
Do Abiotic Stresses Affect the Aroma of Damask Roses?

 , , ,
, , ,  and
and
Abstract
:1. Introduction
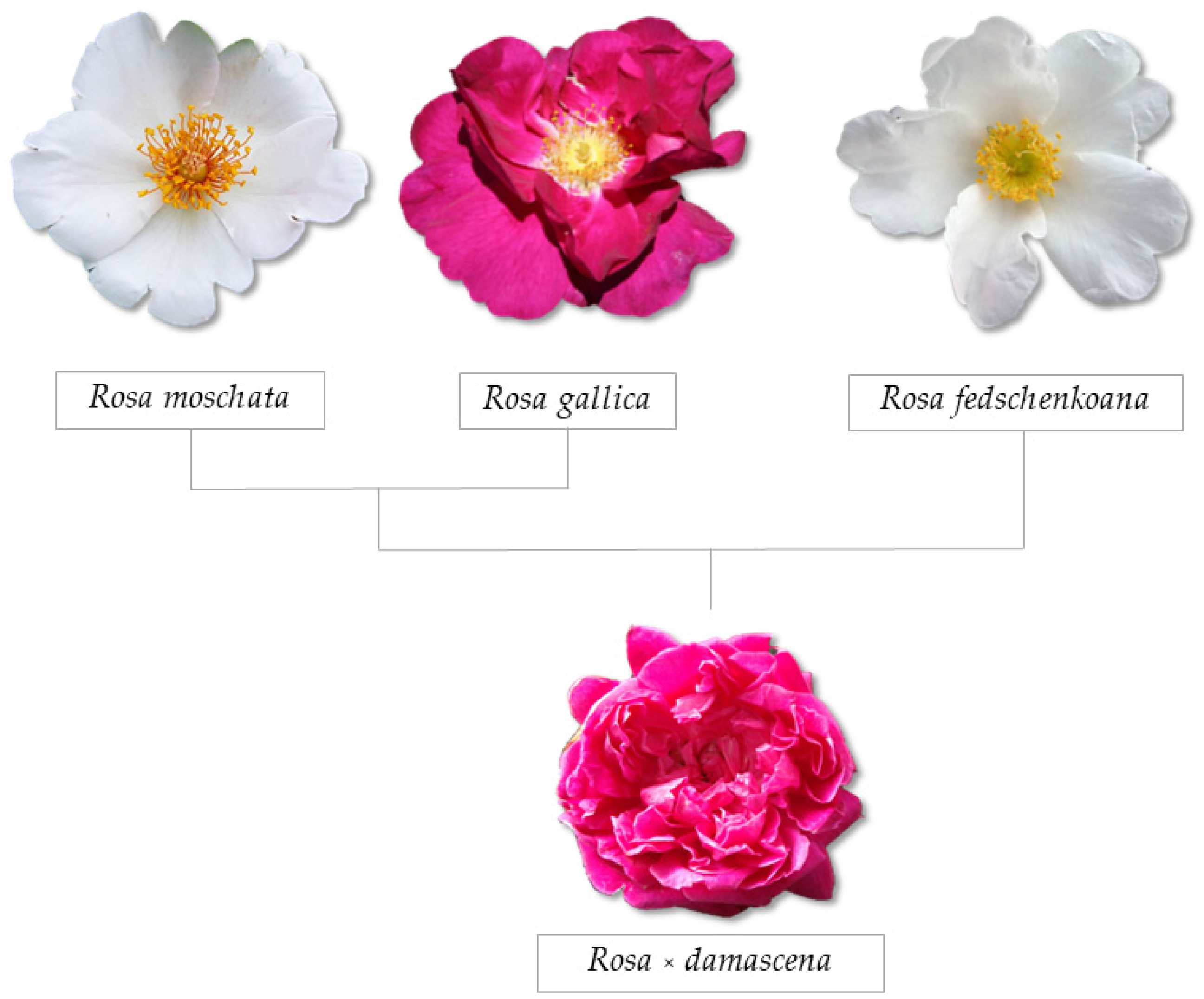
2. Taxonomy of Damask Rose
3. Biosynthesis of the Aromatic Volatiles in Damask Rose
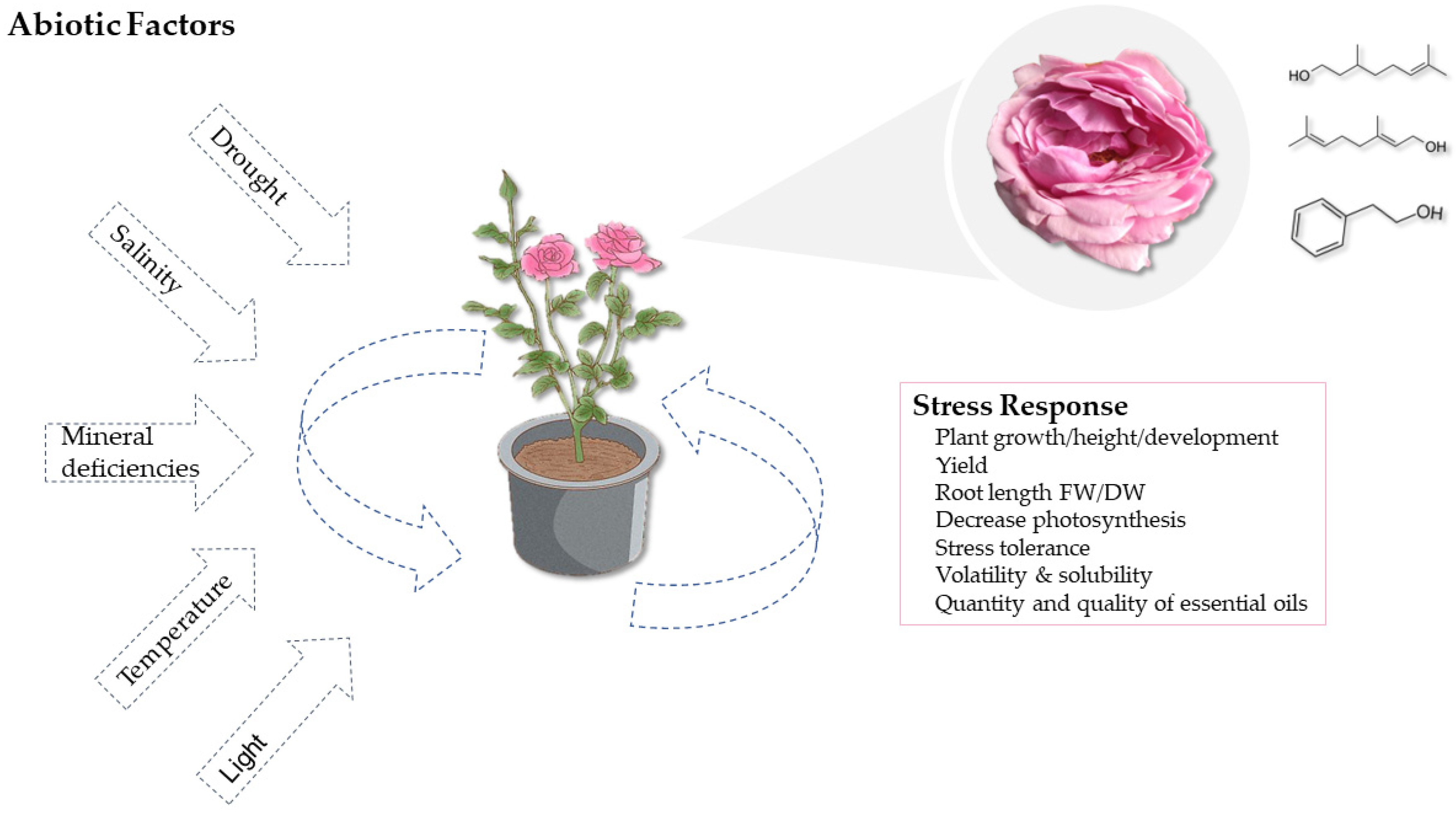
4. Agronomic Aspects Influencing the Biosynthesis of Aromatic Compounds in Roses
5. Influence of Abiotic Stressors on Aromatic Profile of Damask Rose
5.1. Drought Stress
5.2. Salt Stress
5.3. Nutrient Stress
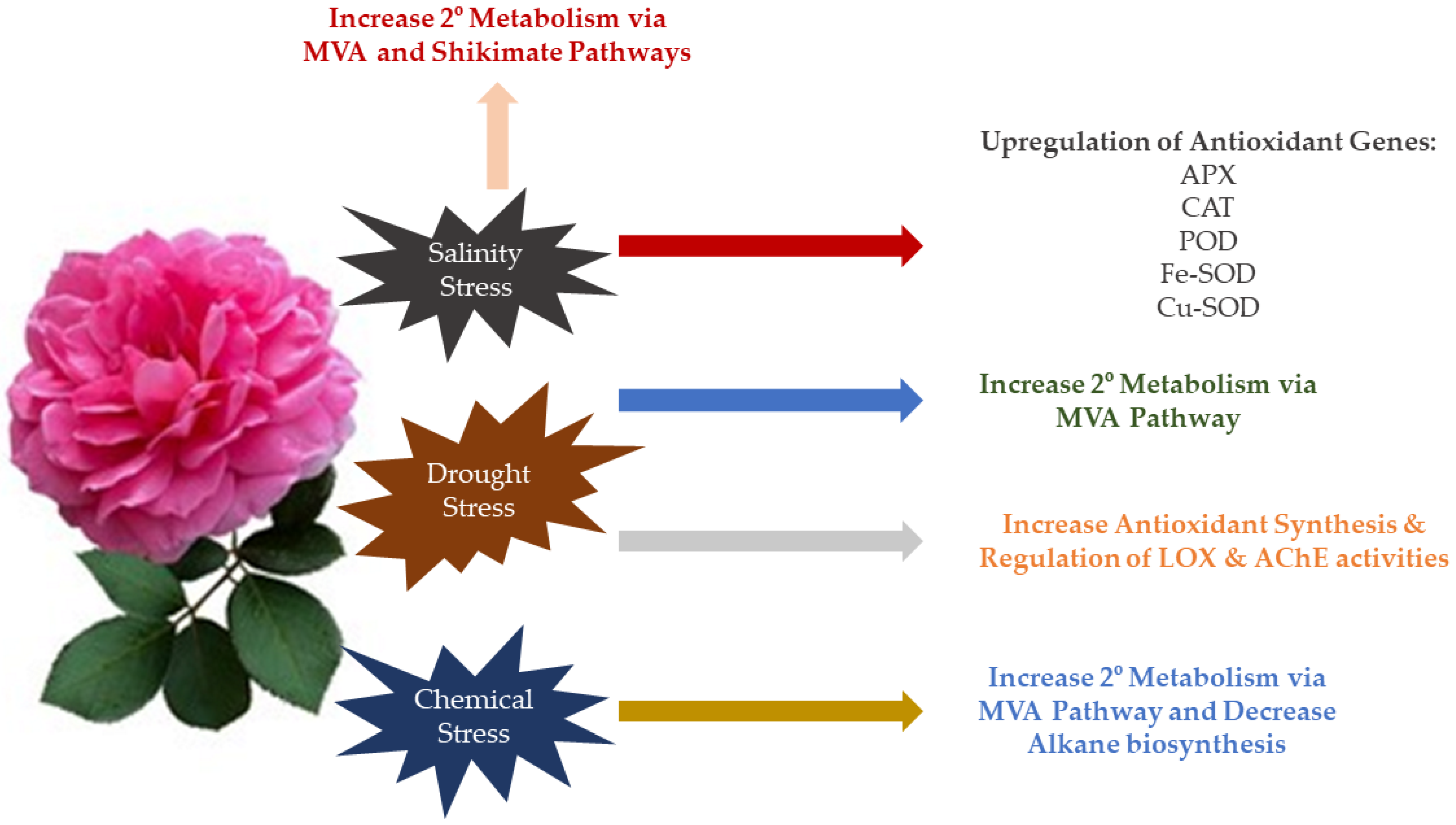
6. Molecular Bases of Abiotic Stress Response in Roses
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farooq, A.; Kiani, M.; Khan, M.A.; Riaz, A.; Khan, A.A.; Anderson, N.; Byrne, D.H. Microsatellite analysis of Rosa damascena from Pakistan and Iran. Hortic. Environ. Biotechnol. 2013, 54, 141–147. [Google Scholar] [CrossRef]
- Pal, P.K. Evaluation, genetic diversity, recent development of distillation method, challenges and opportunities of Rosa damascena: A review. J. Essent. Oil Bear. Plants 2013, 16, 1–10. [Google Scholar] [CrossRef]
- Naquvi, K.J.; Ansari, S.; Ali, M.; Najmi, A. Volatile oil composition of Rosa damascena Mill.(Rosaceae). J. Pharmacogn. Phytochem. 2014, 2, 117–181. [Google Scholar]
- Ghavam, M.; Afzali, A.; Manca, M.L. Chemotype of damask rose with oleic acid (9 octadecenoic acid) and its antimicrobial effectiveness. Sci. Rep. 2021, 11, 8027. [Google Scholar] [CrossRef]
- Verma, R.S.; Padalia, R.C.; Chauhan, A.; Singh, A.; Yadav, A.K. Volatile constituents of essential oil and rose water of damask rose (Rosa damascena Mill.) cultivars from North Indian hills. Nat. Prod. Res. 2011, 25, 1577–1584. [Google Scholar] [CrossRef]
- Kumar, R.; Sharma, S.; Sood, S.; Agnihotri, V.K.; Singh, B. Effect of diurnal variability and storage conditions on essential oil content and quality of damask rose (Rosa damascena Mill.) flowers in north western Himalayas. Sci. Hortic. 2013, 154, 102–108. [Google Scholar] [CrossRef]
- Balaj, N.; Prishtina, K. Garden Roses (Rosa x hybrida) as a landscape architecture plants: Large blooms and compact growth. In Proceedings of the Architecture and Spatial Planning, Pristina, Kosovo, 26–28 October 2019; p. 62. [Google Scholar]
- Verma, S.R.; Padalia, C.R.; Chauhan, A. Chemical investigation of the volatile components of shade-dried petals of damask rose (Rosa damascena Mill.). Arch. Biol. Sci. 2011, 63, 1111–1115. [Google Scholar] [CrossRef]
- Li, R.; Wang, K.; Wang, D.; Xu, L.; Shi, Y.; Dai, Z.; Zhang, X. Production of plant volatile terpenoids (rose oil) by yeast cell factories. Green Chem. 2021, 23, 5088–5096. [Google Scholar] [CrossRef]
- Kovacheva, N.; Rusanov, K.; Atanassov, I. Industrial cultivation of oil bearing rose and rose oil production in Bulgaria during 21st century, directions and challenges. Biotechnol. Biotechnol. Equip. 2010, 24, 1793–1798. [Google Scholar] [CrossRef]
- Anonymous. Global “Rose Essential Oil Market” 2019 Industry Research Report. Available online: https://www.grandviewresearch.com/industry-analysis/rose-oil-market (accessed on 23 February 2023).
- Kumar, R.; Sharma, S.; Kaundal, M.; Sood, S.; Agnihotri, V.K. Variation in essential oil content and composition of damask rose (Rosa damascena mill) flowers by salt application under mid hills of the western Himalayas. J. Essent. Oil Bear. Plants 2016, 19, 297–306. [Google Scholar] [CrossRef]
- Pal, P.K.; Mahajan, M. Pruning system and foliar application of MgSO4 alter yield and secondary metabolite profile of Rosa damascena under rainfed acidic conditions. Front. Plant Sci. 2017, 8, 507. [Google Scholar] [CrossRef] [PubMed]
- Fatemi, F.; Golbodagh, A.; Hojihosseini, R.; Dadkhah, A.; Akbarzadeh, K.; Salome, D.; Malayeri, M.R.M. Anti-inflammatory effects of deuterium-depleted water plus Rosa damascena mill. Essential oil via cyclooxygenase-2 pathway in rats. Turk. J. Pharm. Sci. 2020, 17, 99. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hameed, E.-S.S.; Bazaid, S.A.; Salman, M.S. Characterization of the phytochemical constituents of Taif rose and its antioxidant and anticancer activities. BioMed Res. Int. 2013, 2013, 345465. [Google Scholar] [CrossRef] [PubMed]
- Boskabady, M.H.; Shafei, M.N.; Saberi, Z.; Amini, S. Pharmacological effects of Rosa damascena. Iran. J. Basic Med. Sci. 2011, 14, 295. [Google Scholar] [PubMed]
- Bashir, A.; Ahmad, T.; Farooq, S.; Lone, W.I.; Manzoor, M.M.; Nalli, Y.; Sultan, P.; Chaubey, A.; Ali, A.; Riyaz-Ul-Hassan, S. A Secondary Metabolite of Cercospora sp., Associated with Rosa damascena Mill., Inhibits Proliferation, Biofilm Production, Ergosterol Synthesis and Other Virulence Factors in Candida albicans. Microb. Ecol. 2022, 85, 1276–1287. [Google Scholar] [CrossRef] [PubMed]
- Ghavam, M. Relationships of irrigation water and soil physical and chemical characteristics with yield, chemical composition and antimicrobial activity of Damask rose essential oil. PLoS ONE 2021, 16, e0249363. [Google Scholar] [CrossRef]
- Bertoli, A.; Fambrini, M.; Doveri, S.; Leonardi, M.; Pugliesi, C.; Pistelli, L. Pollen aroma fingerprint of two sunflower (Helianthus annuus L.) genotypes characterized by different pollen colors. Chem. Biodivers. 2011, 8, 1766–1775. [Google Scholar] [CrossRef]
- Tambe, E.; Gotmare, S. Study of variation and identification of chemical composition in Rosa species oil collected from different countries. IOSR J. Appl. Chem. 2016, 9, 11–18. [Google Scholar]
- Hirata, H.; Ohnishi, T.; Watanabe, N. Biosynthesis of floral scent 2-phenylethanol in rose flowers. Biosci. Biotechnol. Biochem. 2016, 80, 1865–1873. [Google Scholar] [CrossRef]
- Al-Yasi, H.; Attia, H.; Alamer, K.; Hassan, F.; Ali, E.; Elshazly, S.; Siddique, K.H.; Hessini, K. Impact of drought on growth, photosynthesis, osmotic adjustment, and cell wall elasticity in Damask rose. Plant Physiol. Biochem. 2020, 150, 133–139. [Google Scholar] [CrossRef]
- Kanani, M.; Chamani, E.; Shokouhian, A.A.; Torabi-Giglou, M. Plant secondary metabolism and flower color changes in damask rose at different flowering development stages. Acta Physiol. Plant. 2021, 43, 55. [Google Scholar] [CrossRef]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Amiri, R.; Nikbakht, A.; Etemadi, N. Alleviation of drought stress on rose geranium [Pelargonium graveolens (L.) Herit.] in terms of antioxidant activity and secondary metabolites by mycorrhizal inoculation. Sci. Hortic. 2015, 197, 373–380. [Google Scholar] [CrossRef]
- Joo, S.S.; Kim, Y.B.; Lee, D.I. Antimicrobial and antioxidant properties of secondary metabolites from white rose flower. Plant Pathol. J. 2010, 26, 57–62. [Google Scholar] [CrossRef]
- Riahi, L.; Cherif, H.; Miladi, S.; Neifar, M.; Bejaoui, B.; Chouchane, H.; Masmoudi, A.S.; Cherif, A. Use of plant growth promoting bacteria as an efficient biotechnological tool to enhance the biomass and secondary metabolites production of the industrial crop Pelargonium graveolens L’Hér. under semi-controlled conditions. Ind. Crops Prod. 2020, 154, 112721. [Google Scholar] [CrossRef]
- Picazo-Aragonés, J.; Terrab, A.; Balao, F. Plant volatile organic compounds evolution: Transcriptional regulation, epigenetics and polyploidy. Int. J. Mol. Sci. 2020, 21, 8956. [Google Scholar] [CrossRef]
- Zhou, L.; Yu, C.; Cheng, B.; Wan, H.; Luo, L.; Pan, H.; Zhang, Q. Volatile compound analysis and aroma evaluation of tea-scented roses in China. Ind. Crops Prod. 2020, 155, 112735. [Google Scholar] [CrossRef]
- Tholl, D.; Hossain, O.; Weinhold, A.; Röse, U.S.; Wei, Q. Trends and applications in plant volatile sampling and analysis. Plant J. 2021, 106, 314–325. [Google Scholar] [CrossRef]
- Cornelissen, T. Climate change and its effects on terrestrial insects and herbivory patterns. Neotrop. Entomol. 2011, 40, 155–163. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, W.; Sun, L.; Zhao, F.; Huang, B.; Yang, W.; Tao, Y.; Wang, J.; Yuan, Z.; Fan, G. The genome of Prunus mume. Nat. Commun. 2012, 3, 1318. [Google Scholar] [CrossRef]
- Soundararajan, P.; Won, S.Y.; Kim, J.S. Insight on Rosaceae family with genome sequencing and functional genomics perspective. BioMed Res. Int. 2019, 2019, 7519687. [Google Scholar] [CrossRef] [PubMed]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B.; Gareth Jones, E.; Tibpromma, S.; Tennakoon, D.S.; Dissanayake, A.J.; Jayasiri, S.C. Fungal diversity notes 709–839: Taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Haider, N. A brief review on plant taxonomy and its components. J. Plant Sci. Res. 2018, 34, 277–292. [Google Scholar] [CrossRef]
- Farinati, S.; Rasori, A.; Varotto, S.; Bonghi, C. Rosaceae fruit development, ripening and post-harvest: An epigenetic perspective. Front. Plant Sci. 2017, 8, 1247. [Google Scholar] [CrossRef] [PubMed]
- Mimida, N.; Li, J.; Zhang, C.; Moriya, S.; Moriya-Tanaka, Y.; Iwanami, H.; Honda, C.; Oshino, H.; Takagishi, K.; Suzuki, A. Divergence of TERMINAL FLOWER1-like genes in Rosaceae. Biol. Plant. 2012, 56, 465–472. [Google Scholar] [CrossRef]
- Veluru, A. Genetic Diversity Analysis of Rose (Rosa X hybrida L.) Cultivars based on Morphological Markers. Int. J. Curr. Microbiol. App. Sci 2021, 10, 1349–1355. [Google Scholar] [CrossRef]
- Hummer, K.E.; Janick, J. Rosaceae: Taxonomy, Economic Importance, Genomics. In Genetics and Genomics of Rosaceae; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–17. [Google Scholar]
- Ritz, C.; Schmuths, H.; Wissemann, V. Evolution by reticulation: European dogroses originated by multiple hybridization across the genus Rosa. J. Hered. 2005, 96, 4–14. [Google Scholar] [CrossRef]
- Venkatesha, K.; Gupta, A.; Rai, A.N.; Jambhulkar, S.J.; Bisht, R.; Padalia, R.C. Recent developments, challenges, and opportunities in genetic improvement of essential oil-bearing rose (Rosa damascena): A review. Ind. Crops Prod. 2022, 184, 114984. [Google Scholar] [CrossRef]
- Mileva, M.; Ilieva, Y.; Jovtchev, G.; Gateva, S.; Zaharieva, M.M.; Georgieva, A.; Dimitrova, L.; Dobreva, A.; Angelova, T.; Vilhelmova-Ilieva, N. Rose flowers—A delicate perfume or a natural healer? Biomolecules 2021, 11, 127. [Google Scholar] [CrossRef]
- Nunes, H.; Miguel, M.G. Rosa damascena essential oils: A brief review about chemical composition and biological properties. Trends Phytochem. Res. 2017, 1, 111–128. [Google Scholar]
- Labban, L.; Thallaj, N. The medicinal and pharmacological properties of Damascene Rose (Rosa damascena ): A review. Int. J. Herb. Med. 2020, 8, 33–37. [Google Scholar]
- Nasri, F.; Fadakar, A.; Saba, M.K.; Yousefi, B. Study of indole butyric acid (IBA) effects on cutting rooting improving some of wild genotypes of damask roses (Rosa damascena Mill.). J. Agric. Sci. 2015, 60, 263–275. [Google Scholar] [CrossRef]
- Jabbarzadeh, Z.; Khosh-Khui, M. Factors affecting tissue culture of Damask rose (Rosa damascena Mill.). Sci. Hortic. 2005, 105, 475–482. [Google Scholar] [CrossRef]
- Paul, I.; Poddar Sarkar, M.; Bhadoria, P.B.S. Floral secondary metabolites in context of biotic and abiotic stress factors. Chemoecology 2021, 32, 49–68. [Google Scholar] [CrossRef]
- Mahajan, M.; Kuiry, R.; Pal, P.K. Understanding the consequence of environmental stress for accumulation of secondary metabolites in medicinal and aromatic plants. J. Appl. Res. Med. Aromat. Plants 2020, 18, 100255. [Google Scholar] [CrossRef]
- Akhavan, H.; Zarezadeh Mehrizi, R. Effects of damask rose (Rosa damascena Mill.) extract on chemical, microbial, and sensory properties of sohan (an Iranian confection) during storage. J. Food Qual. Hazards Control 2016, 3, 97–106. [Google Scholar]
- Hamedi, B.; Pirbalouti, A.G.; Rajabzadeh, F. Manures, vermicompost, and chemical fertilizer impacts on the yield and volatile compounds of the damask rose (Rosa damascena Mill.) flower petals. Ind. Crops Prod. 2022, 187, 115470. [Google Scholar] [CrossRef]
- Karami, A.; Khosh-Khui, M.; Salehi, H.; Saharkhiz, M.J. Correlation between anthocyanin and essential oil content of Damask rose (Rosa damascena Mill.). J. Med. Plants By-Prod. 2012, 1, 3–6. [Google Scholar]
- Karami, A.; Jandoust, S. Comparison scent compound emitted from flowers of damask rose and Persian musk rose. Med. Aromat. Plants 2016, 5, 1000259. [Google Scholar] [CrossRef]
- Kumar, R.; Sharma, S.; Sood, S.; Agnihotri, V.K.; Singh, V.; Singh, B. Evaluation of several Rosa damascena varieties and Rosa bourboniana accession for essential oil content and composition in western Himalayas. J. Essent. Oil Res. 2014, 26, 147–152. [Google Scholar] [CrossRef]
- Anonymous. The Good Scents Company Information System. Available online: http://www.thegoodscentscompany.com/index.html (accessed on 1 February 2023).
- Ferreira, V.; Ortín, N.; Escudero, A.; López, R.; Cacho, J. Chemical characterization of the aroma of Grenache rose wines: Aroma extract dilution analysis, quantitative determination, and sensory reconstitution studies. J. Agric. Food Chem. 2002, 50, 4048–4054. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.-Y.; Xue, J.; Cai, X.-D.; Guo, J.; Li, B.; Wu, S. Assessment of the key aroma compounds in rose-based products. J. Food Drug Anal. 2016, 24, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Chalova, V.I.; Manolov, I.G.; Manolova, V.S. Challenges for commercial organic production of oil-bearing rose in Bulgaria. Biol. Agric. Hortic. 2017, 33, 183–194. [Google Scholar] [CrossRef]
- Yaghoobi, M.; Farimani, M.M.; Sadeghi, Z.; Asghari, S.; Rezadoost, H. Chemical analysis of Iranian Rosa damascena essential oil, concrete, and absolute oil under different bio-climatic conditions. Ind. Crops Prod. 2022, 187, 115266. [Google Scholar] [CrossRef]
- Sell, C. Chemistry of Essential Oils. In Handbook of Essential Oils; CRC Press: Boca Raton, FL, USA, 2020; pp. 161–189. [Google Scholar]
- Butnariu, M. Plants as source of essential oils and perfumery applications. In Bioprospecting of Plant Biodiversity for Industrial Molecules; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2021; pp. 261–292. [Google Scholar]
- Sampaio, B.L.; Costa, F.B.D. Influence of abiotic environmental factors on the main constituents of the volatile oils of Tithonia diversifolia. Rev. Bras. Farmacogn. 2018, 28, 135–144. [Google Scholar] [CrossRef]
- Feng, X.; Zhang, W.; Wu, W.; Bai, R.; Kuang, S.; Shi, B.; Li, D. Chemical composition and diversity of the essential oils of Juniperus rigida along the elevations in Helan and Changbai Mountains and correlation with the soil characteristics. Ind. Crops Prod. 2021, 159, 113032. [Google Scholar] [CrossRef]
- Verdeguer, M.; Sánchez-Moreiras, A.M.; Araniti, F. Phytotoxic effects and mechanism of action of essential oils and terpenoids. Plants 2020, 9, 1571. [Google Scholar] [CrossRef]
- Vaičiulytė, V.; Ložienė, K.; Taraškevičius, R.; Butkienė, R. Variation of essential oil composition of Thymus pulegioides in relation to soil chemistry. Ind. Crops Prod. 2017, 95, 422–433. [Google Scholar] [CrossRef]
- Najem, W.; El Beyrouthy, M.; Wakim, L.H.; Neema, C.; Ouaini, N. Essential oil composition of Rosa damascena Mill. from different localities in Lebanon. Acta Bot. Gall. 2011, 158, 365–373. [Google Scholar] [CrossRef]
- Lima, G.P.; Vianello, F. Review on the main differences between organic and conventional plant-based foods. Int. J. Food Sci. Technol. 2011, 46, 1–13. [Google Scholar] [CrossRef]
- Erdal, İ.; Munduz, H. Comparing the Nutritional Status of Organic and Conventional Oil-Bearing Rose (Rosa damascena Mill.) Gardens in Lakes Region With Leaf and Flower Analyzes. Toprak Su Derg. 2017, 6, 26–31. [Google Scholar] [CrossRef]
- Todorova, M.; Dobreva, A.; Petkova, N.; Grozeva, N.; Gerdzhikova, M.; Veleva, P. Organic vs conventional farming of oil-bearing rose: Effect on essential oil and antioxidant activity. BioRisk 2022, 17, 271–285. [Google Scholar] [CrossRef]
- Lamalakshmi Devi, E.; Kumar, S.; Basanta Singh, T.; Sharma, S.K.; Beemrote, A.; Devi, C.P.; Chongtham, S.; Singh, C.H.; Yumlembam, R.A.; Haribhushan, A. Adaptation strategies and defence mechanisms of plants during environmental stress. In Medicinal Plants and Environmental Challenges; Springer: Cham, Switzerland, 2017; pp. 359–413. [Google Scholar]
- O’Brien, A.M. The Confluence of Abiotic and Biotic Forces: Local Adaptation of Teosinte in the Context of Rhizosphere Biota and Environmental Gradients; University of California: Davis, CA, USA, 2017. [Google Scholar]
- Sulmon, C.; Van Baaren, J.; Cabello-Hurtado, F.; Gouesbet, G.; Hennion, F.; Mony, C.; Renault, D.; Bormans, M.; El Amrani, A.; Wiegand, C. Abiotic stressors and stress responses: What commonalities appear between species across biological organization levels? Environ. Pollut. 2015, 202, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A. Plant Abiotic Stress Challenges from the Changing Environment; Frontiers Media SA: Lausanne, Switzerland, 2016; Volume 7, p. 1123. [Google Scholar]
- Wheeler, T.; Von Braun, J. Climate change impacts on global food security. Science 2013, 341, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral volatiles: From biosynthesis to function. Plant Cell Environ. 2014, 37, 1936–1949. [Google Scholar] [CrossRef]
- Wiese, N.; Fischer, J.; Heidler, J.; Lewkowski, O.; Degenhardt, J.; Erler, S. The terpenes of leaves, pollen, and nectar of thyme (Thymus vulgaris) inhibit growth of bee disease-associated microbes. Sci. Rep. 2018, 8, 14634. [Google Scholar] [CrossRef]
- Farré-Armengol, G.; Filella, I.; Llusià, J.; Peñuelas, J. β-Ocimene, a key floral and foliar volatile involved in multiple interactions between plants and other organisms. Molecules 2017, 22, 1148. [Google Scholar] [CrossRef]
- Knauer, A.C.; Schiestl, F.P. The effect of pollinators and herbivores on selection for floral signals: A case study in Brassica rapa. Evol. Ecol. 2017, 31, 285–304. [Google Scholar] [CrossRef]
- Huang, M.; Sanchez-Moreiras, A.M.; Abel, C.; Sohrabi, R.; Lee, S.; Gershenzon, J.; Tholl, D. The major volatile organic compound emitted from Arabidopsis thaliana flowers, the sesquiterpene (E)-β-caryophyllene, is a defense against a bacterial pathogen. New Phytol. 2012, 193, 997–1008. [Google Scholar] [CrossRef]
- Balilashaki, K.; Vahedi, M.; Zaid, A.; Wani, S.H. Unraveling Omics Based Technologies in Enhancing Abiotic Stress in Genus Rosa: Progress and Prospects. J. Pl. Sci. Res. 2019, 35, 25–38. [Google Scholar] [CrossRef]
- Samynathan, R.; Shanmugam, K.; Nagarajan, C.; Murugasamy, H.; Ilango, R.V.J.; Shanmugam, A.; Venkidasamy, B.; Thiruvengadam, M. The effect of abiotic and biotic stresses on the production of bioactive compounds in tea (Camellia sinensis (L.) O. Kuntze). Plant Gene 2021, 27, 100316. [Google Scholar] [CrossRef]
- Hessini, K.; Wasli, H.; Al-Yasi, H.M.; Ali, E.F.; Issa, A.A.; Hassan, F.A.; Siddique, K.H. Graded moisture deficit effect on secondary metabolites, antioxidant, and inhibitory enzyme activities in leaf extracts of Rosa damascena Mill. var. trigentipetala. Horticulturae 2022, 8, 177. [Google Scholar] [CrossRef]
- Abdelmajeed, N.A.; Danial, E.N.; Ayad, H.S. The effect of environmental stress on qualitative and quantitative essential oil of aromatic and medicinal plants. Arch. Des Sci. 2013, 66, 100–120. [Google Scholar]
- Sharifi Ashourabadi, E.; Tabaei-Aghdaei, S.R.; Mirza, M.; Nadery, M.; Nadery, B. Effect of water deficit stress on yield and essential oil components of Rosa damascena Mill. J. Med. Plants By-Prod 2023. [Google Scholar]
- Hassan, F.; Ali, E.; Alamer, K. Exogenous application of polyamines alleviates water stress-induced oxidative stress of Rosa damascena Miller var. trigintipetala Dieck. South Afr. J. Bot. 2018, 116, 96–102. [Google Scholar] [CrossRef]
- Farahani, H.; Sajedi, N.A.; Madani, H.; Changizi, M.; Naeini, M.R. Effect of foliar-applied silicon on flower yield and essential oil composition of damask rose (Rosa damascena Miller) under water deficit stress. Silicon 2021, 13, 4463–4472. [Google Scholar] [CrossRef]
- Kiymaz, S.; Altun, B.; Ertek, A. Effect of different water regimes and nitrogen applications on the growth, yield, essential oil content, and quality parameters of the oil rose (Rosa damascena Mill.). J. Plant Nutr. 2022, 45, 2108–2122. [Google Scholar] [CrossRef]
- Yousefi, B. Screening of Rosa damascena Mill. landraces for flower yield and essential oil content in cold climates. Folia Hortic. 2016, 28, 31–40. [Google Scholar] [CrossRef]
- Saxena, B.; Shukla, K.; Giri, B. Arbuscular mycorrhizal fungi and tolerance of salt stress in plants. In Arbuscular Mycorrhizas and Stress Tolerance of Plants; Springer: Singapore, 2017; pp. 67–97. [Google Scholar]
- Nawaz, K.; Hussain, K.; Majeed, A.; Khan, F.; Afghan, S.; Ali, K. Fatality of salt stress to plants: Morphological, physiological and biochemical aspects. Afr. J. Biotechnol. 2010, 9, 5475–5480. [Google Scholar]
- El Ghali, T.; Marah, H.; Qurtobi, M.; Raibi, F.; Bellarbi, M.; Amenzou, N.; El Mansouri, B. Geochemical and isotopic characterization of groundwater and identification of hydrogeochemical processes in the Berrechid aquifer of central Morocco. Carbonates Evaporites 2020, 35, 37. [Google Scholar] [CrossRef]
- Shavrukov, Y. Salt stress or salt shock: Which genes are we studying? J. Exp. Bot. 2013, 64, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Modi, P.; Dave, A.; Vijapura, A.; Patel, D.; Patel, M. Effect of abiotic stress on crops. In Sustainable Crop Production; IntechOpen: London, UK, 2020; p. 3. [Google Scholar]
- Kant, K.; Gupta, S.; Kaur, N.; Jindal, P.; Ali, A. Novel foliar approaches enhancing active constituents, flower yield and essential oil content in Damask rose (Rosa damascena Mill.): A review. J. Plant Nutr. 2023, 46, 4532–4558. [Google Scholar] [CrossRef]
- Jiang, C.; Xu, J.; Zhang, H.; Zhang, X.; Shi, J.; Li, M.; Ming, F. A cytosolic class I small heat shock protein, RcHSP17. 8, of Rosa chinensis confers resistance to a variety of stresses to Escherichia coli, yeast and Arabidopsis thaliana. Plant Cell Environ. 2009, 32, 1046–1059. [Google Scholar] [CrossRef] [PubMed]
- Cha-Um, S.; Kirdmanee, C. In Vitro Flowering of Miniature Roses (Rosa × hybrida L.’Red Imp’) in Response to Salt Stress. Eur. J. Hortic. Sci. 2010, 75, 239. [Google Scholar]
- Cai, X.; Niu, G.; Starman, T.; Hall, C. Response of six garden roses (Rosa × hybrida L.) to salt stress. Sci. Hortic. 2014, 168, 27–32. [Google Scholar] [CrossRef]
- Li, X.; Wan, S.; Kang, Y.; Chen, X.; Chu, L. Chinese rose (Rosa chinensis) growth and ion accumulation under irrigation with waters of different salt contents. Agric. Water Manag. 2016, 163, 180–189. [Google Scholar] [CrossRef]
- Li, X.; Kang, Y.; Wan, S.; Chen, X.; Xu, J. Effect of drip-irrigation with saline water on Chinese rose (Rosa chinensis) during reclamation of very heavy coastal saline soil in a field trial. Sci. Hortic. 2015, 186, 163–171. [Google Scholar] [CrossRef]
- El-Sharnouby, M.E.; Montaser, M.M.; Abdallah, S.M. Oil and flower production in Rosa damascena trigintipetala Dieck under salinity stress in Taif region, Saudi Arabia. Sustainability 2021, 13, 4547. [Google Scholar] [CrossRef]
- Hassan, F.; Al-Yasi, H.; Ali, E.; Alamer, K.; Hessini, K.; Attia, H.; El-Shazly, S. Mitigation of salt-stress effects by moringa leaf extract or salicylic acid through motivating antioxidant machinery in damask rose. Can. J. Plant Sci. 2020, 101, 157–165. [Google Scholar] [CrossRef]
- Omidi, M.; Khandan-Mirkohi, A.; Kafi, M.; Zamani, Z.; Ajdanian, L.; Babaei, M. Biochemical and molecular responses of Rosa damascena mill. cv. Kashan to salicylic acid under salinity stress. BMC Plant Biol. 2022, 22, 373. [Google Scholar] [CrossRef]
- Attia, H.; Al-Yasi, H.; Alamer, K.; Ali, E.; Hassan, F.; Elshazly, S.; Hessini, K. Induced anti-oxidation efficiency and others by salt stress in Rosa damascena Miller. Sci. Hortic. 2020, 274, 109681. [Google Scholar] [CrossRef]
- Kumar, R.; Sharma, S.; Sood, S.; Kaundal, M.; Agnihotri, V.K. Effect of manures and inorganic fertilizers on growth, yield, and essential oil of damask rose (Rosa damascena Mill.) and chemical properties of soil in western Himalayas. J. Plant Nutr. 2017, 40, 1604–1615. [Google Scholar] [CrossRef]
- Secco, D.; Whelan, J.; Rouached, H.; Lister, R. Nutrient stress-induced chromatin changes in plants. Curr. Opin. Plant Biol. 2017, 39, 1–7. [Google Scholar] [CrossRef]
- Ali, E.F.; Hassan, F.; Abdel-Rahman, S.S.; Siddique, K.H. Foliar application of potassium and zinc enhances the productivity and volatile oil content of damask rose (Rosa damascena miller var. trigintipetala dieck). Acta Sci. Pol. Hortorum Cultus 2021, 20, 101–114. [Google Scholar] [CrossRef]
- Hellemans, T.; Landschoot, S.; Dewitte, K.; Van Bockstaele, F.; Vermeir, P.; Eeckhout, M.; Haesaert, G. Impact of crop husbandry practices and environmental conditions on wheat composition and quality: A review. J. Agric. Food Chem. 2018, 66, 2491–2509. [Google Scholar] [CrossRef]
- Juric, S.; Vlahovicek-Kahlina, K.; Duralija, B.; Bandic, L.; Nekic, P.; Vincekovic, M. Stimulation of plant secondary metabolites synthesis in soilless cultivated strawberries (Fragaria × ananassa Duchesne) using zinc-alginate microparticles. Turk. J. Agric. For. 2021, 45, 324–334. [Google Scholar] [CrossRef]
- Ntobela, B. The Effect of Light Intensity on the Growth and Anti-Pest Activities of Leek (Allium porrum) against the Grapevine Mealybug (Planococcus ficus) and a Phytopathogenic Fungus (Fusarium oxysporum); Cape Peninsula University of Technology: Cape Town, South Africa, 2020. [Google Scholar]
- Kumar, R.; Sharma, S.; Kaundal, M.; Sharma, S.; Thakur, M. Response of damask rose (Rosa damascena Mill.) to foliar application of magnesium (Mg), copper (Cu) and zinc (Zn) sulphate under western Himalayas. Ind. Crops Prod. 2016, 83, 596–602. [Google Scholar] [CrossRef]
- Brady, N.C.; Weil, R.R.; Weil, R.R. The Nature and Properties of Soils; Prentice Hall: Upper Saddle River, NJ, USA, 2008; Volume 13. [Google Scholar]
- Shohayeb, M.; Arida, H.; Abdel-Hameed, E.-S.S.; Bazaid, S. Effects of Macro-and Microelements in Soil of Rose Farms in Taif on Essential Oil Production by Rosa damascena Mill. J. Chem. 2015, 2015, 935235. [Google Scholar] [CrossRef]
- Pal, P.K.; Mahajan, M.; Agnihotri, V.K. Foliar application of plant nutrients and kinetin modifies growth and essential oil profile in Rosa damascena under acidic conditions. Acta Physiol. Plant. 2016, 38, 176. [Google Scholar] [CrossRef]
- Ali, Q.; Shahid, S.; Ali, S.; Javed, M.T.; Iqbal, N.; Habib, N.; Hussain, S.M.; Shahid, S.A.; Noreen, Z.; Hussain, A.I. Trehalose Metabolism in Plants under Abiotic Stresses. In Approaches for Enhancing Abiotic Stress Tolerance in Plants; CRC Press: Boca Raton, FL, USA, 2019; pp. 349–364. [Google Scholar]
- Jia, X.; Feng, H.; Bu, Y.; Ji, N.; Lyu, Y.; Zhao, S. Comparative transcriptome and weighted gene co-expression network analysis identify key transcription factors of Rosa chinensis ‘Old Blush’after exposure to a gradual drought stress followed by recovery. Front. Genet. 2021, 12, 690264. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Chakradhar, D. Analysis and optimization of WEDM performance characteristics of Inconel 706 for aerospace application. Silicon 2018, 10, 921–930. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Odour Type | RI | % Peak Area | References | |
|---|---|---|---|---|---|
| Damask Rose | Musk Rose | ||||
| Monoterpene hydrocarbons | |||||
| α-Pinene | herbal, green | 932 | 14.153 ±1.028 | 0.563 ± 0.155 | [29,52,53] |
| β-Pinene | herbal, green | 974 | 0.916 ± 0.710 | 0.036 ± 0.062 | [29,52,53] |
| β-Myrcene | herbal, green | 988 | 0.833 ± 0.434 | - | [29,52,53] |
| α-Terpinene | herbal, green | 1014 | 0.113 ± 0.070 | - | [29,53] |
| γ-terpinene | herbal, green | 1055 | 0.223 ± 0.152 | 0.106 ± 0.184 | [29,53] |
| Limonene | herbal, green | 1027 | 0.190 ± 0.113 | 0.123 ± 0.131 | [29,50,52,53] |
| Terpinolene | herbal, green | 1086 | 0.060 ± 0.000 | - | [29,53] |
| Oxygenated monoterpenes | |||||
| Hexanol | fruity, floral | 861 | 0.050 ± 0.000 | - | [29,53] |
| Linalool | fruity, floral | 1097 | 0.090 ± 0.014 | - | [29,50,52,53] |
| dihydro-β-Ionone | woody | 1435 | - | 0.181 ± 0.314 | [29,53,54] |
| (E)-β-Ionone | floral | 1482 | - | 1.431 ± 0.252 | [29,53,54] |
| Terpinen-4-ol | woody, earthy | 1174 | 0.070 ± 0.000 | - | [53,54] |
| β-Citronellol | floral, rose | 1226 | 35.530 ± 1.821 | - | [29,50,52,53,54] |
| Neral | sweet, citrus | 1238 | 0.615 ± 0.304 | - | [52,53,54] |
| Eugenol | spicy | 1353 | - | 1.151 ± 0.088 | [52,53,54,55] |
| Geranial | fruity, floral | 1268 | 0.345 ± 0.403 | - | [29,52,53] |
| 2-Phenyl ethyl acetate | floral | 1254 | - | 0.377 ± 0.342 | [53,54] |
| 2-Phenyl propyl butanoate | fruity | 1484 | - | 0.105 ± 0.182 | [53,54] |
| Geranyl acetate | fruity, floral | 1317 | 4.906 ± 0.833 | - | [29,53] |
| Geranyl propanoate | floral | 1496 | 0.640 ± 0.000 | - | [53,54] |
| Sesquiterpenes hydrocarbons | |||||
| Sabinene | woody | 970 | 0.340 ± 0.155 | - | [53,54] |
| (E)-β-Farnesene | woody | 1459 | 0.640 ± 0.000 | - | [53,54] |
| α-Selinene | n/d | 1498 | 1.580 ± 0.675 | - | [53,54] |
| Hexadecen-1-ol | n/d | 1866 | 0.180 ± 0.000 | - | [52,53,54] |
| 1-Tricosene | n/d | 2285 | - | 1.972 ± 0.416 | [53,54] |
| Oxygenated sesquiterpenes | |||||
| trans-Rose oxide | floral | 1125 | 0.200 ± 0.096 | - | [52,53,56] |
| Benzaldehyde | fruity | 957 | 0.146 ± 0.081 | - | [53,54] |
| p-Cymene | terpenic | 1022 | - | 0.060 ± 0.104 | [53,54] |
| Benzyl alcohol | floral | 1029 | 0.186 ± 0.075 | - | [53,54] |
| Benzene acetaldehyde | green | 1041 | - | 0.103 ± 0.091 | [53,54] |
| 2-Phenylethyl alcohol | floral, rose | 1110 | 36.600 ± 2.052 | 54.152 ± 1.340 | [52,53,55] |
| Methyl eugenol | spicy | 1402 | 0.185 ± 0.077 | - | [52,53,54] |
| n-Octadecanol | n/d | 2072 | - | 0.491 ± 0.450 | [52,53] |
| Aliphatic hydrocarbons | |||||
| Hexacosane | n/d | 2554 | 0.210 ± 0.000 | - | [53] |
| 1-Nonadecene | n/d | 1874 | - | 15.576 ± 1.708 | [29,50,53] |
| n-Nonadecane | n/d | 1900 | 2.350 ± 0.385 | 8.147 ± 0.143 | [29,50,52,53] |
| Heneicosane | n/d | 2105 | 0.210 ± 0.000 | 8.175 ± 0.801 | [50,52,53] |
| Tricosane | n/d | 2285 | - | 1.196 ± 0.071 | [50,53] |
| n-Tetradecane | n/d | 1401 | 0.290 ± 0.000 | - | [53] |
| n-Pentadecane | n/d | 1499 | - | 0.140 ± 0.242 | [50,52,53] |
| 1-Heptadecene | n/d | 1669 | - | 1.261 ± 0.152 | [50,52,53] |
| n-Heptadecane | n/d | 1698 | 0.646 ± 0.61 | 1.711 ± 0.067 | [50,52,53] |
| Total | 97.057 ± 0.347 | 99.556 ± 0.561 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charoimek, N.; Phusuwan, S.; Petcharak, C.; Huanhong, K.; Prasad, S.K.; Junmahasathien, T.; Khemacheewakul, J.; Sommano, S.R.; Sunanta, P. Do Abiotic Stresses Affect the Aroma of Damask Roses? Plants 2023, 12, 3428. https://doi.org/10.3390/plants12193428
Charoimek N, Phusuwan S, Petcharak C, Huanhong K, Prasad SK, Junmahasathien T, Khemacheewakul J, Sommano SR, Sunanta P. Do Abiotic Stresses Affect the Aroma of Damask Roses? Plants. 2023; 12(19):3428. https://doi.org/10.3390/plants12193428
Chicago/Turabian StyleCharoimek, Nutthawut, Sirinun Phusuwan, Chaleerak Petcharak, Kiattisak Huanhong, Shashanka K. Prasad, Taepin Junmahasathien, Julaluk Khemacheewakul, Sarana Rose Sommano, and Piyachat Sunanta. 2023. "Do Abiotic Stresses Affect the Aroma of Damask Roses?" Plants 12, no. 19: 3428. https://doi.org/10.3390/plants12193428
APA StyleCharoimek, N., Phusuwan, S., Petcharak, C., Huanhong, K., Prasad, S. K., Junmahasathien, T., Khemacheewakul, J., Sommano, S. R., & Sunanta, P. (2023). Do Abiotic Stresses Affect the Aroma of Damask Roses? Plants, 12(19), 3428. https://doi.org/10.3390/plants12193428