. [Google Scholar] [CrossRef]
- Das Purkayastha, M.; Borah, A.K.; Saha, S.; Manhar, A.K.; Mandal, M.; Mahanta, C.L. Effect of maleylation on physicochemical and functional properties of rapeseed protein isolate. J. Food Sci. Technol. 2016, 53, 1784–1797. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, Z.; Razavi, S.M.A.; Varidi, M. Sequential ultrasound and transglutaminase treatments improve functional, rheological, and textural properties of whey protein concentrate. Innov. Food Sci. Emerg. Technol. 2017, 43, 207–215. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, Q.; Xiong, Y.L. A pH shift approach to the improvement of interfacial properties of plant seed proteins. Curr. Opin. Food Sci. 2018, 19, 50–56. [Google Scholar] [CrossRef]
- Liu, Q.; Geng, R.; Zhao, J.; Chen, Q.; Kong, B. Structural and gel textural properties of soy protein isolate when subjected to extreme acid pH-shifting and mild heating processes. J. Agric. Food Chem. 2015, 63, 4853–4861. [Google Scholar] [CrossRef]
- Li, N.; Qi, G.; Sun, X.S.; Stamm, M.J.; Wang, D. Physicochemical Properties and Adhesion Performance of Canola Protein Modified with Sodium Bisulfite. J. Am. Oil Chem. Soc. 2012, 89, 897–908. [Google Scholar] [CrossRef]
- Vuorela, S.; Meyer, A.S.; Heinonen, M. Impact of Isolation Method on the Antioxidant Activity of Rapeseed Meal Phenolics. J. Agric. Food Chem. 2004, 52, 8202–8207. [Google Scholar] [CrossRef]
- Pirestani, S.; Nasirpour, A.; Keramat, J.; Desobry, S. Preparation of chemically modified canola protein isolate with gum Arabic by means of Maillard reaction under wet-heating conditions. Carbohydr. Polym. 2017, 155, 201–207. [Google Scholar] [CrossRef]
- Labuckas, D.O.; Maestri, D.M.; Perelló, M.; Martínez, M.L.; Lamarque, A.L. Phenolics from walnut (Juglans regia L.) kernels: Antioxidant activity and interactions with proteins. Food Chem. 2008, 107, 607–612. [Google Scholar] [CrossRef]
- Thiyam, U.; Stöckmann, H.; Felde, T.Z.; Schwarz, K. Antioxidative effect of the main sinapic acid derivatives from rapeseed and mustard oil by-products. Eur. J. Lipid Sci. Technol. 2006, 108, 239–248. [Google Scholar] [CrossRef]
- Thiyam, U.; Pickardt, C.; Ungewiss, J.; Alfred, B. De-oiled rapeseed and a protein isolate: Characterization of sinapic acid deriva-tives by HPLC-DAD and LC-MS. Eur. Food Res. Technol. 2009, 229, 825–831. [Google Scholar] [CrossRef]
- Jiang, J.; Chen, J.; Xiong, Y.L. Structure and emulsifying properties of soy protein isolate subjected to acid and alkaline pH-shifting process. J. Agric. Food Chem. 2009, 57, 7576–7583. [Google Scholar] [CrossRef]
- Von Staszewski, M.; Ruiz-Henestrosa, V.M.P.; Pilosof, A.M.R. Green tea polyphenols-β-lactoglobulin nanocomplexes: Interfacial behavior, emulsification and oxidation stability of fish oil. Food Hydrocoll. 2014, 35, 505–511. [Google Scholar] [CrossRef]
- Sha, L.; Koosis, A.O.; Wang, Q.; True, A.D.; Xiong, Y.L. Interfacial dilatational and emulsifying properties of ultrasound-treated pea protein. Food Chem. 2021, 350, 129271. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Jiang, J.; Xiong, Y.L. High pressure homogenization combined with pH shift treatment: A process to produce physically and oxidatively stable hemp milk. Food Res. Int. 2018, 106, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Yuan, F.; Gao, Y.; Panya, A.; McClements, D.; Decker, E.A. Influence of whey protein–beet pectin conjugate on the properties and digestibility of β-carotene emulsion during in vitro digestion. Food Chem. 2014, 156, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, R.A.; Pinheiro, A.C.; Vicente, A.A.; Cunha, R.L. In vitro digestion of oil-in-water emulsions stabilized by whey protein nanofibrils. Food Res. Int. 2017, 99, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Diosady, L. Removal of phenolic compounds in the production of high-quality canola protein isolates. Food Res. Int. 2002, 35, 23–30. [Google Scholar] [CrossRef]
- Sosulski, F. Organoleptic and nutritional effects of phenolic compounds on oilseed protein products: A review. J. Am. Oil Chem. Soc. 1979, 56, 711–715. [Google Scholar] [CrossRef]
- Szydłowska-Czerniak, A.; Trokowski, K.; Karlovits, G.; Szłyk, E. Determination of Antioxidant Capacity, Phenolic Acids, and Fatty Acid Composition of Rapeseed Varieties. J. Agric. Food Chem. 2010, 58, 7502–7509. [Google Scholar] [CrossRef]
- Malik, M.A.; Saini, C.S. Polyphenol removal from sunflower seed and kernel: Effect on functional and rheological properties of protein isolates. Food Hydrocoll. 2017, 63, 705–715. [Google Scholar] [CrossRef]
- Stübler, A.-S.; Lesmes, U.; Juadjur, A.; Heinz, V.; Rauh, C.; Shpigelman, A.; Aganovic, K. Impact of pilot-scale processing (thermal, PEF, HPP) on the stability and bioaccessibility of polyphenols and proteins in mixed protein- and polyphenol-rich juice systems. Innov. Food Sci. Emerg. Technol. 2020, 64, 102426. [Google Scholar] [CrossRef]
- Kozlowska, H.; Naczk, M.; Shahidi, F.; Zadernowski, R. Phenolic Acids and Tannins in Rapeseed and Canola. In Canola and Rapeseed; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1990; pp. 193–210. [Google Scholar]
- Wu, J.; Muir, A. Comparative Structural, Emulsifying, and Biological Properties of 2 Major Canola Proteins, Cruciferin and Napin. J. Food Sci. 2008, 73, C210–C216. [Google Scholar] [CrossRef] [PubMed]
- Subaşı, B.G.; Casanova, F.; Capanoglu, E.; Ajalloueian, F.; Sloth, J.J.; Mohammadifar, M.A. Protein extracts from de-oiled sunflower cake: Structural, physico-chemical and functional properties after removal of phenolics. Food Biosci. 2020, 38, 100749. [Google Scholar] [CrossRef]
- Li, L.; He, M.; Wang, N.; Li, M.; Wu, C.; Li, Y.; Teng, F. Sepctroscopic analysis of the effect of vitamin B12-soy protein isolate on the soy protein isolate structure. J. Mol. Liq. 2021, 325, 115148. [Google Scholar] [CrossRef]
- Cao, Y.Y.; Xiong, Y.L.; Cao, Y.G.; True, A.D. Interfacial properties of whey protein foams as influenced by preheating and phe-nolic binding at neutral pH. Food Hydrocoll. 2018, 82, 379–387. [Google Scholar] [CrossRef]
- Lucassen-Reynders, E.H.; Lucassen, J.; Garrett, P.R.; Giles, D.; Hollway, F. Dynamic Surface Measurements as a Tool to Obtain Equation-of-State Data for Soluble Monolayers. Adv. Chem. 1975, 21, 272–285. [Google Scholar] [CrossRef]
- Mitropoulos, V.; Mütze, A.; Fischer, P. Mechanical properties of protein adsorption layers at the air/water and oil/water in-terface: A comparison in light of the thermodynamical stability of proteins. Adv. Colloid Interface Sci. 2014, 206, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, S.D.; von Staszewski, M.; Pilosof, A.M. Green tea polyphenols-whey proteins nanoparticles: Bulk, interfacial and foaming behavior. Food Hydrocoll. 2015, 50, 108–115. [Google Scholar] [CrossRef]
- Singh, H.; Ye, A. Structural and biochemical factors affecting the digestion of protein-stabilized emulsions. Curr. Opin. Colloid Interface Sci. 2013, 18, 360–370. [Google Scholar] [CrossRef]
- Kasprzak, M.M.; Houdijk, J.G.M.; Liddell, S.; Davis, K.; Olukosi, O.A.; Kightley, S.; White, G.; Wiseman, J. Rapeseed napin and cruciferin are readily digested by poultry. J. Anim. Physiol. Anim. Nutr. 2016, 101, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ye, A.; Lee, S.J.; Singh, H. Influence of gastric digestive reaction on subsequent in vitro intestinal digestion of sodium caseinate-stabilized emulsions. Food Funct. 2012, 3, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lin, Q.; Ye, A.; Han, J.; Singh, H. Flocculation of oil-in-water emulsions stabilised by milk protein ingredients under gastric conditions: Impact on in vitro intestinal lipid digestion. Food Hydrocoll. 2019, 88, 272–282. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, J.; Nie, Y.; Sun, X.; Xiong, Y.L. Partial Removal of Phenolics Coupled with Alkaline pH Shift Improves Canola Protein Interfacial Properties and Emulsion in In Vitro Digestibility. Foods 2021, 10, 1283. https://doi.org/10.3390/foods10061283
Jiang J, Nie Y, Sun X, Xiong YL. Partial Removal of Phenolics Coupled with Alkaline pH Shift Improves Canola Protein Interfacial Properties and Emulsion in In Vitro Digestibility. Foods. 2021; 10(6):1283. https://doi.org/10.3390/foods10061283
Chicago/Turabian StyleJiang, Jiang, Yunqing Nie, Xuemei Sun, and Youling L. Xiong. 2021. "Partial Removal of Phenolics Coupled with Alkaline pH Shift Improves Canola Protein Interfacial Properties and Emulsion in In Vitro Digestibility" Foods 10, no. 6: 1283. https://doi.org/10.3390/foods10061283
APA StyleJiang, J., Nie, Y., Sun, X., & Xiong, Y. L. (2021). Partial Removal of Phenolics Coupled with Alkaline pH Shift Improves Canola Protein Interfacial Properties and Emulsion in In Vitro Digestibility. Foods, 10(6), 1283. https://doi.org/10.3390/foods10061283