Phenological Shifts Since 1830 in 29 Native Plant Species of California and Their Responses to Historical Climate Change



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
- Have the selected plant species changed their day of year (DOY) in the preflowering, flowering, fruiting and growth phenophases due to the effects of climate change in California?
- Has the flowering DOY changed across the different Jepson ecoregions due to the effects of climate change based on the selected species?
2. Materials and Methods
2.1. Study Area
2.2. Target Species and Phenological Data
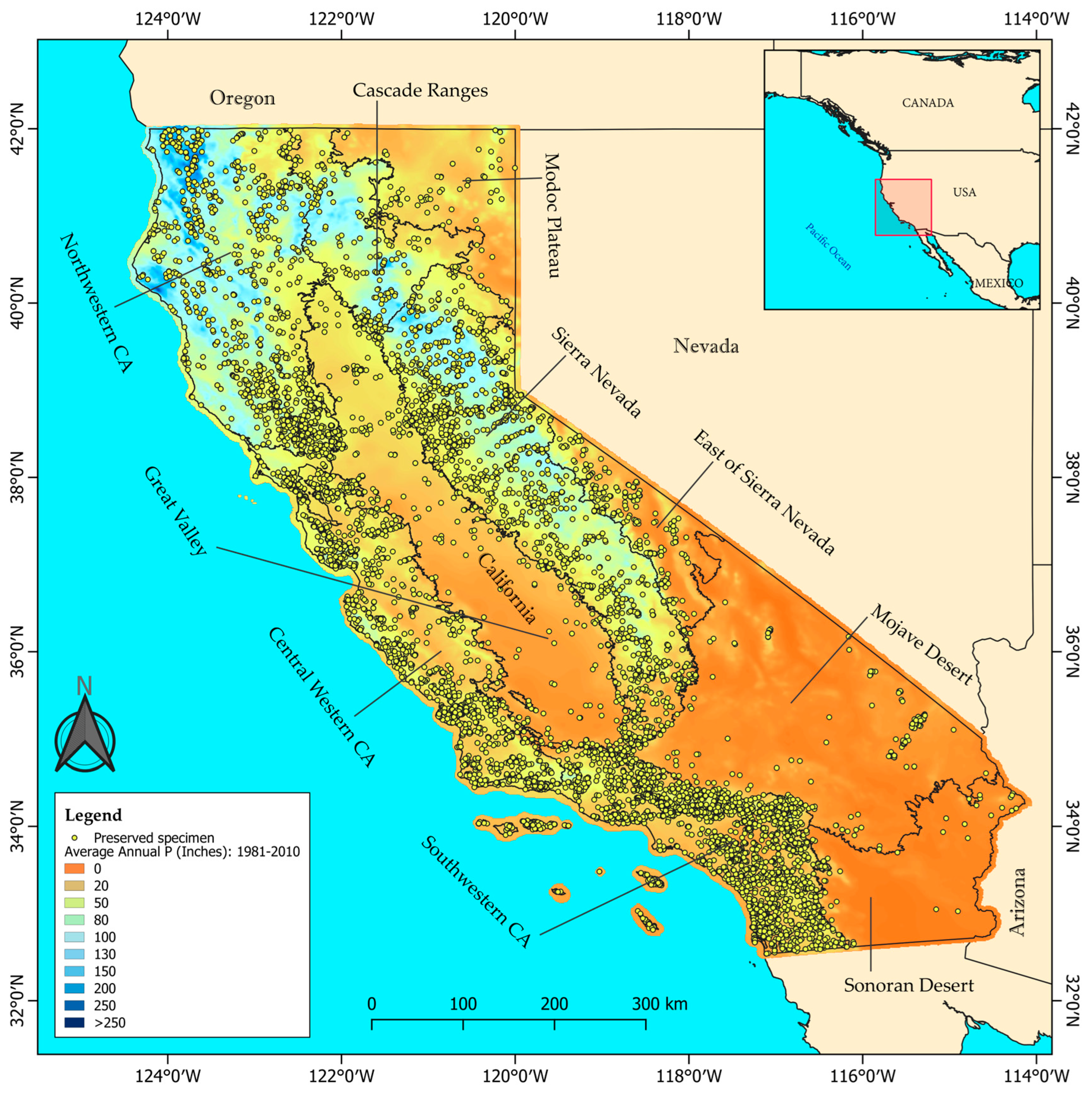
2.3. Climate Data Treatment
2.4. Statistical Analysis
3. Results
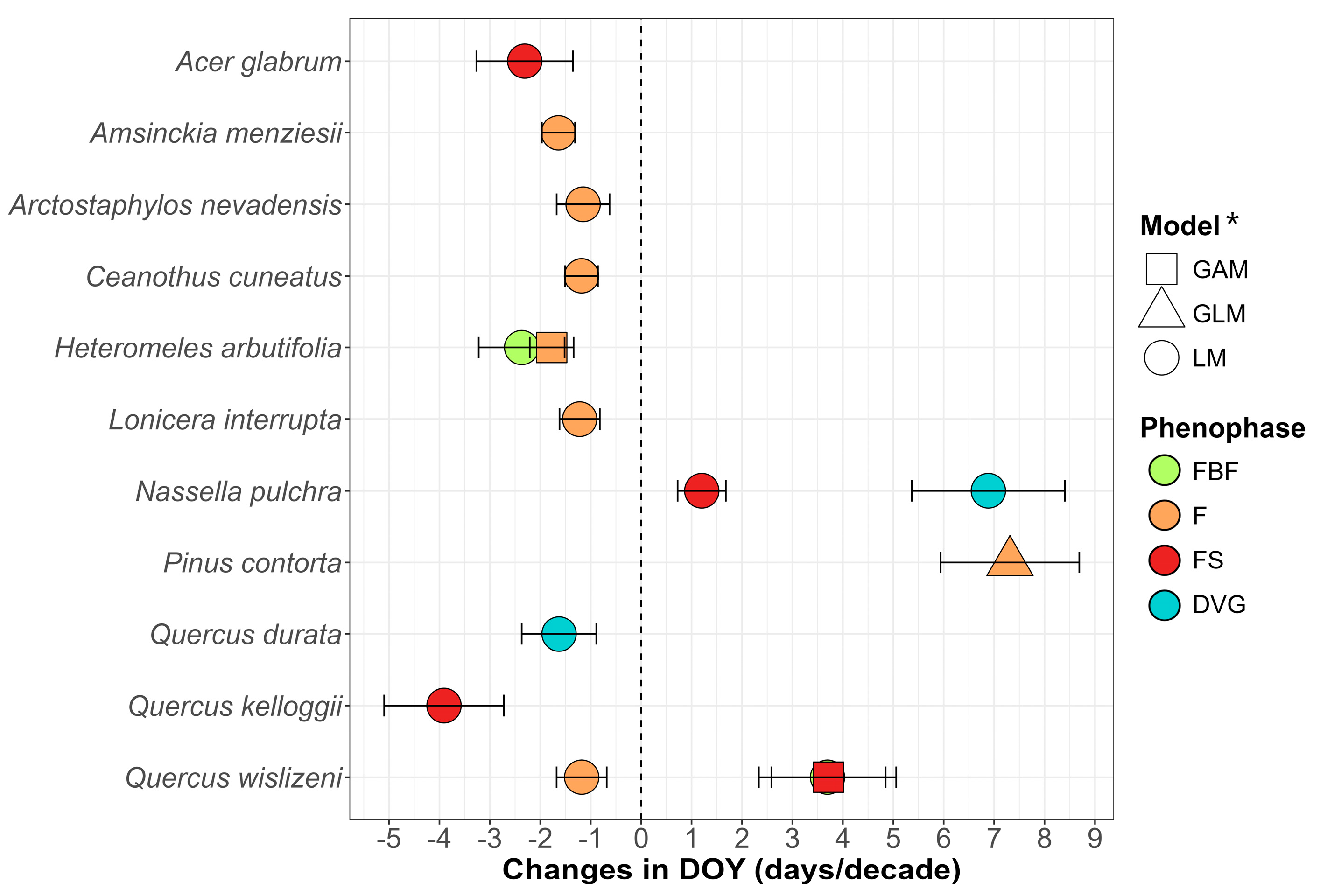
3.1. Phenological Trends by Species
3.1.1. Phenological Trends by Species over Years
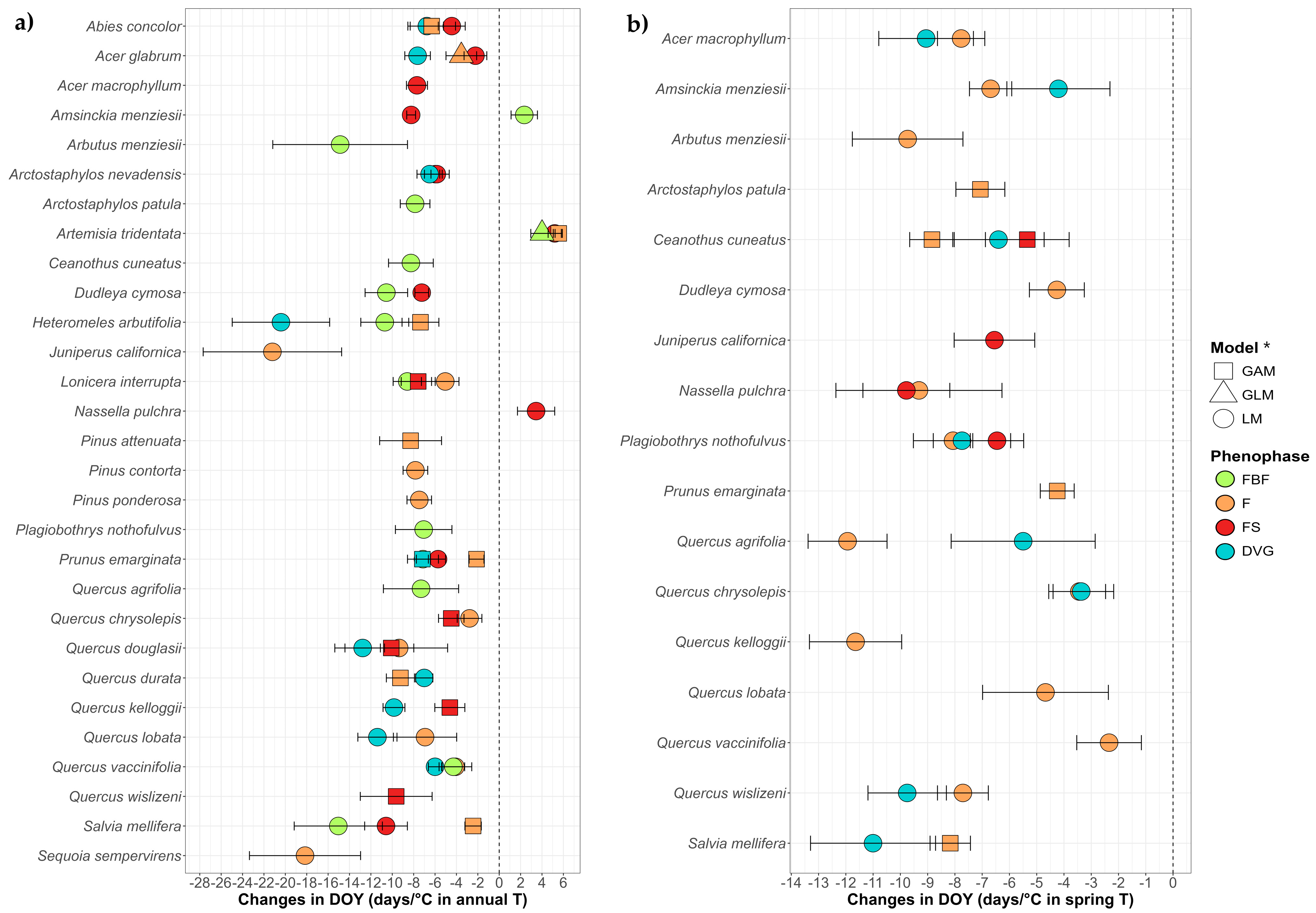
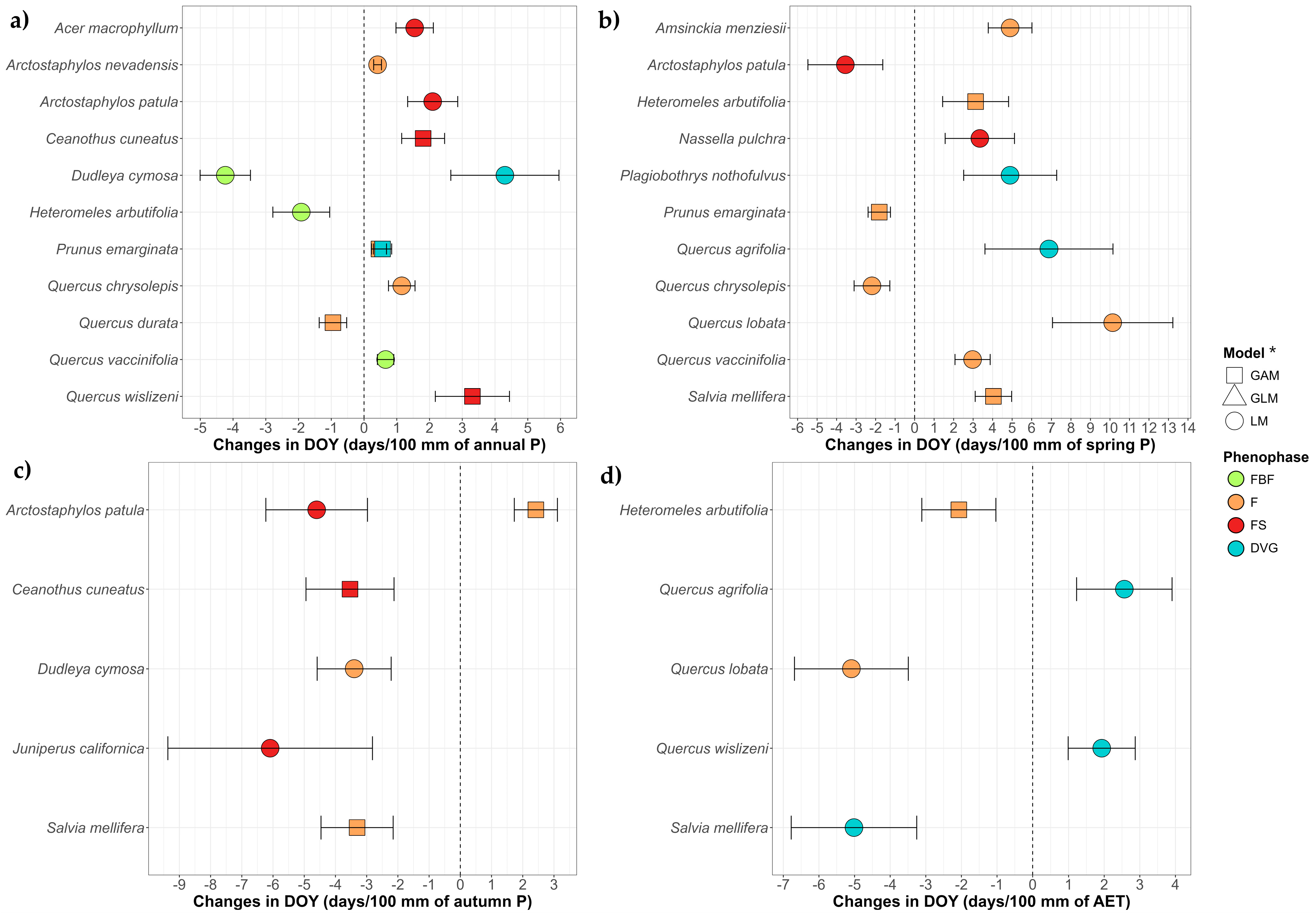
3.1.2. Phenological Trends by Species Through Climate Variables
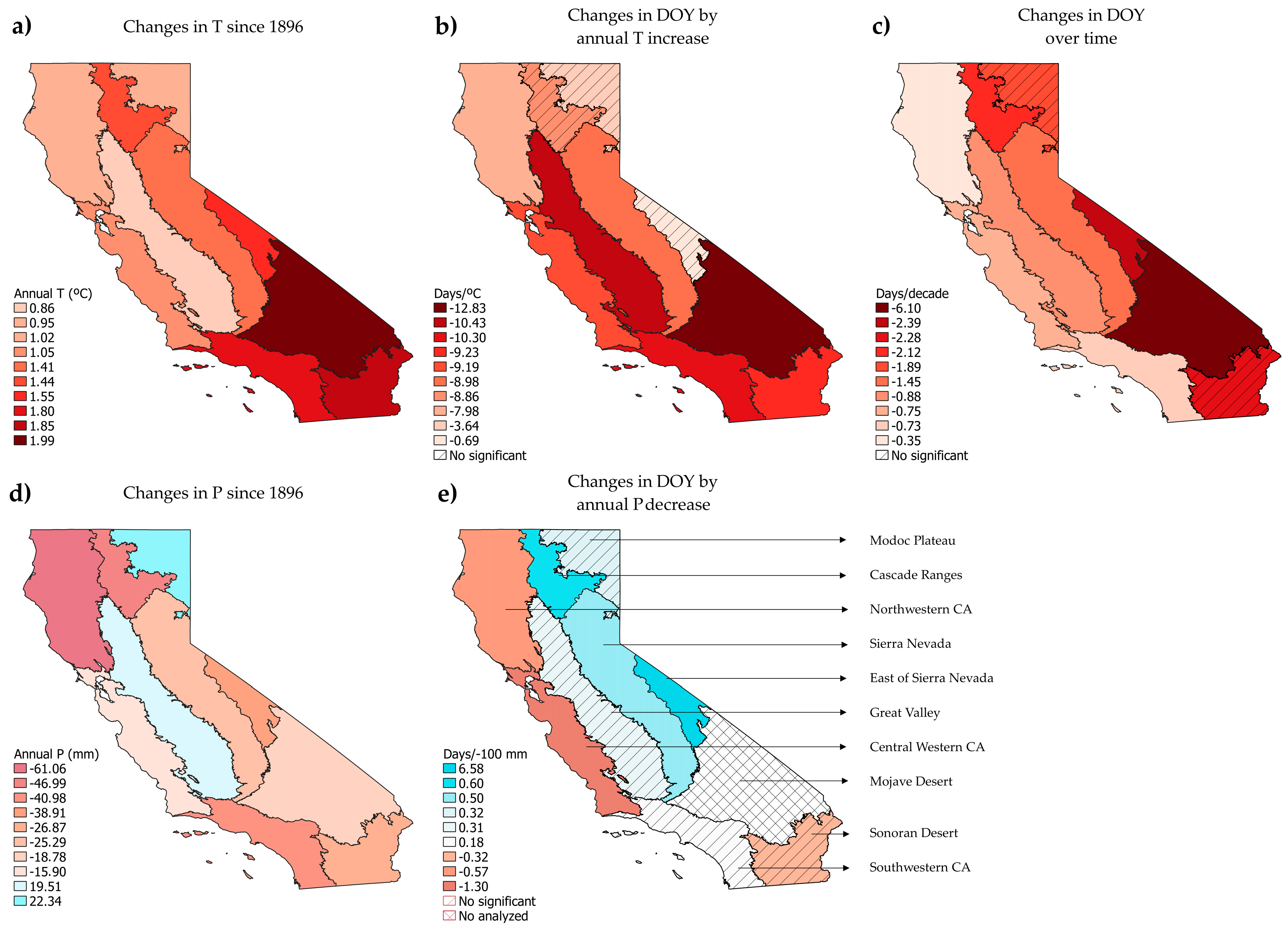
3.2. Phenological Trends by Jepson Ecoregion
4. Discussion
4.1. Phenological Response to Climate Change
4.2. Phenology Shifts in Jepson Ecoregions and Impacts over Vegetation Groups
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baldwin, B.G.; Thornhill, A.H.; Freyman, W.A.; Ackerly, D.D.; Kling, M.M.; Morueta-Holme, N.; Mishler, B.D. Species Richness and Endemism in the Native Flora of California. Am. J. Bot. 2017, 104, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger Climate Classification Updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Franklin, J.; MacDonald, G.M. Climate Change and California Sustainability—Challenges and Solutions. Proc. Natl. Acad. Sci. USA 2024, 121, e2405458121. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.; Franklin, J.; Hernandez, R.R.; Ikegami, M.; Safford, H.D.; Thorne, J.H. Climate Change and California’s Terrestrial Biodiversity. Proc. Natl. Acad. Sci. USA 2024, 121, e2310074121. [Google Scholar] [CrossRef]
- Hickman, J.C. (Ed.) The Jepson Manual: Higher Plants of California; University of California Press: Berkeley, CA, USA, 1993; ISBN 978-0-520-08255-7. [Google Scholar]
- Burge, D.O.; Thorne, J.H.; Harrison, S.P.; O’Brien, B.C.; Rebman, J.P.; Shevock, J.R.; Alverson, E.R.; Hardison, L.K.; Rodríguez, J.D.; Junak, S.A.; et al. Plant Diversity and Endemism in the California Floristic Province. Madroño 2016, 63, 3–206. [Google Scholar] [CrossRef]
- USNVC (United States National Vegetation Classification) Database [Version 2.04]; Fed. Geogr. Data Comm. Veg. Subcomm. 2022. Available online: https://usnvc.org/explore-classification/ (accessed on 30 January 2024).
- Faber-Langendoen, D.; Keeler-Wolf, T.; Meidinger, D.; Tart, D.; Hoagland, B.; Josse, C.; Navarro, G.; Ponomarenko, S.; Saucier, J.-P.; Weakley, A.; et al. EcoVeg: A New Approach to Vegetation Description and Classification. Ecol. Monogr. 2014, 84, 533–561. [Google Scholar] [CrossRef]
- Pignatti, S.; Oberdorfer, E.; Schaminée, J.H.J.; Westhoff, V. On the Concept of Vegetation Class in Phytosociology. J. Veg. Sci. 1995, 6, 143–152. [Google Scholar] [CrossRef]
- Thorne, J.H.; Boyton, R.M.; Holguin, A.J.; Stewart, J.A.E.; Bjorkman, J. A Climate Change Vulnerability Assessment of California’s Terrestrial Vegetation; California Department of Wildlife and Fisheries: Sacramento, CA, USA, 2016. [Google Scholar]
- SWAP. California State Wildlife Action Plan 2015 Update; California Department of Fish and Wildlife (CDFW): Sacramento, CA, USA, 2015. [Google Scholar]
- IPCC. Climate Change 2021–The Physical Science Basis: Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, 1st ed.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; ISBN 978-1-00-915789-6. [Google Scholar]
- Lionello, P.; Scarascia, L. The Relation between Climate Change in the Mediterranean Region and Global Warming. Reg. Environ. Change 2018, 18, 1481–1493. [Google Scholar] [CrossRef]
- Luna, M.Y.; Guijarro, J.A.; López, J.A. A Monthly Precipitation Database for Spain (1851–2008): Reconstruction, Homogeneity and Trends. Adv. Sci. Res. 2012, 8, 1–4. [Google Scholar] [CrossRef]
- IPCC. Global Warming of 1.5 °C: IPCC Special Report on Impacts of Global Warming of 1.5 °C Above Pre-Industrial Levels in Context of Strengthening Response to Climate Change, Sustainable Development, and Efforts to Eradicate Poverty, 1st ed.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2018; ISBN 978-1-00-915794-0. [Google Scholar]
- Barnett, T.P.; Pierce, D.W.; Hidalgo, H.G.; Bonfils, C.; Santer, B.D.; Das, T.; Bala, G.; Wood, A.W.; Nozawa, T.; Mirin, A.A.; et al. Human-Induced Changes in the Hydrology of the Western United States. Science 2008, 319, 1080–1083. [Google Scholar] [CrossRef]
- OEHHA. Indicators of Climate Change in California, 4th ed.; California Environmental Protection Agency, Office of Environmental Health Hazard Assessment, OEHHA: Sacramento, CA, USA, 2022.
- Swain, D.L.; Langenbrunner, B.; Neelin, J.D.; Hall, A. Increasing Precipitation Volatility in Twenty-First-Century California. Nat. Clim. Change 2018, 8, 427–433. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and Future Köppen-Geiger Climate Classification Maps at 1-Km Resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [PubMed]
- Beck, H.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Lutsko, N.J.; Dufour, A.; Zeng, Z.; Jiang, X.; Van Dijk, A.I.J.M.; Miralles, D.G. High-Resolution (1 Km) Köppen-Geiger Maps for 1901–2099 Based on Constrained CMIP6 Projections. Sci. Data 2023, 10, 724. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C.; Yohe, G. A Globally Coherent Fingerprint of Climate Change Impacts across Natural Systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Menzel; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A.; et al. European Phenological Response to Climate Change Matches the Warming Pattern. Glob. Change Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Keeling, C.D.; Chin, J.F.S.; Whorf, T.P. Increased Activity of Northern Vegetation Inferred from Atmospheric CO2 Measurements. Nature 1996, 382, 146–149. [Google Scholar] [CrossRef]
- Love, N.L.R.; Park, I.W.; Mazer, S.J. A New Phenological Metric for Use in Pheno-climatic Models: A Case Study Using Herbarium Specimens of Streptanthus tortuosus. Appl. Plant Sci. 2019, 7, e11276. [Google Scholar] [CrossRef]
- Tang, J.; Körner, C.; Muraoka, H.; Piao, S.; Shen, M.; Thackeray, S.J.; Yang, X. Emerging Opportunities and Challenges in Phenology: A Review. Ecosphere 2016, 7, e01436. [Google Scholar] [CrossRef]
- Bertin, R.I. Plant Phenology And Distribution In Relation To Recent Climate Change. J. Torrey Bot. Soc. 2008, 135, 126–146. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I.; Comas, P. Changed Plant and Animal Life Cycles from 1952 to 2000 in the Mediterranean Region: Phenological Effects of Climate Warming. Glob. Change Biol. 2002, 8, 531–544. [Google Scholar] [CrossRef]
- Rathcke, B.; Lacey, E.P. Phenological Patterns of Terrestrial Plants. Annu. Rev. Ecol. Syst. 1985, 16, 179–214. [Google Scholar] [CrossRef]
- Westman, W.E. Seasonal Dimorphism of Foliage in Californian Coastal Sage Scrub. Oecologia 1981, 51, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Morellato, L.P.C.; Alberton, B.; Alvarado, S.T.; Borges, B.; Buisson, E.; Camargo, M.G.G.; Cancian, L.F.; Carstensen, D.W.; Escobar, D.F.E.; Leite, P.T.P.; et al. Linking Plant Phenology to Conservation Biology. Biol. Conserv. 2016, 195, 60–72. [Google Scholar] [CrossRef]
- Rafferty, N.E.; Bertelsen, C.D.; Bronstein, J.L. Later Flowering Is Associated with a Compressed Flowering Season and Reduced Reproductive Output in an Early Season Floral Resource. Oikos 2016, 125, 821–828. [Google Scholar] [CrossRef]
- Navarro, T.; Nieto, J.M.; Pérez Latorre, A.V.; Cabezudo, B. Estudios Fenomorfológicos En La Vegetación Del Sur de España. III: Comportamiento Estacional de Una Comunidad de Badlands (Tabernas, Almería, España). Acta Bot. Malacit. 1993, 18, 189–198. [Google Scholar] [CrossRef]
- Orshan, G. Plant Pheno-Morphological Studies in Mediterranean Type Ecosystems; Springer: Dordrecht, The Netherlands, 1988; ISBN 978-94-009-3107-7. [Google Scholar]
- Lieth, H. Purposes of a Phenology Book. In Phenology and Seasonality Modeling; Lieth, H., Ed.; Springer: Berlin/Heidelberg, Germany, 1974; Volume 8, pp. 3–19. ISBN 978-3-642-51865-2. [Google Scholar]
- Picornell, A.; Buters, J.; Rojo, J.; Traidl-Hoffmann, C.; Damialis, A.; Menzel, A.; Bergmann, K.C.; Werchan, M.; Schmidt-Weber, C.; Oteros, J. Predicting the Start, Peak and End of the Betula Pollen Season in Bavaria, Germany. Sci. Total Environ. 2019, 690, 1299–1309. [Google Scholar] [CrossRef]
- Schwartz, M.D. Detecting the Onset of Spring: A Possible Application of Phenological Models. Clim. Res. 1990, 1, 23–29. [Google Scholar] [CrossRef]
- Mazer, S.J.; Gerst, K.L.; Matthews, E.R.; Evenden, A. Species-Specific Phenological Responses to Winter Temperature and Precipitation in a Water-Limited Ecosystem. Ecosphere 2015, 6, art98. [Google Scholar] [CrossRef]
- Wang, J.; Liu, D.; Ciais, P.; Peñuelas, J. Decreasing Rainfall Frequency Contributes to Earlier Leaf Onset in Northern Ecosystems. Nat. Clim. Change 2022, 12, 386–392. [Google Scholar] [CrossRef]
- Pearson, K.D.; Love, N.L.R.; Ramirez-Parada, T.; Mazer, S.J.; Yost, J.M. Phenological Trends in the California Poppy (Eschscholzia californica): Digitized Herbarium Specimens Reveal Intraspecific Variation in the Sensitivity of Flowering Date to Climate Change. Madroño 2021, 68, 343–359. [Google Scholar] [CrossRef]
- Primack, D.; Imbres, C.; Primack, R.B.; Miller-Rushing, A.J.; Del Tredici, P. Herbarium Specimens Demonstrate Earlier Flowering Times in Response to Warming in Boston. Am. J. Bot. 2004, 91, 1260–1264. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Parada, T.H.; Park, I.W.; Record, S.; Davis, C.C.; Ellison, A.M.; Mazer, S.J. Plasticity and Not Adaptation Is the Primary Source of Temperature-Mediated Variation in Flowering Phenology in North America. Nat. Ecol. Evol. 2024, 8, 467–476. [Google Scholar] [CrossRef]
- Zettlemoyer, M.A.; Peterson, M.L. Does Phenological Plasticity Help or Hinder Range Shifts Under Climate Change? Front. Ecol. Evol. 2021, 9, 689192. [Google Scholar] [CrossRef]
- Del Río, S.; Álvarez-Esteban, R.; Cano, E.; Pinto-Gomes, C.; Penas, Á. Potential Impacts of Climate Change on Habitat Suitability of Fagus sylvatica L. Forests in Spain. Plant Biosyst.-Int. J. Deal. Asp. Plant Biol. 2018, 152, 1205–1213. [Google Scholar] [CrossRef]
- Hidalgo-Triana, N.; Casimiro-Soriguer Solanas, F.; Solakis Tena, A.; Manteca-Bautista, D.; Picornell, A.; García-Sánchez, J.; Navarro, T.; Pérez-Latorre, A.V. Assessment Protocol to Evaluate the Degree of Conservation of Habitats of Community Interest: A Case Study for the 5220* HCI in the Westernmost Localities of Europe. Land 2023, 12, 190. [Google Scholar] [CrossRef]
- Kesseli, J.E. The Climates of California According to the Koppen Classification. Geogr. Rev. 1942, 32, 476–480. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; ter Steege, H.; Morgan, H.D.; van der Heijden, M.G.A.; et al. A Handbook of Protocols for Standardised and Easy Measurement of Plant Functional Traits Worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Miller-Rushing, A.J.; Primack, R.B.; Primack, D.; Mukunda, S. Photographs and Herbarium Specimens as Tools to Document Phenological Changes in Response to Global Warming. Am. J. Bot. 2006, 93, 1667–1674. [Google Scholar] [CrossRef]
- CaraDonna, P.J.; Iler, A.M.; Inouye, D.W. Shifts in Flowering Phenology Reshape a Subalpine Plant Community. Proc. Natl. Acad. Sci. USA 2014, 111, 4916–4921. [Google Scholar] [CrossRef]
- Miller-Rushing, A.J.; Primack, R.B. Global Warming and Flowering Times in Thoreau’s Concord: A Community Perspective. Ecology 2008, 89, 332–341. [Google Scholar] [CrossRef]
- Moussus, J.-P.; Julliard, R.; Jiguet, F. Featuring 10 Phenological Estimators Using Simulated Data: Featuring the Behaviour of Phenological Estimators. Methods Ecol. Evol. 2010, 1, 140–150. [Google Scholar] [CrossRef]
- Calinger, K.M.; Queenborough, S.; Curtis, P.S. Herbarium Specimens Reveal the Footprint of Climate Change on Flowering Trends across North-central North America. Ecol. Lett. 2013, 16, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Diskin, E.; Proctor, H.; Jebb, M.; Sparks, T.; Donnelly, A. The Phenology of Rubus fruticosus in Ireland: Herbarium Specimens Provide Evidence for the Response of Phenophases to Temperature, with Implications for Climate Warming. Int. J. Biometeorol. 2012, 56, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Bertin, R.I. Climate Change and Flowering Phenology in Worcester County, Massachusetts. Int. J. Plant Sci. 2015, 176, 107–119. [Google Scholar] [CrossRef]
- Milla, R.; Castro-Díez, P.; Montserrat-Martí, G. Phenology of Mediterranean Woody Plants from NE Spain: Synchrony, Seasonality, and Relationships among Phenophases. Flora—Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 190–199. [Google Scholar] [CrossRef]
- Navarro, T.; Hidalgo-Triana, N. Variations in Leaf Traits Modulate Plant Vegetative and Reproductive Phenological Sequencing Across Arid Mediterranean Shrublands. Front. Plant Sci. 2021, 12, 708367. [Google Scholar] [CrossRef]
- CCH2 Portal. CCH2: Specimen Data from the Consortium of California Herbaria. 2024. Available online: https://www.cch2.org/portal/ (accessed on 30 September 2024).
- Flint, L.; Flint, A.; Stern, M. The Basin Characterization Model—A Regional Water Balance Software Package; U.S. Geological Survey Techniques and Methods 6–H1; U.S. Geological Survey: Reston, VA, USA, 2021; p. 85. [Google Scholar]
- Daly, C.; Halbleib, M.; Smith, J.I.; Gibson, W.P.; Doggett, M.K.; Taylor, G.H.; Curtis, J.; Pasteris, P.P. Physiographically Sensitive Mapping of Climatological Temperature and Precipitation across the Conterminous United States. Int. J. Climatol. 2008, 28, 2031–2064. [Google Scholar] [CrossRef]
- Flint, L.E.; Flint, A.L. Downscaling Future Climate Scenarios to Fine Scales for Hydrologic and Ecological Modeling and Analysis. Ecol. Process. 2012, 1, 2. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Gordo, O.; Sanz, J.J. Impact of Climate Change on Plant Phenology in Mediterranean Ecosystems. Glob. Change Biol. 2010, 16, 1082–1106. [Google Scholar] [CrossRef]
- Gallagher, R.V.; Hughes, L.; Leishman, M.R. Phenological Trends among Australian Alpine Species: Using Herbarium Records to Identify Climate-Change Indicators. Aust. J. Bot. 2009, 57, 1. [Google Scholar] [CrossRef]
- Menzel, A.; Yuan, Y.; Matiu, M.; Sparks, T.; Scheifinger, H.; Gehrig, R.; Estrella, N. Climate Change Fingerprints in Recent European Plant Phenology. Glob. Change Biol. 2020, 26, 2599–2612. [Google Scholar] [CrossRef] [PubMed]
- Picornell, A.; Smith, M.; Rojo, J. Climate Change Related Phenological Decoupling in Species Belonging to the Betulaceae Family. Int. J. Biometeorol. 2023, 67, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Nelder, J.A.; Wedderburn, R.W.M. Generalized Linear Models. J. R. Stat. Soc. Ser. Gen. 1972, 135, 370. [Google Scholar] [CrossRef]
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman & Hall/CRC texts in statistical science; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2017; ISBN 978-1-4987-2833-1. [Google Scholar]
- Marra, G.; Wood, S.N. Practical Variable Selection for Generalized Additive Models. Comput. Stat. Data Anal. 2011, 55, 2372–2387. [Google Scholar] [CrossRef]
- Aho, K.; Derryberry, D.; Peterson, T. Model Selection for Ecologists: The Worldviews of AIC and BIC. Ecology 2014, 95, 631–636. [Google Scholar] [CrossRef]
- Mote, P.W. Climate-Driven Variability and Trends in Mountain Snowpack in Western North America. J. Clim. 2006, 19, 6209–6220. [Google Scholar] [CrossRef]
- Thorne, J.H.; Boynton, R.M.; Flint, L.E.; Flint, A.L. The Magnitude and Spatial Patterns of Historical and Future Hydrologic Change in California’s Watersheds. Ecosphere 2015, 6, 1–30. [Google Scholar] [CrossRef]
- Bernal, M.; Estiarte, M.; Peñuelas, J. Drought Advances Spring Growth Phenology of the Mediterranean Shrub Erica multiflora. Plant Biol. 2011, 13, 252–257. [Google Scholar] [CrossRef]
- Jentsch, A.; Kreyling, J.; Boettcher-Treschkow, J.; Beierkuhnlein, C. Beyond Gradual Warming: Extreme Weather Events Alter Flower Phenology of European Grassland and Heath Species. Glob. Change Biol. 2009, 15, 837–849. [Google Scholar] [CrossRef]
- Chen, L.; Huang, J.-G.; Ma, Q.; Hänninen, H.; Rossi, S.; Piao, S.; Bergeron, Y. Spring Phenology at Different Altitudes Is Becoming More Uniform under Global Warming in Europe. Glob. Change Biol. 2018, 24, 3969–3975. [Google Scholar] [CrossRef]
- Gordo, O.; Sanz, J.J. Long-term Temporal Changes of Plant Phenology in the Western Mediterranean. Glob. Change Biol. 2009, 15, 1930–1948. [Google Scholar] [CrossRef]
- Vogel, J. Drivers of Phenological Changes in Southern Europe. Int. J. Biometeorol. 2022, 66, 1903–1914. [Google Scholar] [CrossRef] [PubMed]
- Doi, H.; Katano, I. Phenological Timings of Leaf Budburst with Climate Change in Japan. Agric. For. Meteorol. 2008, 148, 512–516. [Google Scholar] [CrossRef]
- Estrella, N.; Sparks, T.H.; Menzel, A. Trends and Temperature Response in the Phenology of Crops in Germany. Glob. Change Biol. 2007, 13, 1737–1747. [Google Scholar] [CrossRef]
- Rondinel-Mendoza, K.V.; Lorite, J.; Marín-Rodulfo, M.; Cañadas, E.M. Tracking Phenological Changes over 183 Years in Endemic Species of a Mediterranean Mountain (Sierra Nevada, SE Spain) Using Herbarium Specimens. Plants 2024, 13, 522. [Google Scholar] [CrossRef]
- Bock, A.; Sparks, T.H.; Estrella, N.; Jee, N.; Casebow, A.; Schunk, C.; Leuchner, M.; Menzel, A. Changes in First Flowering Dates and Flowering Duration of 232 Plant Species on the Island of Guernsey. Glob. Change Biol. 2014, 20, 3508–3519. [Google Scholar] [CrossRef]
- Reed, P.B.; Pfeifer-Meister, L.E.; Roy, B.A.; Johnson, B.R.; Bailes, G.T.; Nelson, A.A.; Boulay, M.C.; Hamman, S.T.; Bridgham, S.D. Prairie Plant Phenology Driven More by Temperature than Moisture in Climate Manipulations across a Latitudinal Gradient in the Pacific Northwest, USA. Ecol. Evol. 2019, 9, 3637–3650. [Google Scholar] [CrossRef]
- Kopp, C.W.; Neto-Bradley, B.M.; Lipsen, L.P.J.; Sandhar, J.; Smith, S. Herbarium Records Indicate Variation in Bloom-Time Sensitivity to Temperature across a Geographically Diverse Region. Int. J. Biometeorol. 2020, 64, 873–880. [Google Scholar] [CrossRef]
- Mooney, H.A.; Parsons, D.J.; Kummerow, J. Plant Development in Mediterranean Climates. In Phenology and Seasonality Modeling; Lieth, H., Ed.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 1974; Volume 8, pp. 255–267. ISBN 978-3-642-51865-2. [Google Scholar]
- Schwartz, M.D.; Karl, T.R. Spring Phenology: Nature’s Experiment to Detect the Effect of “Green-Up” on Surface Maximum Temperatures. Mon. Weather Rev. 1990, 118, 883–890. [Google Scholar] [CrossRef]
- Donohue, K. Germination Timing Influences Natural Selection on Life-history Characters in Arabidopsis thaliana. Ecology 2002, 83, 1006–1016. [Google Scholar] [CrossRef]
- Ettinger, A.K.; Gee, S.; Wolkovich, E.M. Phenological Sequences: How Early-season Events Define Those That Follow. Am. J. Bot. 2018, 105, 1771–1780. [Google Scholar] [CrossRef] [PubMed]
- Kozlowski, J. Optimal Alllocation of Resources to Growth and Reproduction: Implications for Age and Size at Maturity. Trends Ecol. Evol. 1992, 7, 15–19. [Google Scholar] [CrossRef]
- Zhang, X.; Tarpley, D.; Sullivan, J.T. Diverse Responses of Vegetation Phenology to a Warming Climate. Geophys. Res. Lett. 2007, 34, L19405. [Google Scholar] [CrossRef]
- Sherry, R.A.; Zhou, X.; Gu, S.; Arnone, J.A.; Schimel, D.S.; Verburg, P.S.; Wallace, L.L.; Luo, Y. Divergence of Reproductive Phenology under Climate Warming. Proc. Natl. Acad. Sci. USA 2007, 104, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.M.I.; Tansey, C.; Smithers, R.J.; Phillimore, A.B. Predicting a Change in the Order of Spring Phenology in Temperate Forests. Glob. Change Biol. 2015, 21, 2603–2611. [Google Scholar] [CrossRef]
- Wilkinson, M.; Eaton, E.L.; Morison, J.I.L. Variation in the Date of Budburst in Quercus robur and Q. Petraea across a Range of Provenances Grown in Southern England. Eur. J. For. Res. 2017, 136, 1–12. [Google Scholar] [CrossRef]
- Fryer, J.L. Quercus wislizeni. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory: Missoula, MT, USA, 2012. Available online: https://www.fs.usda.gov/database/feis/plants/tree/quewis/all.html (accessed on 10 January 2025).
- Augspurger, C.K.; Salk, C.F. Constraints of Cold and Shade on the Phenology of Spring Ephemeral Herb Species. J. Ecol. 2017, 105, 246–254. [Google Scholar] [CrossRef]
- Risser, P.; Cottam, G. Influence of Temperature on the Dormancy of Some Spring Emphemerals. Ecology 1967, 48, 500–503. [Google Scholar] [CrossRef]
- Alecrim, E.F.; Sargent, R.D.; Forrest, J.R.K. Higher-latitude Spring-flowering Herbs Advance Their Phenology More than Trees with Warming Temperatures. J. Ecol. 2023, 111, 156–169. [Google Scholar] [CrossRef]
- Fazlioglu, F. Shifts in the Timing of the Early Flowering in Plants from a Semi-Arid Ecoregion under Climate Change. Biologia 2019, 74, 437–446. [Google Scholar] [CrossRef]
- Sweet, L.C.; Holt, J.S. Establishment Stage Competition between Exotic Crimson Fountaingrass (Pennisetum Setaceum, C4) and Native Purple Needlegrass (Stipa Pulchra, C3). Invasive Plant Sci. Manag. 2015, 8, 139–150. [Google Scholar] [CrossRef]
- Davis, C.C.; Willis, C.G.; Primack, R.B.; Miller-Rushing, A.J. The Importance of Phylogeny to the Study of Phenological Response to Global Climate Change. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3201–3213. [Google Scholar] [CrossRef] [PubMed]
- Parker, V.T. Absence of Flowering Shifts in Arctostaphylos and Ceanothus over the Past Century of Climate Warming. Madroño 2021, 68, 461–472. [Google Scholar] [CrossRef]
- Cayan, D.R.; Dettinger, M.D.; Kammerdiener, S.A.; Caprio, J.M.; Peterson, D.H. Changes in the Onset of Spring in the Western United States. Bull. Am. Meteorol. Soc. 2001, 82, 399–415. [Google Scholar] [CrossRef]
- Mahaffey, A.A.; Ewers, F.W.; Bozak, K.; Bobich, E.G. Factors Affecting Survival of California Juniper in Its Lower Elevational Range in the Northwestern Sonoran Desert. Plant Ecol. 2020, 221, 501–514. [Google Scholar] [CrossRef]
- Meng, L.; Zhou, Y.; Gu, L.; Richardson, A.D.; Peñuelas, J.; Fu, Y.; Wang, Y.; Asrar, G.R.; De Boeck, H.J.; Mao, J.; et al. Photoperiod Decelerates the Advance of Spring Phenology of Six Deciduous Tree Species under Climate Warming. Glob. Change Biol. 2021, 27, 2914–2927. [Google Scholar] [CrossRef]
- Miller-Rushing, A.J.; Inouye, D.W.; Primack, R.B. How Well Do First Flowering Dates Measure Plant Responses to Climate Change? The Effects of Population Size and Sampling Frequency. J. Ecol. 2008, 96, 1289–1296. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, T.; El-Kassaby, Y.A. Contributions of Dynamic Environmental Signals during Life-Cycle Transitions to Early Life-History Traits in Lodgepole Pine (Pinus contorta Dougl.). Biogeosciences 2015, 13, 2945–2958. [Google Scholar] [CrossRef]
- Betzen, J.J.; Ramsey, A.; Omdal, D.; Ettl, G.J.; Tobin, P.C. Bigleaf Maple, Acer macrophyllum Pursh, Decline in Western Washington, USA. For. Ecol. Manag. 2021, 501, 119681. [Google Scholar] [CrossRef]
- Armstrong-Herniman, W.; Greenwood, S. The Role of Winter Precipitation as a Climatic Driver of the Spring Phenology of Five California Quercus Species (Fagaceae). Madroño 2021, 68, 450–460. [Google Scholar] [CrossRef]
- Papper, P.D.; Ackerly, D.D. Partitioning Genetic and Environmental Components of Phenological Variation in Quercus douglasii (Fagaceae). Madroño 2021, 68, 425–433. [Google Scholar] [CrossRef]
- Gerst, K.L.; Rossington, N.L.; Mazer, S.J. Phenological Responsiveness to Climate Differs among Four Species of Quercus in North America. J. Ecol. 2017, 105, 1610–1622. [Google Scholar] [CrossRef]
- Pardee, G.L.; Jensen, I.O.; Inouye, D.W.; Irwin, R.E. The Individual and Combined Effects of Snowmelt Timing and Frost Exposure on the Reproductive Success of Montane Forbs. J. Ecol. 2019, 107, 1970–1981. [Google Scholar] [CrossRef]
- Zhang, F.; Biederman, J.A.; Dannenberg, M.P.; Yan, D.; Reed, S.C.; Smith, W.K. Five Decades of Observed Daily Precipitation Reveal Longer and More Variable Drought Events Across Much of the Western United States. Geophys. Res. Lett. 2021, 48, e2020GL092293. [Google Scholar] [CrossRef]
- Bowers, J.E. Has Climatic Warming Altered Spring Flowering Date of Sonoran Desert Shrubs? Southwest. Nat. 2007, 52, 347–355. [Google Scholar] [CrossRef]
- Crimmins, T.M.; Crimmins, M.A.; David Bertelsen, C. Complex Responses to Climate Drivers in Onset of Spring Flowering across a Semi-arid Elevation Gradient. J. Ecol. 2010, 98, 1042–1051. [Google Scholar] [CrossRef]
- Waser, N.M. Pollinator Availability as a Determinant of Flowering Time in Ocotillo (Fouquieria splendens). Oecologia 1979, 39, 107–121. [Google Scholar] [CrossRef]
- Chuine, I. Why Does Phenology Drive Species Distribution? Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3149–3160. [Google Scholar] [CrossRef]
- Visser, M.E.; Noordwijk, A.J.V.; Tinbergen, J.M.; Lessells, C.M. Warmer Springs Lead to Mistimed Reproduction in Great Tits (Parus major). Proc. R. Soc. Lond. B Biol. Sci. 1998, 265, 1867–1870. [Google Scholar] [CrossRef]
- Inouye, D.W. Effects of Climate Change on Phenology, Frost Damage, and Floral Abundance of Montane Wildflowers. Ecology 2008, 89, 353–362. [Google Scholar] [CrossRef]
- Rafferty, N.E.; Diez, J.M.; Bertelsen, C.D. Changing Climate Drives Divergent and Nonlinear Shifts in Flowering Phenology across Elevations. Curr. Biol. 2020, 30, 432–441.e3. [Google Scholar] [CrossRef]
- Iler, A.M.; Compagnoni, A.; Inouye, D.W.; Williams, J.L.; CaraDonna, P.J.; Anderson, A.; Miller, T.E.X. Reproductive Losses Due to Climate Change-induced Earlier Flowering Are Not the Primary Threat to Plant Population Viability in a Perennial Herb. J. Ecol. 2019, 107, 1931–1943. [Google Scholar] [CrossRef]
- Wei, J.; Jin, Q.; Yang, Z.; Dirmeyer, P.A. Role of Ocean Evaporation in California Droughts and Floods. Geophys. Res. Lett. 2016, 43, 6554–6562. [Google Scholar] [CrossRef]
- Hu, F.; Zhang, L.; Liu, Q.; Chyi, D. Environmental Factors Controlling the Precipitation in California. Atmosphere 2021, 12, 997. [Google Scholar] [CrossRef]
- Forister, M.L.; Shapiro, A.M. Climatic Trends and Advancing Spring Flight of Butterflies in Lowland California. Glob. Change Biol. 2003, 9, 1130–1135. [Google Scholar] [CrossRef]
- Thomas, C.D.; Singer, M.C.; Boughton, D.A. Catastrophic Extinction of Population Sources in a Butterfly Metapopulation. Am. Nat. 1996, 148, 957–975. [Google Scholar] [CrossRef]
- Halsch, C.A.; Shapiro, A.M.; Fordyce, J.A.; Nice, C.C.; Thorne, J.H.; Waetjen, D.P.; Forister, M.L. Insects and Recent Climate Change. Proc. Natl. Acad. Sci. USA 2021, 118, e2002543117. [Google Scholar] [CrossRef]
- Augspurger, C.K. Spring 2007 Warmth and Frost: Phenology, Damage and Refoliation in a Temperate Deciduous Forest. Funct. Ecol. 2009, 23, 1031–1039. [Google Scholar] [CrossRef]
- Sillett, S.C.; Antoine, M.E.; Carroll, A.L.; Graham, M.E.; Chin, A.R.O.; Van Pelt, R. Rangewide Climatic Sensitivities and Non-Timber Values of Tall Sequoia sempervirens Forests. For. Ecol. Manag. 2022, 526, 120573. [Google Scholar] [CrossRef]
- Rosbakh, S.; Hartig, F.; Sandanov, D.V.; Bukharova, E.V.; Miller, T.K.; Primack, R.B. Siberian Plants Shift Their Phenology in Response to Climate Change. Glob. Change Biol. 2021, 27, 4435–4448. [Google Scholar] [CrossRef]
- Augspurger, C.K. Reconstructing Patterns of Temperature, Phenology, and Frost Damage over 124 Years: Spring Damage Risk Is Increasing. Ecology 2013, 94, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Gorchov, D.L. Pattern, Adaptation, and Constraint in Fruiting Synchrony within Vertebrate-Dispersed Woody Plants. Oikos 1990, 58, 169. [Google Scholar] [CrossRef]
- Saldivar, J.A.; Rankin, D.T.; Wilson-Rankin, E.E. Sites Dominated by Common Fiddleneck (Amsinckia menziesii var. intermedia) Support Diverse Plant-Pollinator Interactions. Plant Ecol. 2024, 225, 535–542. [Google Scholar] [CrossRef]
- Colautti, R.I.; Eckert, C.G.; Barrett, S.C.H. Evolutionary Constraints on Adaptive Evolution during Range Expansion in an Invasive Plant. Proc. R. Soc. B Biol. Sci. 2010, 277, 1799–1806. [Google Scholar] [CrossRef]
- Haggerty, B.P.; Galloway, L.F. Response of Individual Components of Reproductive Phenology to Growing Season Length in a Monocarpic Herb. J. Ecol. 2011, 99, 242–253. [Google Scholar] [CrossRef]
- Munguía-Rosas, M.A.; Ollerton, J.; Parra-Tabla, V.; De-Nova, J.A. Meta-Analysis of Phenotypic Selection on Flowering Phenology Suggests That Early Flowering Plants Are Favoured: Phenotypic Selection on Flowering Phenology. Ecol. Lett. 2011, 14, 511–521. [Google Scholar] [CrossRef]
- Lustenhouwer, N.; Wilschut, R.A.; Williams, J.L.; Van Der Putten, W.H.; Levine, J.M. Rapid Evolution of Phenology during Range Expansion with Recent Climate Change. Glob. Change Biol. 2018, 24, e534–e544. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solakis-Tena, A.; Hidalgo-Triana, N.; Boynton, R.; Thorne, J.H. Phenological Shifts Since 1830 in 29 Native Plant Species of California and Their Responses to Historical Climate Change. Plants 2025, 14, 843. https://doi.org/10.3390/plants14060843
Solakis-Tena A, Hidalgo-Triana N, Boynton R, Thorne JH. Phenological Shifts Since 1830 in 29 Native Plant Species of California and Their Responses to Historical Climate Change. Plants. 2025; 14(6):843. https://doi.org/10.3390/plants14060843
Chicago/Turabian StyleSolakis-Tena, Andros, Noelia Hidalgo-Triana, Ryan Boynton, and James H. Thorne. 2025. "Phenological Shifts Since 1830 in 29 Native Plant Species of California and Their Responses to Historical Climate Change" Plants 14, no. 6: 843. https://doi.org/10.3390/plants14060843
APA StyleSolakis-Tena, A., Hidalgo-Triana, N., Boynton, R., & Thorne, J. H. (2025). Phenological Shifts Since 1830 in 29 Native Plant Species of California and Their Responses to Historical Climate Change. Plants, 14(6), 843. https://doi.org/10.3390/plants14060843