
Physiological and Biochemical Adaptations to Repeated Drought–Rehydration Cycles in Ochroma lagopus Swartz: Implications for Growth and Stress Resilience


Abstract
1. Introduction
2. Results
2.1. Phenotypic Changes in O. lagopus Seedlings as the Number of Drought–Rehydration Cycles Increases
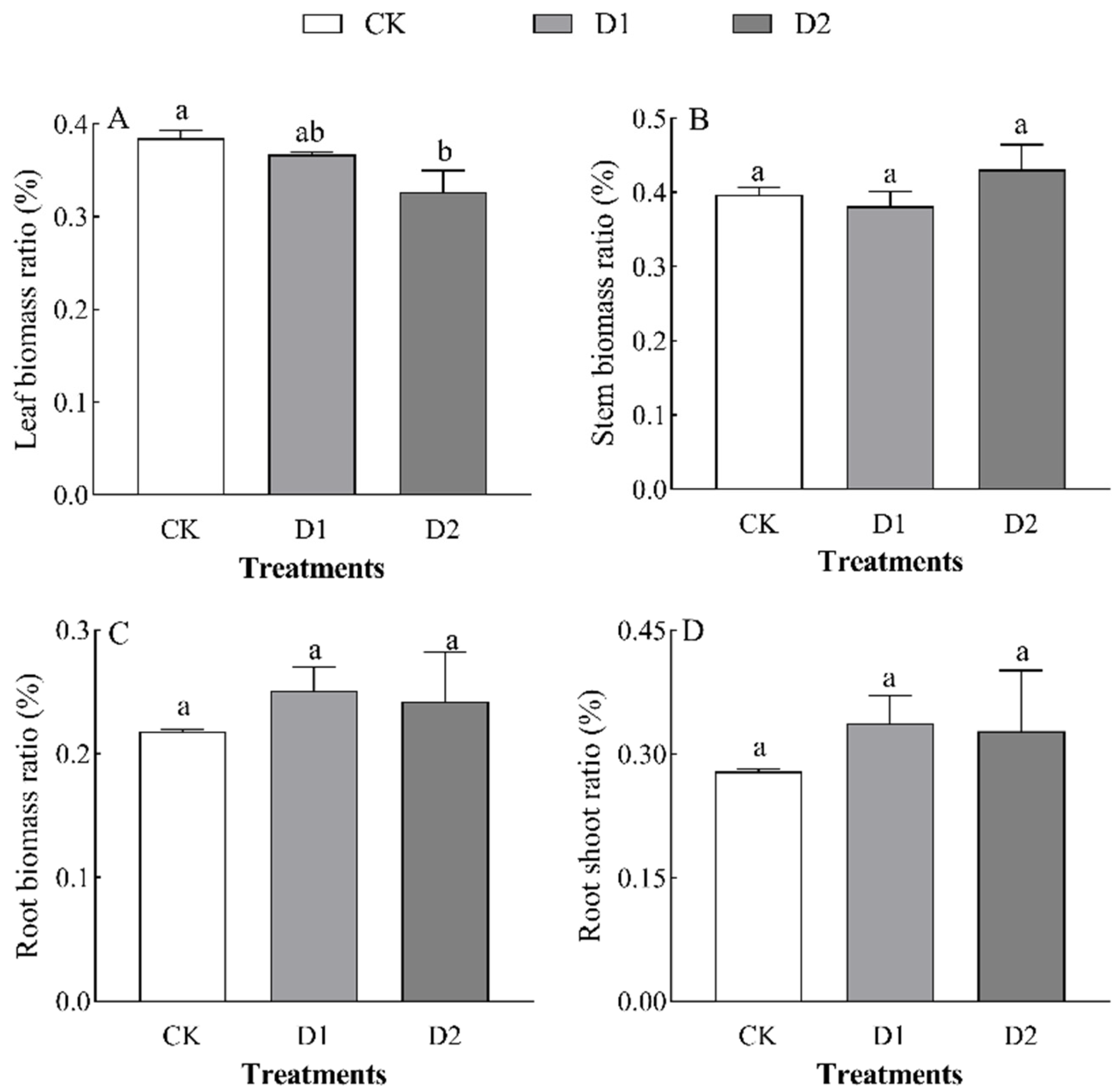
2.2. Effects of Increases in the Number of Drought–Rehydration Cycles on Biomass Changes and the Distribution of O. lagopus Seedlings
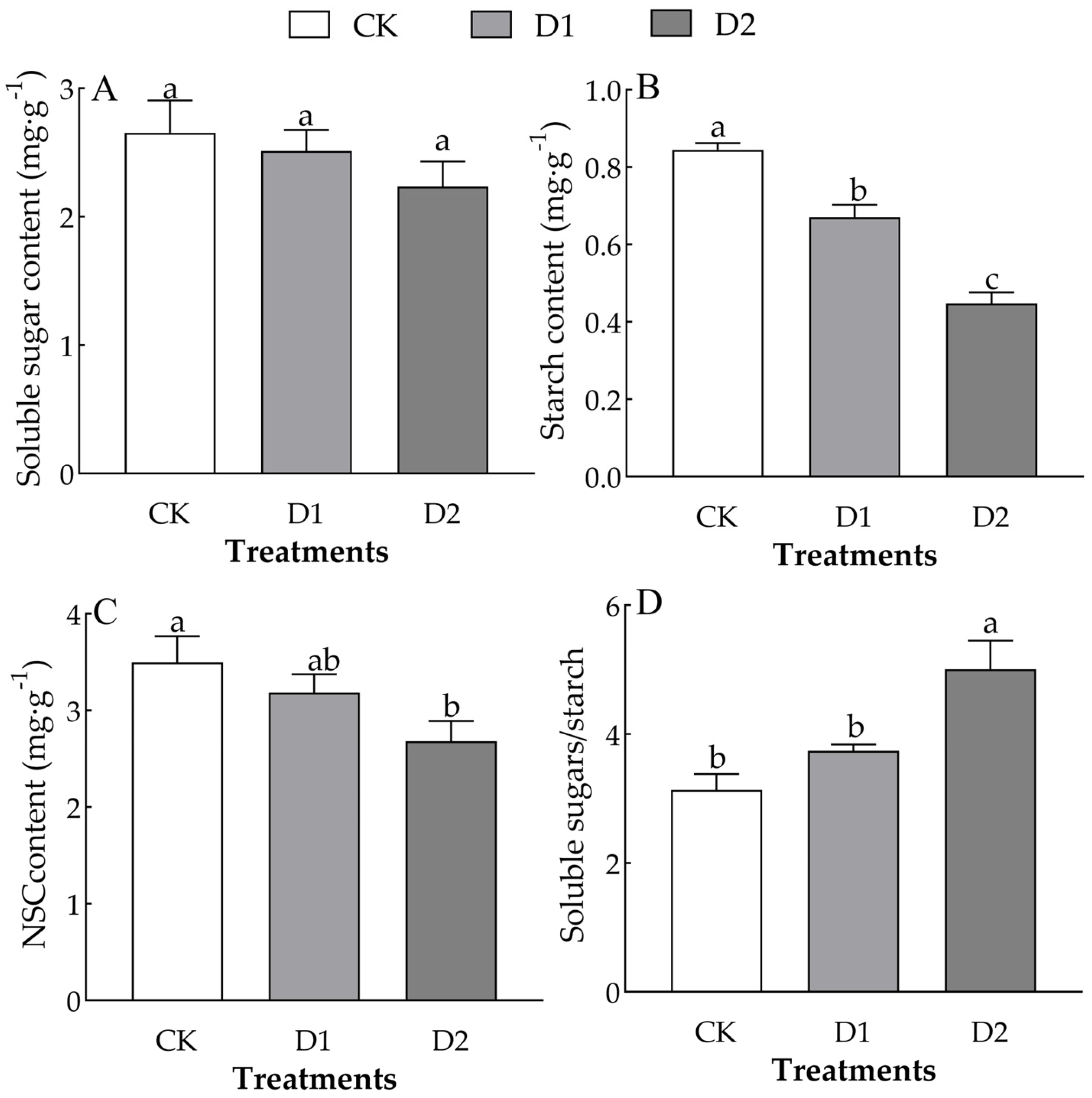
2.3. Changes in the NSCs of O. lagopus Leaves as the Number of Drought–Rehydration Cycles Increases
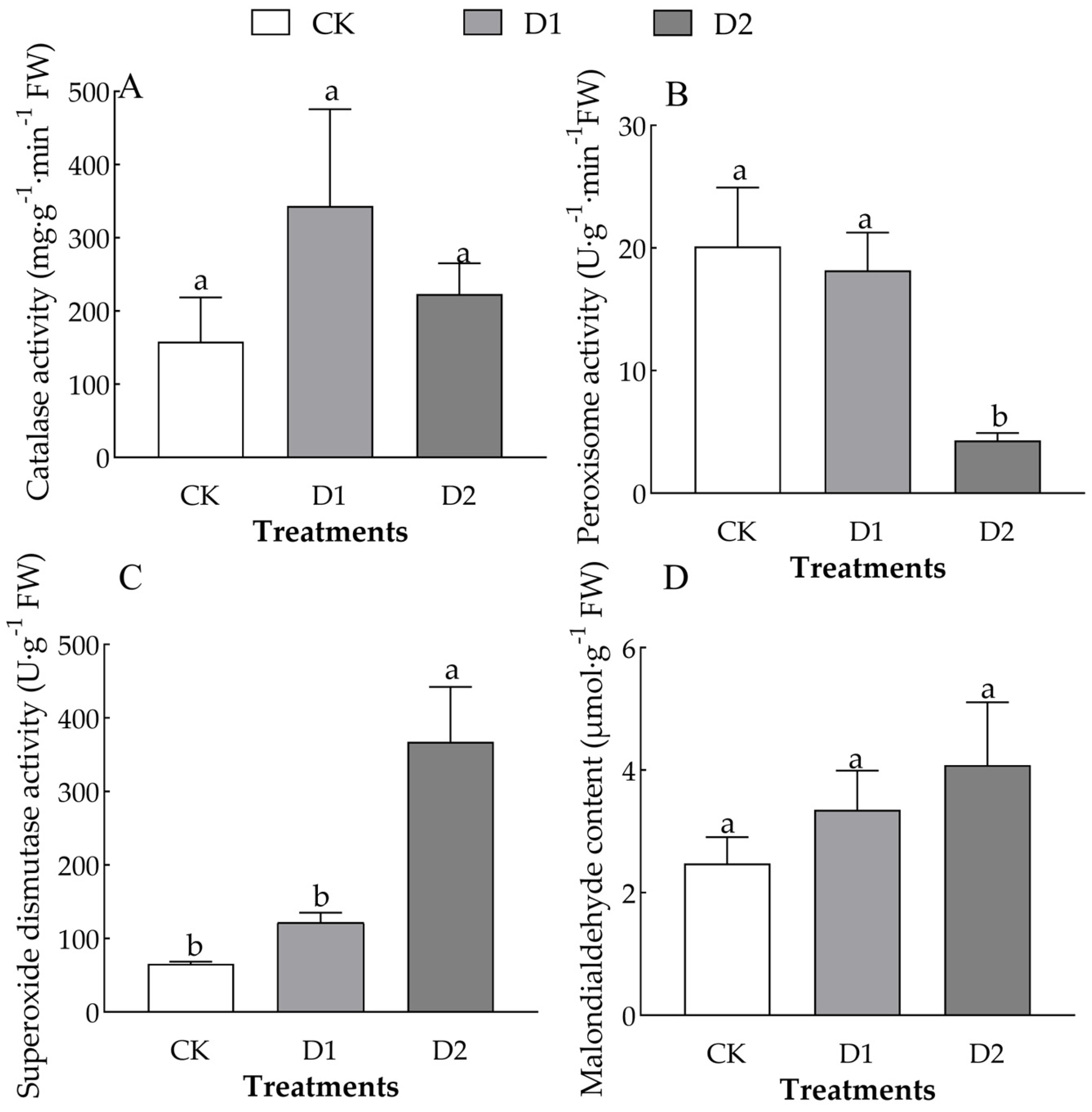
2.4. Changes in the MDA Content and Peroxidase Enzyme Activity of O. lagopus Needles as the Number of Drought–Rehydration Cycles Increases
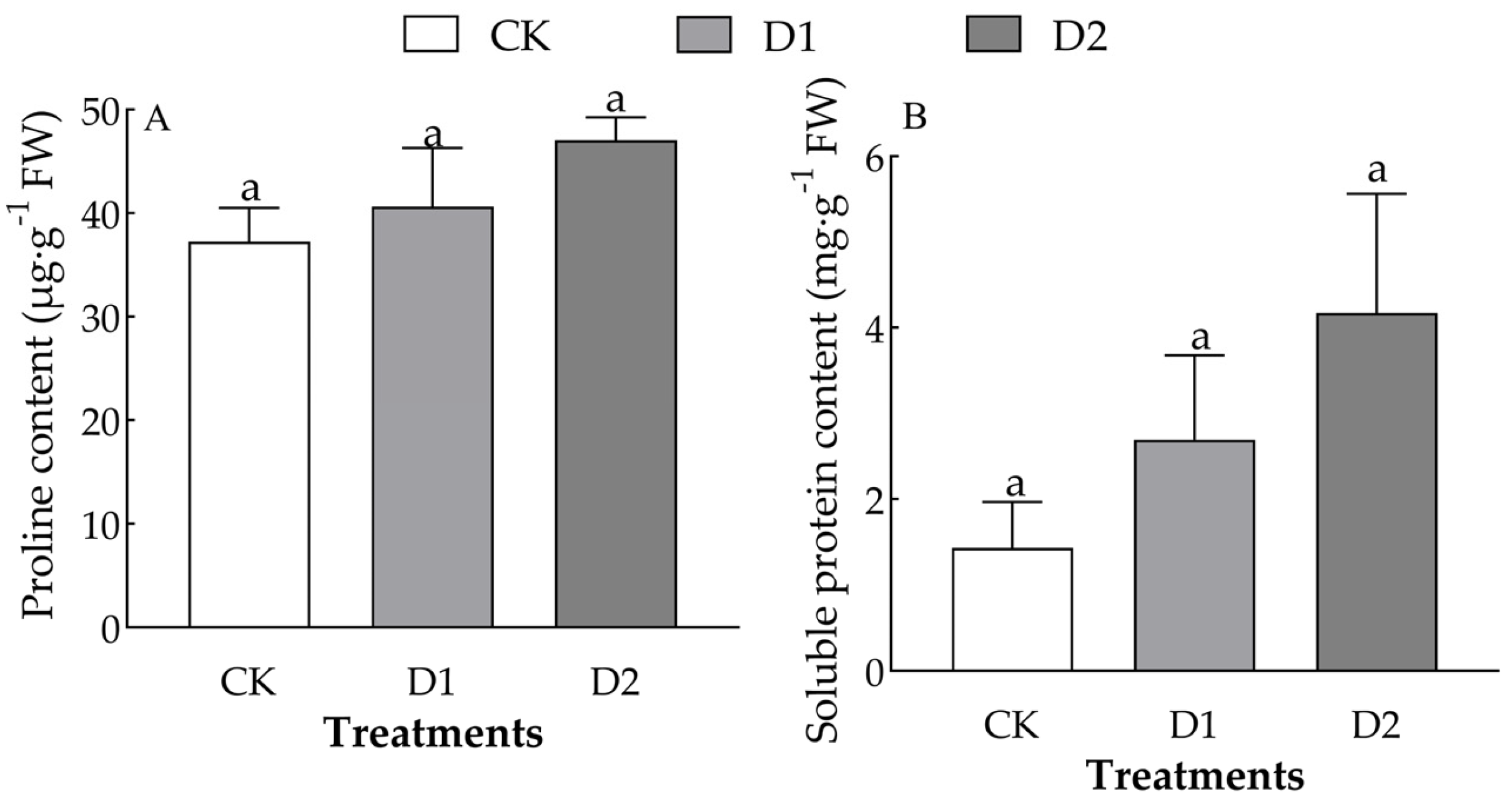
2.5. Effect of Increases in the Number of Drought–Rehydration Cycles on the Proline and Soluble Protein Content of O. lagopus Leaves
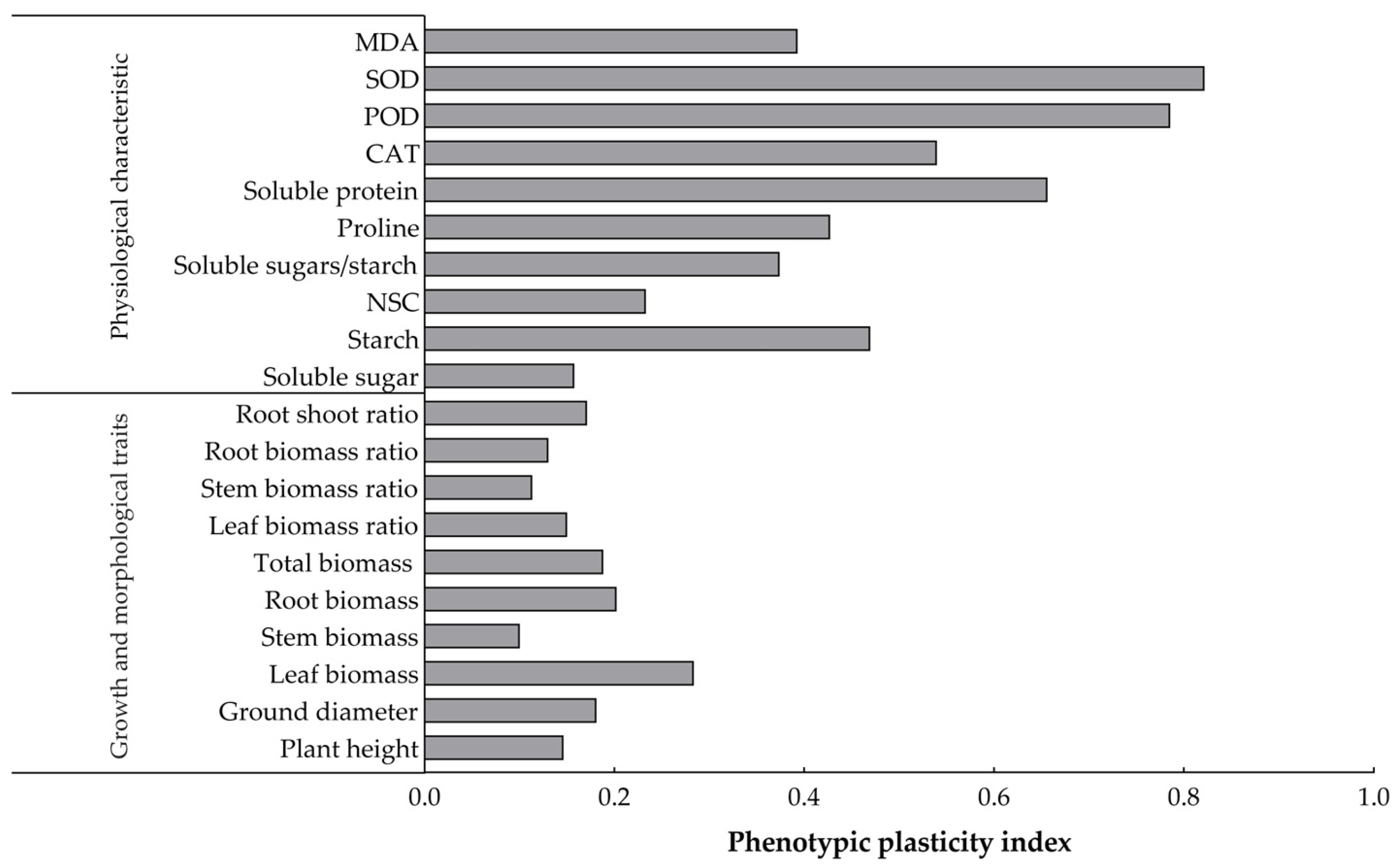
2.6. Plasticity Analysis of the Effects of Increases in the Number of Drought–Rehydration Cycles on the Growth and Leaf Physiological and Biochemical Indexes of O. lagopus Seedlings
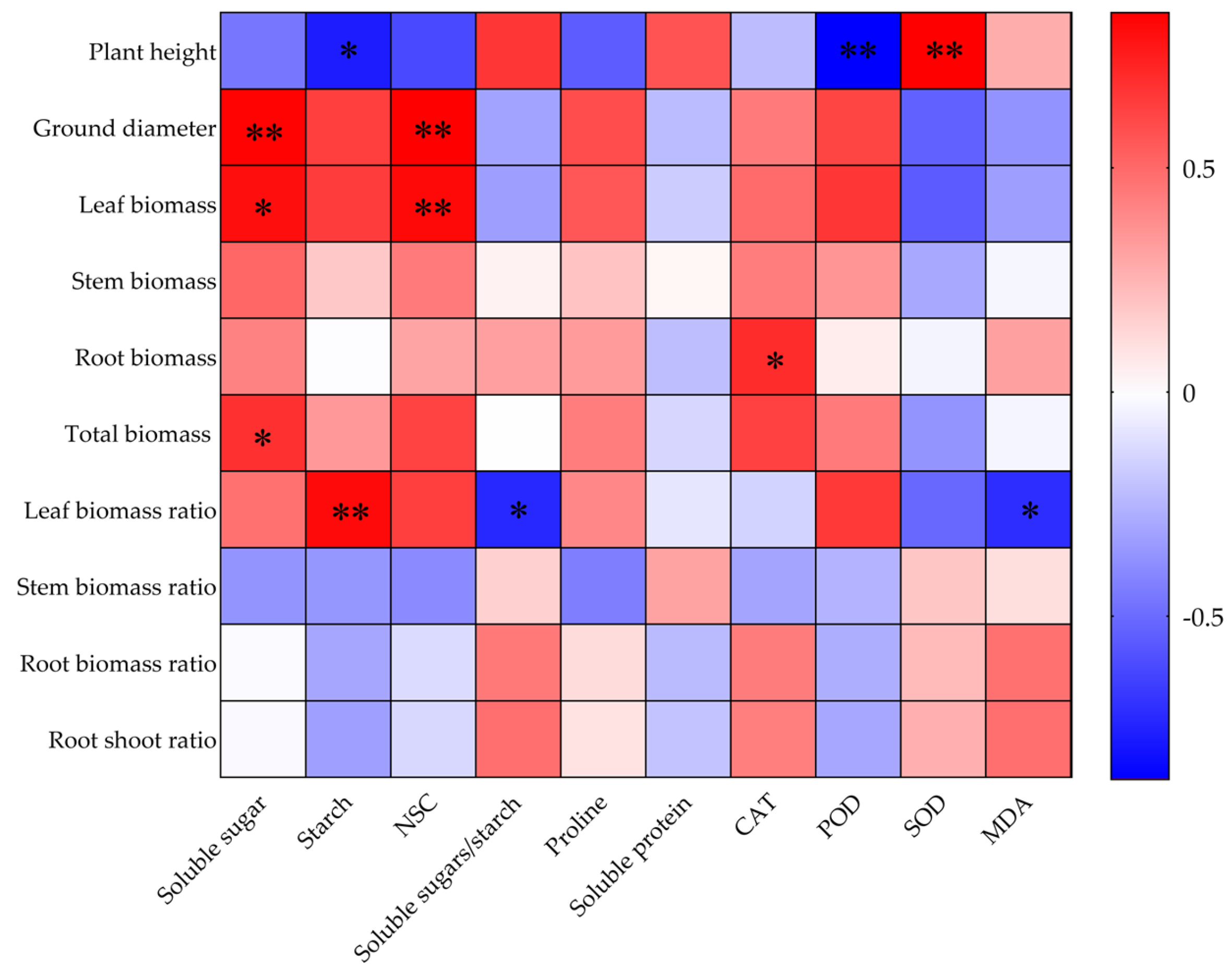
2.7. Correlations Between Growth Indices and Leaf Physiological and Biochemical Indices of O. lagopus Seedlings as the Number of Drought–Rehydration Cycles Increases
3. Discussion
3.1. Effects of Increases in the Number of Drought–Rehydration Cycles on the Growth and Biomass of O. lagopus Seedlings
3.2. Effects of Different Numbers of Drought–Rehydration Cycles on Leaf Physiology and Biochemistry of O. lagopus Leaves
3.3. Plasticity Analysis of the Growth and Physiological and Biochemical Indices of O. lagopus Seedlings in Response to Different Numbers of Drought–Rehydration Cycles
4. Materials and Methods
4.1. Study Area
4.2. Plant Material and Experimental Design
4.3. Plant Sampling
4.4. Determination of Leaf Physiological and Biochemical Indexes
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Anderegg, W.R.I.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Ashraf, U.; Zohaib, A.; Mohsin, T.; Muhammad, N.; Iftikhar, A.; Tahira, T.; Usman, N. Growth and developmental responses of crop plants under drought stress: A review. Agric. Plant Sci. 2017, 104, 267–276. [Google Scholar] [CrossRef]
- Liu, Y.; Xiao, J.; Sun, J.; Zhao, Z.; Deng, X.; Wu, J.; Zhang, D.; Bao, Y. Seasonal variation in C:N:P stoichiometry, nonstructural carbohydrates, and carbon isotopes of two coniferous pioneer tree species in subtropical China. Front. Plant Sci. 2023, 27, 1225436. [Google Scholar] [CrossRef] [PubMed]
- Ingram, J.; Bartels, D. The molecular basis of dehydration tolerance in plants. Annu. Rev. Plant Biol. 1996, 47, 377–403. [Google Scholar] [CrossRef]
- Rodrigues, M.L.; Pacheco, C.M.A.; Chaves, M.M. Soil-Plant water relations, root distribution and biomass partitioning in Lupinus albus L. under drought conditions. J. Exp. Bot. 1995, 46, 947–956. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; John, S.; Adam, W.; David, G.W.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Kofidis, G.; Bosabalidis, A.M.; Chartzoulakis, K. Leaves anatomical alterations induced by drought stress in two avocado cultivars. J. Biol. Res. 2004, 1, 115–120. [Google Scholar]
- Gebauer, R.; Plichta, R.; Urban, J.; Daniel, V.; Martina, H. The resistance and resilience of European beech seedlings to drought stress during the period of leaves development. Tree Physiol. 2020, 40, 1147–1164. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-Inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef]
- Fife, D.N.; Nambiar, E.K.S.; Saur, E. Retranslocation of foliar nutrients in evergreen tree species planted in a Mediterranean environment. Tree Physiol. 2008, 28, 187–196. [Google Scholar] [CrossRef]
- Abid, M.; Ali, S.; Qi, L.K.; Rizwan, Z.; Tian, Z.W.; Dong, J.L.; Snider, T.D. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef] [PubMed]
- Tian, K.; Wang, Y.; Chen, D.; Cao, M.; Jie, L. Influence of Drought Stress and Post-Drought Rewatering on Phytoremediation Effect of Arabidopsis thaliana. Bull. Environ. Contam. Toxicol. 2022, 108, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Furlan, A.; Bianucci, E.; Carmen, D.; Tordable, M.; Kleinert, A.; Valentine, A.; Castro, S. Dynamic responses of photosynthesis and the antioxidant system during a drought and rehydration cycle in peanut plants. Funct. Plant Biol. 2016, 43, 337–345. [Google Scholar] [CrossRef]
- Ilyas, M.; Nisar, M.; Khan, N.; Ali, H.; Aamir, H.K.; Kashif, H.; Shah, F.; Aziz, K.; Abid, U. Drought Tolerance Strategies in Plants: A Mechanistic Approach. J. Plant Growth Regul. 2021, 40, 926–944. [Google Scholar] [CrossRef]
- Liu, F.; Zhao, Y.; Wang, X.; Wang, B.; Bao, F.; He, K. Physiological response and drought resistance evaluation of Gleditsia sinensis seedlings under drought-rehydration state. Sci. Rep. 2023, 13, 19963. [Google Scholar] [CrossRef]
- Aleem, M.; Raza, M.M.; Haider, M.S.; Atif, R.M.; Ali, Z.; Bhat, J.A. Comprehensive RNA-seq analysis revealed molecular pathways and genes associated with drought tolerance in wild soybean (Glycine soja Sieb. and Zucc.). Physiol. Plant. 2021, 172, 703–728. [Google Scholar] [CrossRef]
- Rabisa, Z.; Muhammad, S.N.; Muhammad, J.S.; Sughra, H.; Asma, I. Plant survival under drought stress: Implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef]
- Shao, C.; Duan, H.; Ding, G.; Luo, X.; Fu, Y.; Lou, Q. Physiological and biochemical dynamics of Pinus massoniana Lamb. seedlings under extreme drought stress and during recovery. Forests 2022, 13, 65. [Google Scholar] [CrossRef]
- Sun, C.; Gao, X.; Chen, X.; Fu, J.; Zhang, Y. Metabolic and growth responses of maize to successive drought and re-watering cycles. Agric. Water Manag. 2016, 172, 62–73. [Google Scholar] [CrossRef]
- Cao, X.; Jia, J.; Zhang, C.; Li, H.; Liu, T.; Jiang, X.; Polle, A.; Peng, C.; Luo, Z.B. Anatomical, physiological and transcriptional responses of two contrasting poplar genotypes to drought and re-watering. Physiol. Plant. 2014, 151, 480–494. [Google Scholar] [CrossRef]
- Musembi, K.B.; Githiri, S.M.; Yencho, G.C.; Sibiya, J. Combining ability and heterosis for yield and drought tolerance traits under managed drought stress in sweetpotato. Euphytica 2015, 201, 423–440. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Gopi, R.; Sankar, B.; Muthiah, G.; Rajaram, P. Differential responsesin water use efficiency in two varieties of Catharanthus roseus under drought stress. CR Biol. 2008, 331, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.X.; Wu, J.W.; Jing, H.Q. Non-Structural carbohydrate (NSC) content and C:N:P stoichiometry of Pinus yunnanensis seedling needles in response to shade treatment. Ind. Crops Prod. 2024, 210, 118138. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.; Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Schiestl-Aalto, P.; Ryhti, K.; Mäkelä, A.; Peltoniemeim, M.; Back, J.; Kulmala, L. Analysis of the NSC storage dynamics in tree organs reveals the allocation to belowground symbionts in the framework of whole tree carbon balance. Front. For. Glob. Change 2019, 2, 17. [Google Scholar] [CrossRef]
- Ramírez-Briones, E.; Rodríguez-Macías, R.; Salcedo-Pérez, E.; Martínez-Gallardo, N.; Tiessen, A.; Molina-Torres, J.; Délano-Frier, J.P.; Zanudo, H. Seasonal variation in non-structural carbohydrates, sucrolytic activity and secondary metabolites in deciduous and perennial Diospyros species sampled in Western Mexico. PLoS ONE 2017, 12, e0187235. [Google Scholar] [CrossRef]
- Rosas, T.; Galiano, L.; Ogaya, R.; Peñuelas, J.; Martínez-Vilalta, J. Dynamics of non-structural carbohydrates in three Mediterranean woody species following long-term experimental drought. Front. Plant Sci. 2013, 4, 400. [Google Scholar] [CrossRef]
- Latt, C.R.; Nair, P.; Kang, B.T. Reserve carbohydrate levels in the boles and structural roots of five multipurpose tree species in a seasonally dry tropical climate. For. Ecol. Manag. 2001, 146, 145–158. [Google Scholar] [CrossRef]
- Malagat, B.; Iamba, K. Germination of O. lagopus Seedlings under Different Sowing Media in East New Britain, Papua New Guinea. Asian J. Res. Agric. For. 2021, 7, 73–88. [Google Scholar] [CrossRef]
- Fang, Y.J.; Xiong, L.Z. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Li, G.; Si, M.; Zhang, C.; Shen, Z.; Wang, S.; Shao, J. Responses of plant biomass and biomass allocation to experimental drought: A global phylogenetic meta-analysis. Agric. For. Meteorol. 2024, 347, 109917. [Google Scholar] [CrossRef]
- Ali, S.; Hayat, K.; Iqbal, A.; Xie, L.N. Implications of abscisic acid in the drought stress tolerance of plants. Agronomy 2020, 10, 1323. [Google Scholar] [CrossRef]
- Xu, B.; Li, F.; Shan, L.; Ma, Y.; Ichizen, N.; Huang, J. Gas exchange, biomass partition, and water relationships of three grass seedlings under water stress. Weed Biol. Manag. 2006, 6, 79–88. [Google Scholar] [CrossRef]
- Hultine, K.R.; Grady, K.C.; Wood, T.E.; Shuster, S.M.; Stella, J.C.; Whitham, T.G. Climate change perils for dioecious plant species. Nat. Plants 2016, 2, 16109. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Deng, X.; Xiao, W.; Shi, Z.; Zeng, L.; Lei, L. Combined Effects of Drought and Shading on Growth and Non-Structural Carbohydrates in Pinus massoniana Lamb. Seedlings. Forests 2019, 11, 18. [Google Scholar] [CrossRef]
- Wang, M.J.; Liang, W.W.; Fan, L.L.; Lu, H.L.; Lv, Y.B.; Zhou, S.; Zhang, L.L.; Ma, X. Physiological response of Vitis amurensis Rupr. Seedlings to drought stress and rewatering. IOP Conf. Ser. Earth Environ. Sci. 2021, 697, 012032. [Google Scholar] [CrossRef]
- Cao, H.; Wang, X.W.; Han, Z.H.; Xu, X.F.; Wang, Y.Z. Relationship between changes of endopeptidases activity and active oxygen in Malus hupehensis leaves during senescence induced by water stress. Sci. Agric. Sin. 2004, 37, 274–279. [Google Scholar]
- Sairam, R.K.; Saxena, D.C. Oxidative stress and antioxidants in wheat genotypes: A possible mechanism of water stress tolerance. J. Agron. Crop Sci. 2000, 184, 55–61. [Google Scholar] [CrossRef]
- Selote, D.S.; Khanna-Chopra, R. Drought acclimation confers oxidative stress tolerance by inducing coordinated antioxidant defense at cellular and subcellular level in leaves of wheat seedlings. Physiol. Plant. 2006, 127, 494–506. [Google Scholar] [CrossRef]
- Kim, E.J.; Choi, J.Y.; Yu, M.R.; Kim, M.Y. Total polyphenols, total flavonoid contents, and antioxidant activity of Korean natural and medicinal plants. Korean J. Food Sci. Technol. 2012, 44, 337–342. [Google Scholar] [CrossRef]
- Menezes-Silva, P.E.; Sanglard, L.M.; Ávila, R.T.; Morais, L.E.; Martins, S.C.V.; Nobres, P.; Patreze, C.M.; Ferreira, M.A.; Araújo, W.L.; Fernie, A.R.; et al. Photosynthetic and mefigolic acclimation to repeated drought events play key roles in drought tolerance in coffee. J. Exp. Bot. 2017, 68, 4309–4322. [Google Scholar] [CrossRef] [PubMed]
- Poorter, L.; Kitajima, K. Carbohydrate storage and light requirements of tropical moist and dry forest tree species. Ecology 2007, 88, 1000–1011. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.P.; Rodrigues, M.L.; Ricardo, C.P.; Oliveira, M.M.; Lopes, C.M. How plants cope with water stress in the field: Physiological and molecular mechanisms. Funct. Plant Biol. 2002, 29, 239–249. [Google Scholar]
- Guo, X.; Peng, C.; Li, T.; Huang, J.; Song, H.; Zhu, Q.; Wang, M. The Effects of Drought and Re-Watering on Non-Structural Carbohydrates of Pinus tabulaeformis Seedlings. Biology 2021, 10, 281. [Google Scholar] [CrossRef]
- Trifilò, P.; Kiorapostolou, N.; Petruzzellis, F.; Vitti, S.; Petit, G.; Maria, A.; Gullo, L.; Nardini, A.; Casolo, V. Hydraulic recovery from xylem embolism in excised branches of twelve woody species: Relationships with parenchyma cells and non-structural carbohydrates. Plant Physiol. Biochem. 2019, 139, 513–520. [Google Scholar] [CrossRef]
- Hassan, I.; Chen, Y.; Muhammad, W.; Syed, T.R.; Muhammad, S.; Zeeshan, A. Salinity and exogenous H2O2 improve gas exchange, osmoregulation, and antioxidant metabolism in quinoa under drought stress. Physiol. Plant. 2023, 175, e14057. [Google Scholar]
- Mu, Q.; Cai, H.J.; Sun, S.K.; Wen, S.; Xu, J.; Dong, M.; Saddique, Q. The physiological response of winter wheat under short-term drought conditions and the sensitivity of different indices to soil water changes. Agric. Water Manag. 2021, 243, 106475. [Google Scholar] [CrossRef]
- Dubey, R.S. Protein synthesis by plants under stressful conditions. Handb. Plant Crop Stress 1999, 2, 365–397. [Google Scholar]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, L.; Liu, Y.; Sun, J.; Xiao, J.; Dong, Q.; Li, L.; Zhang, W.; Wang, C.; Wu, J. Effects of drought on non-structural carbohydrates and C, N, and P stoichiometric characteristics of Pinus yunnanensis seedlings. J. For. Res. 2024, 35, 12. [Google Scholar] [CrossRef]
- Zhao, S.J. The Experimental Guide for Plant Physiology, 3rd ed.; Higher Education Press: Beijing, China, 2002. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Sancho, M.A.; De Forchetti, S.M.; Pliego, F. Peroxidase activity and isoenzymes in the culture medium of NaCl adapted tomato suspension cells. Plant Cell Tissue Organ Cult. 1996, 44, 161–167. [Google Scholar] [CrossRef]
- Lei, Y.; Yin, C.; Li, C. Differences in some morphological, physiological, and biochemical responses to drought stress in two contrasting populations of Populus przewalskii. Physiol. Plant. 2006, 127, 182–191. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height/cm | Ground Diameter/mm | ||||
|---|---|---|---|---|---|---|
| Before Treatment | After Treatment | Increment | Before Treatment | After Treatment | Increment | |
| CK | 23.32 ± 0.46 a | 57.67 ± 2.24 b | 34.37 ± 1.83 b | 6.93 ± 0.09 a | 11.40 ± 1.31 a | 4.47 ± 1.39 a |
| D1 | 23.14 ± 0.12 a | 58.47 ± 0.44 b | 35.37 ± 0.33 b | 6.43 ± 0.12 a | 11.27 ± 0.91 a | 4.83 ± 0.79 a |
| D2 | 22.91 ± 0.50 a | 67.57 ± 1.96 a | 44.67 ± 1.50 a | 6.46 ± 0.09 a | 9.33 ± 0.07 a | 3.11 ± 0.03 a |
| Indicators | F | p |
|---|---|---|
| Plant height (cm) | 10.048 | 0.012 |
| Ground diameter (mm) | 1.587 | 0.280 |
| Leaf biomass (g) | 3.116 | 0.118 |
| Stem biomass (g) | 0.211 | 0.815 |
| Root biomass (g) | 0.603 | 0.577 |
| Total biomass (g) | 1.019 | 0.416 |
| Leaf biomass ratio (%) | 4.373 | 0.067 |
| Stem biomass ratio (%) | 1.162 | 0.374 |
| Root biomass ratio (%) | 0.44 | 0.663 |
| Root/shoot ratio (%) | 0.438 | 0.664 |
| Soluble sugars (mg·g−1) | 1.06 | 0.404 |
| Starch (mg·g−1) | 55.494 | 0.000 |
| NSCs (mg·g−1) | 3.308 | 0.108 |
| Soluble sugars/starch | 10.27 | 0.012 |
| Proline (μg·g−1 FW) | 2.746 | 0.142 |
| Soluble protein (mg·g−1 FW) | 1.769 | 0.249 |
| CAT (μmol·g−1 FW) | 1.155 | 0.376 |
| POD U·g−1·min−1 FW | 6.756 | 0.029 |
| SOD (U·g−1 FW) | 13.475 | 0.006 |
| MDA (μmol·g−1 FW) | 1.18 | 0.370 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Sun, J.; Dai, C.; Du, G.; Shi, R.; Wu, J. Physiological and Biochemical Adaptations to Repeated Drought–Rehydration Cycles in Ochroma lagopus Swartz: Implications for Growth and Stress Resilience. Plants 2025, 14, 1636. https://doi.org/10.3390/plants14111636
Liu Y, Sun J, Dai C, Du G, Shi R, Wu J. Physiological and Biochemical Adaptations to Repeated Drought–Rehydration Cycles in Ochroma lagopus Swartz: Implications for Growth and Stress Resilience. Plants. 2025; 14(11):1636. https://doi.org/10.3390/plants14111636
Chicago/Turabian StyleLiu, Yuanxi, Jianli Sun, Cefeng Dai, Guanben Du, Rui Shi, and Junwen Wu. 2025. "Physiological and Biochemical Adaptations to Repeated Drought–Rehydration Cycles in Ochroma lagopus Swartz: Implications for Growth and Stress Resilience" Plants 14, no. 11: 1636. https://doi.org/10.3390/plants14111636
APA StyleLiu, Y., Sun, J., Dai, C., Du, G., Shi, R., & Wu, J. (2025). Physiological and Biochemical Adaptations to Repeated Drought–Rehydration Cycles in Ochroma lagopus Swartz: Implications for Growth and Stress Resilience. Plants, 14(11), 1636. https://doi.org/10.3390/plants14111636