1. Introduction
Reproductive barriers have been observed in the intraspecific and interspecific cross between the AA genome
Oryza species, to which Asian rice (
O. sativa) and African rice (
O. glaberrima) belong. We identified some causal genes of these reproductive barriers, such as hybrid weakness [
1,
2,
3] and hybrid chlorosis [
4] in
Oryza sativa. These barriers are obstacles to the genetic improvement of rice, on which more than half of all people depend for staple food. Reproductive barriers have also been studied in the context of differentiation and speciation [
5,
6,
7]. Detailed studies on reproductive barriers contribute to both rice breeding and evolutionary biology [
8,
9]. Among reproductive barriers, hybrid seed (spikelet) sterility has been intensively studied because it is related to hybrid rice breeding. Asian rice
O. sativa comprises two major subspecific groups,
indica and
japonica. The hybrid between
indica and
japonica shows hybrid vigor, but also shows hybrid seed sterility [
10]. The major gene causing hybrid seed sterility is
S5 [
11]. The hybrid seed sterility caused by the
S5 gene is explained by an egg-killer model: In heterozygotes of
S5-i derived from indica and
S5-j derived from japonica,
S5-i and
S5-j respectively act as killer allele and abortive allele, leading to semi-sterility, with about half of the seeds being sterile. Other genes, such as
S7 and
S9, also follow the same egg-killer model [
12,
13]. The interspecific crosses between
O. sativa and the African rice
O. glaberrima show hybrid vigor but are highly sterile both in pollen and seeds [
14]. Some causes of this hybrid sterility are explained by a gametic eliminator model [
14,
15,
16]: In a heterozygous state of killer allele and abortive allele, both the eggs and pollen carrying the abortive allele are killed in the gametic eliminator model, causing semi-sterility in both spikelet and pollen. In the selfed progeny of the egg-killer model, the ratio of homozygotes of killer allele and heterozygotes are expected to be 1:1, and no homozygotes of the abortive allele are expected to appear. On the other hand, in the selfed progeny of the gametic eliminator model, only homozygotes of the killer allele are expected to appear. Mapping and characterization of the causal genes of hybrid seed and pollen sterility could contribute to hybrid rice breeding.
O. meridionalis is an AA genome
Oryza species from Northern Australia and Indonesia [
17]. Several types of molecular data indicate that this species has diverged from the other AA genome species and that it is distantly related to them [
18,
19]. This divergence is reflected by the low pollen fertility of the hybrid between
O. meridionalis and the other AA genome species [
20,
21,
22]. Li et al. [
23] identified five hybrid pollen sterility genes, all of which follow the pollen-killer model [
15,
16]. No genes conferring hybrid seed sterility have been reported in the cross between
O. meridionalis and the other AA genome species before our recent study [
24], in which we reported seed abortion after fertilization in the progeny from the cross between
O. sativa and
O. meridionalis was controlled by a
SEED DEVELOPMENT 1 (
SDV1) gene and a
SEED DEVELOPMENT 2 (
SDV2) gene. This gene model is not an egg-killer model but a duplication and loss of a functional gene for seed development—a new finding in rice genetics. However, seed sterility is not completely explained by these genes in the cross between an
O. meridionais strain Jpn2 and
O. sativa. In the present study, we report that an egg-killer gene also contributes to the sterility in the above cross combination. Judging from the gene location and gene action, this gene is thought to be a new finding in rice genetics.
3. Discussion
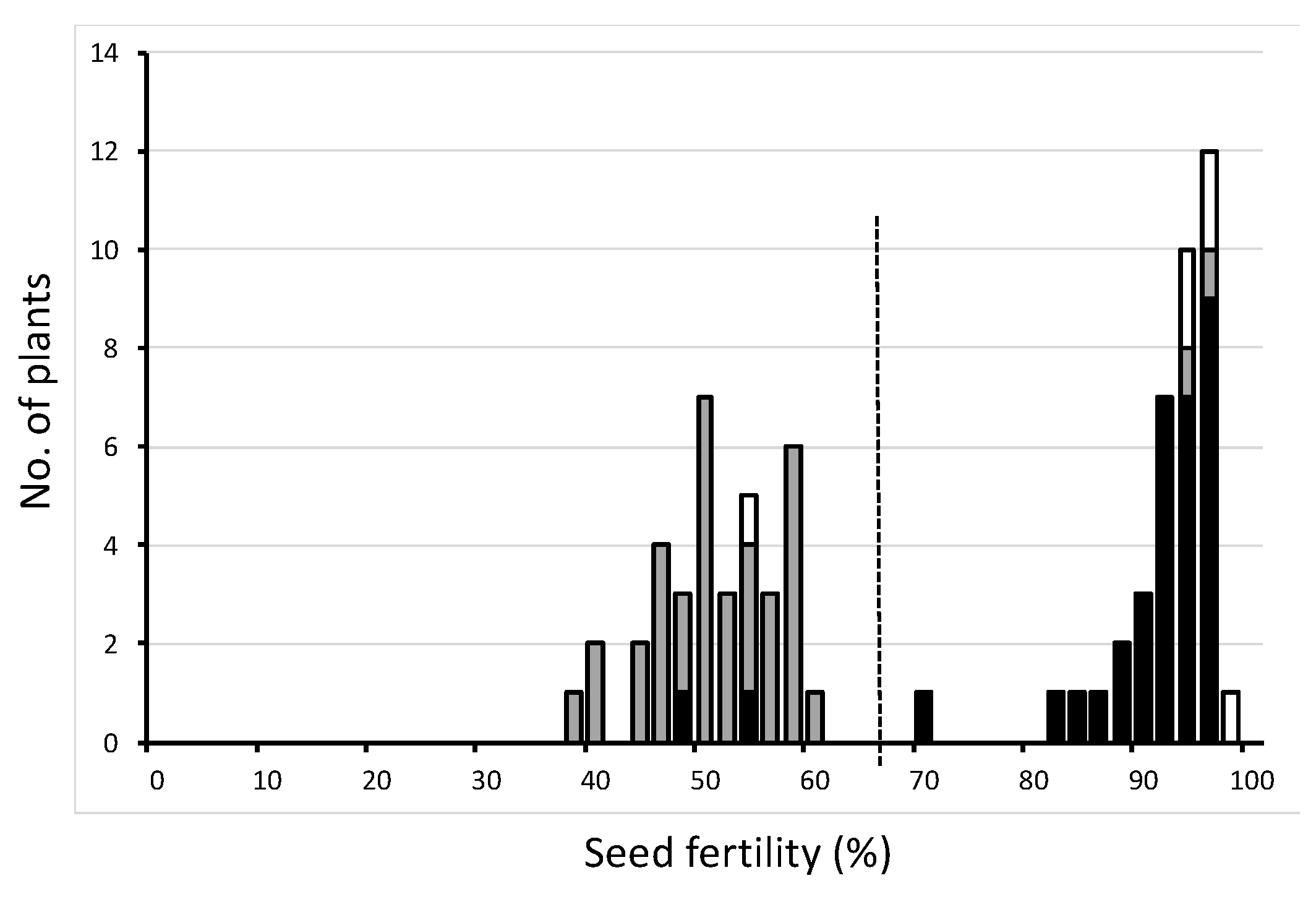
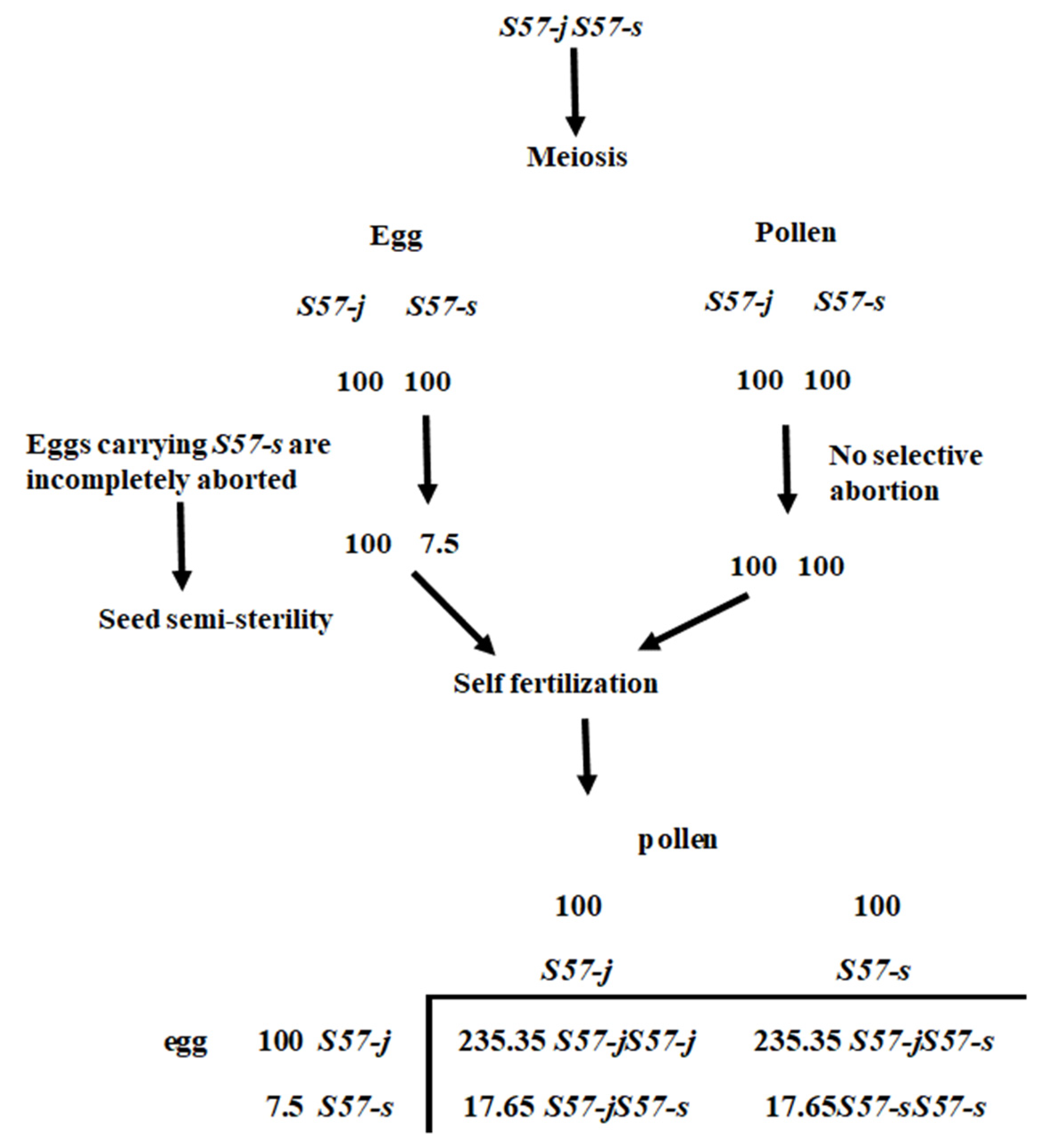
In this study, seed semi-sterility observed in the backcross progeny using an
O. meridionalis strain Jpn2 as donor parent and T65 as a recurrent parent was genetically analyzed. A gene from Jpn2 was the cause of semi-sterility. As shown in
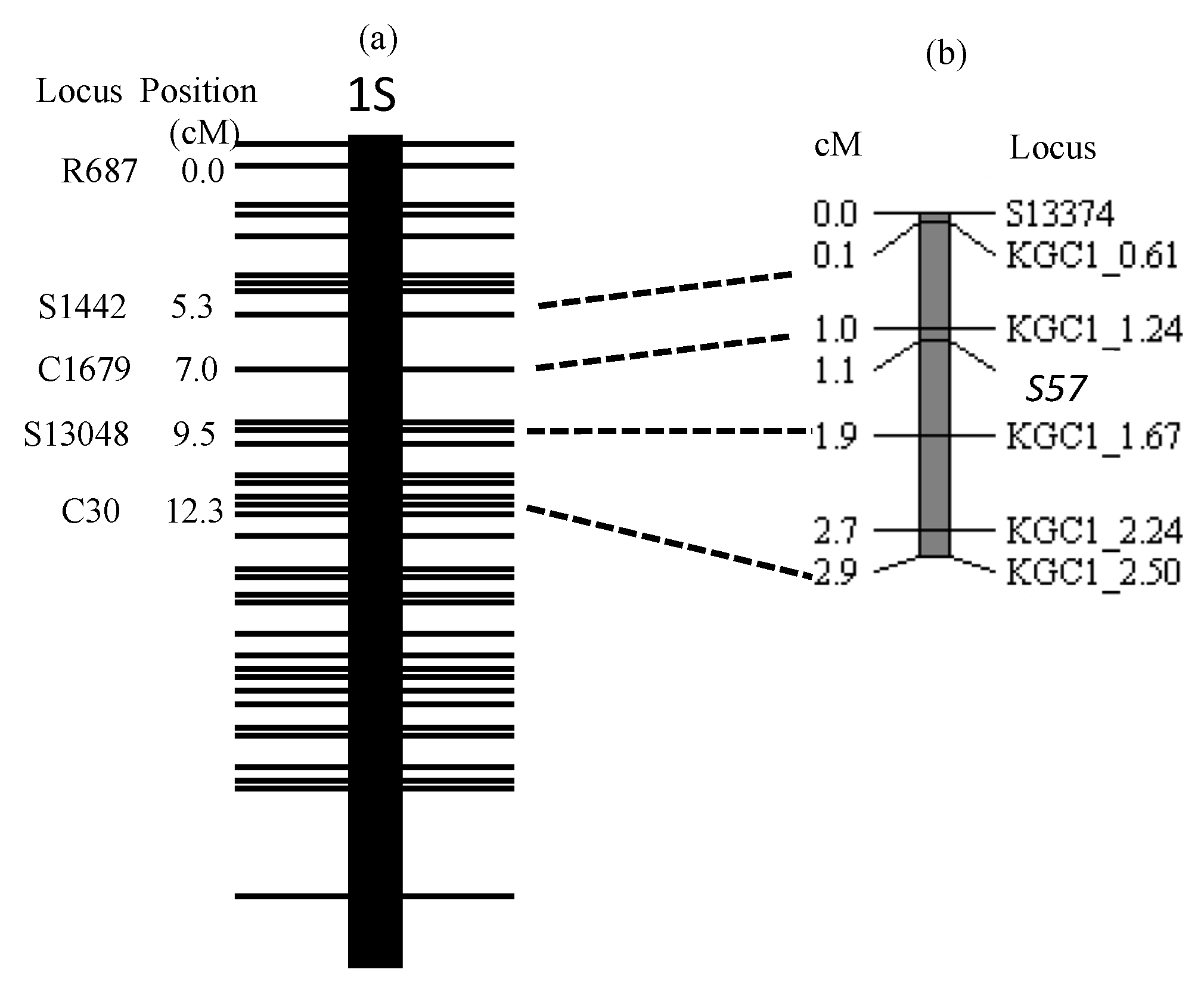
Figure 5, in heterozygous form, eggs carrying the T65 allele were incompletely sterilized. This gene exerts no effect on pollen sterilization. Therefore, this phenomenon can be explained by an egg-killer gene model. Linkage analysis indicated that this gene is located on the distal end of the short arm of chromosome 1 (
Figure 3). Genes with similar functions were observed in intersubspecific crosses in
Oryza sativa, such as
S5 [
11],
S7 [
12] and
S9 [
13]. According to [
27], in the cross between
O. sativa and
O. meridionalis, only pollen-killer genes have been reported. Li et al. [
23] performed linkage analysis of five pollen-killer genes:
S51 located on chromosome 1,
S52 and
S53 located on chromosome 2,
S54 and
S55 on chromosome 7. Yu et al. [
28] also identified the pollen-killer gene
qSHM7 in the cross between
O. sativa and
O. meridionalis and located it on the same chromosomal location as
S55. Furthermore,
O. meridionalis alleles behaved as pollen-killer alleles at the
S51 and
S54 loci and behaved as abortive alleles at the
S52,
S53 and
S55 (
qHMS7) loci. To our knowledge, there have been no reports of egg-killer genes in this chromosomal region, and no report of those found in the cross between
O. sativa and
O. meridionalis. Two egg-killer genes were reported to be located on chromosome 1:
ESA was detected in the cross between
O. sativa and
O. rufipogon. This gene was tightly linked with RM24, located at 18.9 Mb on chromosome 1 of IRGSP 1.0 pseudomolecule [
29].
S40 was detected in the cross between
O. sativa and
O. longistaminata. This gene was tightly linked with RM575, located at 8.1 Mb [
30]. Therefore, the egg-killer gene in this study is a new gene.
In the history of rice gene nomenclature, S with digits have been applied to the gene symbols of both pollen sterility genes and egg sterility genes. According to a database of rice, Oryzabase (Oryzabase.Available online:
https://shigen.nig.ac.jp/rice/oryzabase/locale/change?lang=en accessed on 31 December 2021) [
31], and a recent review on hybrid sterility by Li et al. [
27],
S56(t) is the latest gene symbol, which was found in the cross between
O. sativa and
O. glumaepatula, and caused pollen semi-sterility [
32]. Therefore, we name the gene found in this study as
HYBRID SPIKELET STERILITY 57, with the gene symbol of
S57, following Yoshimura and Nagato [
33] and McCouch and CGSNL [
34]. In the intraspecific crosses among
O. sativa, there have been no reports of hybrid spikelet sterility phenomena caused by a gene located on the distal end of the short arm of chromosome 1. Therefore, all
O.
sativa is thought to share the same allele found in T65. This allele was called
S57-s (s for
sativa). The allele of Jpn2 was not shared by another
O. meridionalis strain, W1297. This allele was called
S57-j (j for Jpn2).
S57 is located in the 430.6 kb chromosomal region encompassed by the two DNA markers KGC1_1.24 and KGC1_.67. We are undertaking fine-mapping of the S57 gene to identify the causal DNA sequence diversifying the function of the S57 gene. According to Rice Genome Annotation Project (Rice Genome Annotation Project. Available online:
http://rice.uga.edu/index.shtml accessed on 15 January 2022) [
35], 69 genes with the gene name LOC_Os01g0**** are located on that region in
O. sativa cultivar Nipponbare genome IRGSP 1.0 pseudomolecule [
35]. We are undertaking fine-mapping of the S57 gene to identify the causal DNA sequence diversifying the function of the S57 gene. In the case of the
S1 gene, the corresponding DNA sequence of killer allele in
O. glaberrima is not present only in
O. sativa genome [
15]. After delimiting the candidate chromosomal region, we will search for the candidate gene in both
O. sativa and
O. meridionalis genomes. The project of sequencing the Jpn2 genome is underway to uncover
S57,
SDV1,
SDV2 and long grain gene(s) from Jpn2 [
36].
In a review by Calvo-Baltanás et al. [
37] on hybrid incompatibility, genetic and molecular studies of hybrid incompatibility in numerous plant species revealed that such self-destructing symptoms in most cases are attributed to autoimmunity: plant immune responses are inadvertently activated in the absence of pathogenic invasion. Most of the hybrid incompatibility, such as hybrid weakness and hybrid necrosis, is explained by a conflict involving a member of the major plant immune receptor family, the nucleotide-binding domain and leucine-rich repeat-containing protein (NLR; formerly known as NBS-LRR). NLR genes are associated with disease resistance traits. According to [
37], hybrid sterility can be explained in the same context. The
S5 gene encodes a disease resistance-related aspartic protease (AP) [
11]. Although the AP family has not yet been investigated in line with NLR activity, members of this family are involved in defense responses with activated salicylic acid in addition to pollen and ovule development in multiple plant species. Furthermore, the
S5 gene is actually a gene complex of three open reading frames, ORF3, ORF4 and ORF5 [
11]. ORF5 encodes AP. AP from
indica S5 allele behaves as a killer. Because products from
indica ORF3 behave as the protector from killer AP, eggs carrying
indica S5 alleles are viable. A pollen-killer gene
qHMS7 is actually composed of two tightly linked genes, ORF2 and ORF3 [
28]. ORF2 encodes a ribosome-inactivating protein (RIP) domain-containing protein, which behaves as a toxic genetic element that aborts pollen in a sporophytic manner, whereas ORF3 is predicted to encode homologous, grass family-specific proteins with a mitochondrial targeting signal at the N terminus, which behaves as an antidote that protects pollen in a gametophytic manner. RIPs are toxic RNA N-glycosidases that affect translation processes and have been implicated in apoptotic pathways in mammalian cells and antiviral, antifungal and insecticidal activities in plants [
38]. The above two abortion models involve proteins related to plant immunity. Therefore, the S57 egg-killer system might evolve from genes conferring plant immunity and be composed of tightly linked plural genes. Fine mapping of this gene will test the universality of killer-protector systems reported earlier.
According to [
27], the
S53 gene [
23], a pollen-killer gene found in the cross between
O. sativa and
O. meridiolis, is located on the same location as other pollen-killer genes,
S22A,
S22B [
39] and
S29 [
40] on the short arm of chromosome 2. In the cross between
O. sativa and
O. meridiolis,
O. sativa allele behaves as pollen-killer and
O. meridionalis allele as abortive allele at
S53 locus. In the cross between
O. sativa and
O. glumaepatula, the
O. sativa allele behaves as pollen-killer and
O. glumaepatula allele as abortive allele at S22A and S22B loci. In the cross between
O. sativa and
O. glaberrima, the
O. glaberrima allele behaves as pollen-killer and
O. sativa allele as abortive allele at
S29 locus. If
S53,
S29 and
S22A or
S22B are located on the same locus, the hierarchy of tentative allelic interaction would be that
O. glaberrima carries the strongest egg-killer allele, followed by
O. sativa, and
O. meridionalis and
O. glumaepatula carry abortive alleles. These kinds of allelic hierarchy could be detected by testcross: The progeny of homozygotes of
S57-j of BC
5F
6 will be the tester lines for the allele of
S57. Introgression lines of AA genome wild rice chromosomal segments under T65 genetic background were developed [
41,
42]. The testcross with homozygotes of
S57-j and intercrosses among introgression lines carrying the introgressed chromosomal segments of the distal end of chromosome 1 would clarify the hierarchy of alleles at the
S57 locus.
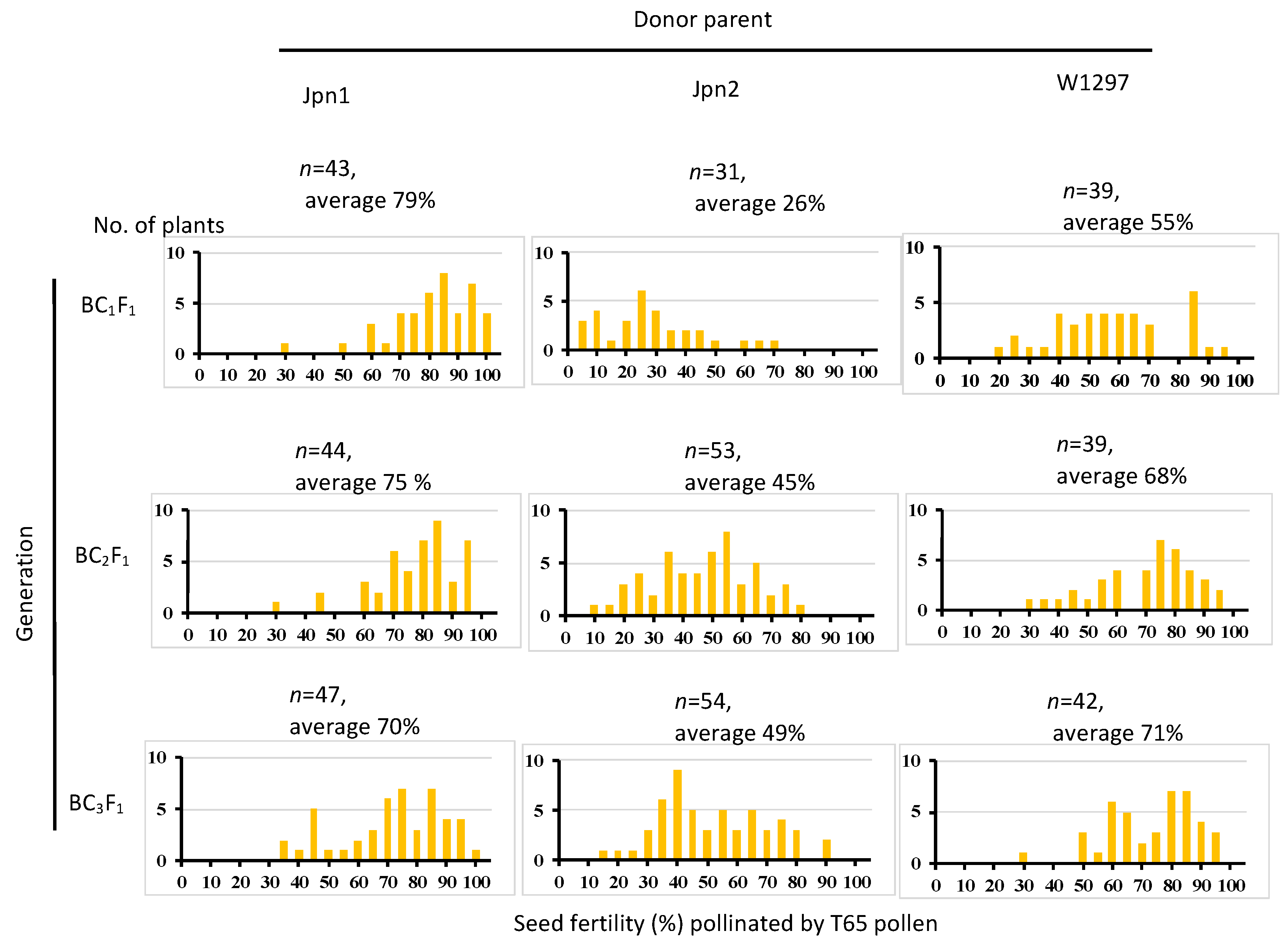
Lam et al. [
22] reported that the hybrid between Jpn2 and W1297, that between Jpn2 and another
O. meridionalis strain W1299, showed very low seed fertility and that the hybrid between W1297 and W1299 showed seed fertility comparable to parental lines. It suggests that W1299 also does not carry the egg-killer allele at
S57. Additionally, W1297, W1299 and Jpn2 have originated from different places in Australia: W1297and W1299 are from the Northern Territory, and Jpn2 is from Queensland. According to Juliano et al. [
43], most crosses between Northern Territory and Queensland accessions produced sterile hybrids, and DNA marker-based analyzes showed
O. meridionalis genetic differentiation corresponding to geographic origin. In
Oryza sativa, the carriers of the egg-killer allele and those of the abortive allele at
S5 corresponded to
indica and
japonica [
8]. The distribution of alleles of
S57 in the wild populations of
O. meridiolnais will test the role of the egg-killer gene in population differentiation in nature.




 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}