
Calcium and Boron Fertilization Improves Soybean Photosynthetic Efficiency and Grain Yield

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Climate Characteristics
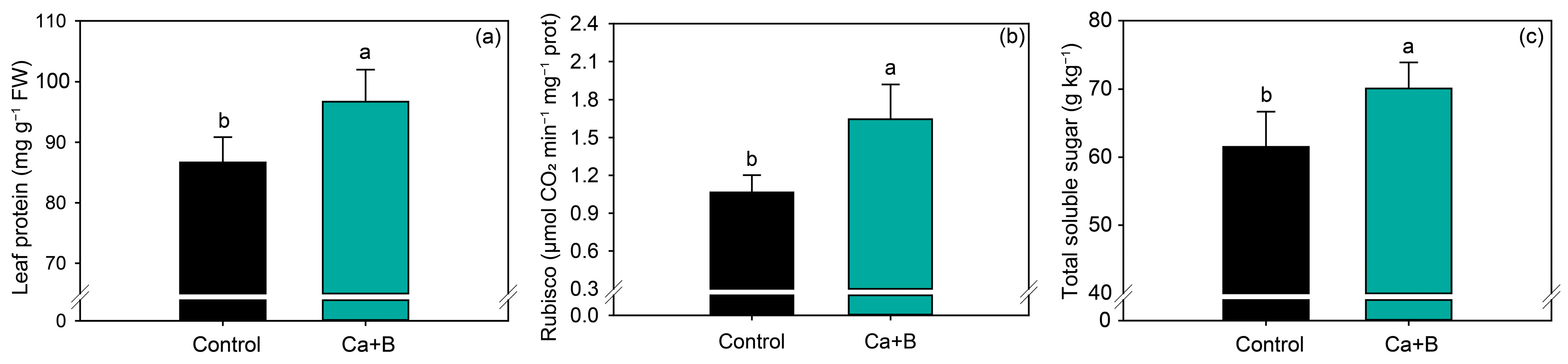
2.2. Nutritional Status, Gas Exchange and Carbon Metabolism
2.3. Yield Components and Grain Yield
3. Discussion
4. Materials and Methods
4.1. Field Description
4.2. Experimental Design and Treatment Establishment
4.3. Soybean Cultivation
4.4. Assessment of Soybean Chemical and Physiological Parameters
4.4.1. Crop Nutritional Status
4.4.2. Gas Exchange Parameters
4.4.3. Total Concentration of Soluble Sugar
4.4.4. Total Leaf Protein Concentration
4.4.5. Photosynthetic Enzyme Activity
4.4.6. Agronomic Parameters and Grain Yield
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borket, C.M.; Farias, J.R.B.; Sfredo, G.J.; Tutida, F.; Spoladori, C.L. Respostas da soja à adubação e disponibilidade de potássio em Latossolo roxo álico. Pesqui. Agropecuária Bras. 1997, 32, 1119–1129. [Google Scholar]
- Fenández, V.; Eichert, E.T. Uptake of hydrophilic solutes through plant leaves: Current state of Knowledge and Perspectives of foliar fertilization. Plant Sci. 2009, 28, 38–68. [Google Scholar] [CrossRef]
- Fernández, V.; Brown, P.H. From plant surface to plant metabolism: The uncertain fate of foliar-applied nutrients. Plant Sci. 2013, 4, 1–5. [Google Scholar] [CrossRef]
- Oliveira, S.M.; Pieroza Júnior, C.; Lago, B.C.; Almeida, R.E.M.; Trivelin, P.C.O.; Favarin, J.L. Grain yield, efficiency and the allocation of foliar N applied to soybean canopies. Crop Sci. 2019, 76, 305–310. [Google Scholar] [CrossRef]
- Patzke, K.; Prananingrum, P.; Klemens, P.A.; Trentmann, O.; Rodrigues, C.M.; Keller, I.; Fernie, A.R.; Geigenberger, P.; Bolter, B.; Lehmann, M.; et al. The Plastidic Sugar Transporter pSuT Influences Flowering and Affects Cold Responses. Plant Physiol. 2019, 179, 569–587. [Google Scholar] [CrossRef] [PubMed]
- Zoz, T.; Steiner, F.; Seidel, E.P.; Castagnara, D.D.; Souza, G.E. Foliar application of calcium and boron improves the spike fertily and yield of wheat. Biosci. J. 2016, 32, 873–880. [Google Scholar] [CrossRef]
- Domingos, C.S.; Besen, M.R.; Esper Neto, M.; Costa, E.J.O.; Scapim, C.A.; Inoue, T.T.; Batista, M.A.; Braccini, A.L. Can calcium and boron leaf application increase soybean yield and seed quality. Acta Agric. Scand. 2021, 71, 171–181. [Google Scholar] [CrossRef]
- Brand, J.J.; Becker, D.W. Evidence for direct roles of calcium in photosynthesis. J. Bioenerg. Biomembr. 1983, 16, 239–249. [Google Scholar] [CrossRef]
- Krichevsky, A.; Kozlovsky, S.V.; Tian, G.W.; Chen, M.H.; Zaltsman, A.; Citovsky, V. How pollen tubes grow. Dev. Biol. 2007, 303, 405–420. [Google Scholar] [CrossRef]
- Fioreze, S.L.; Tochetto, C.; Coelho, A.E.; Melo, H.F. Effects of calcium supply on soybean plants. Comun. Sci. 2018, 9, 219–225. [Google Scholar] [CrossRef]
- Du, W.; Pan, Z.Y.; Hussain, S.B.; Han, Z.X.; Peng, S.A.; Liu, Y.Z. Foliar supplied boron can be transported to roots as a boron-sucrose complex via phloem in citrus trees. Front. Plant Sci. 2020, 11, 250. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Ilyas, N.; Arshad, M.; Meraj, T.A.; Hefft, D.I.; Jan, B.L.; Ahmad, P. The impact of calcium, potassium, and boron application on the growth and yield characteristics of Durum Wheat under Drought conditions. Agronomy 2022, 12, 1917. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Thor, C. Calcium: Nutrient and messenger. Front. Plant Sci. 2019, 10, 1–7. [Google Scholar] [CrossRef]
- Poovaiah, B.W.; Leopold, A.C. Inhibition of Abscission by Calcium. Plant Physiol. 1973, 51, 848–851. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sawicki, M.; Aït Barka, E.; Clément, C.; Vaillant-Gaveau, N.; Jacquard, C. Cross-talk between environmental stresses and plant metabolism during reproductive organ abscission. J. Exp. Bot. 2015, 66, 1707–1719. [Google Scholar] [CrossRef]
- Wimmer, M.; Eichert, T. Review: Mechanisms for boron deficiency-mediated changes in plant water relations. Plant Sci. 2013, 203, 25–32. [Google Scholar] [CrossRef]
- Kaur, G.; Nelson, K.A. Effect of foliar boron fertilization of fine textured soils on corn yields. Agronomy 2015, 5, 1–18. [Google Scholar] [CrossRef]
- Al-Mayahi, A.M.W. Effect of Calcium and Boron on Growth and Development of Callus and Shoot Regeneration of Date Palm Barhee. Can. J. Plant Sci. 2020, 100, 357–364. [Google Scholar] [CrossRef]
- Kabir, R.; Yeasmin, S.; Islam, A.K.M.M.; Sarkar, M.A.R. Effect of Phosphorus, Calcium and Boron on the Growth and Yield of Groundnut (Arachis hypogea L.). Int. J. Bio-Sci. Bio-Technol. 2013, 5, 51–60. [Google Scholar]
- Sahin, S.; Gebologlu, N.; Karaman, M.R. Interactive effect of calcium and boron on growth, quality and mineral content of tomato (Solanum Lycopersicon L.). Fresenius Environ. Bull. 2015, 24, 1624–1628. [Google Scholar]
- Singh, R.; Sharma, R.R.; Tyagi, S.K. Pre-harvest foliar application of calcium and boron influences physiological disorders, fruit yield and quality of strawberry (Fragaria × ananassa Duch.). Sci. Hortic. 2007, 112, 215–220. [Google Scholar] [CrossRef]
- Wen-xin, W.; Jia-ju, C.; En-Sheng, Z.; Han-zhang, L.; Qun, L.; Xue-yun, Y. Effects of calcium and boron on the growth, yield and quality of peanut. Subtrop. Plant Sci. 2001, 30, 20–23. [Google Scholar]
- Sadras, V.O. A quantitative top-down view of interactions between stresses: Theory and analysis of nitrogen-water co-limitation in Mediterranean agro-ecosystems. Aust. J. Agric. Res. 2005, 56, 1151–1157. [Google Scholar] [CrossRef]
- Schon, M.K.; Blevins, D.G. Foliar boron application increase the final number of branches and podas on brances of fielf-grown soybeans. Plant Physiol. 1990, 92, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.R.; Slaton, N.A.; Brye, K.R.; DeLong, R.E. Boron fertilization influences soybean yield and leaf and seed boron concentrations. Agron. J. 2006, 98, 198–205. [Google Scholar] [CrossRef]
- Reinbott, T.M.; Blevins, D.G. Response of soybean to foliar-applied boron and magnesium and soil-applied boron. J. Plant Nutr. 1995, 18, 179–200. [Google Scholar] [CrossRef]
- Terry, N.; Huston, R.P. Effects of calcium on the photosynthesis of intact leaves and isolated chloroplasts of sugar beets. Plant Physiol. 1975, 55, 923–927. [Google Scholar] [CrossRef]
- Brdar-Jokanovic, M. Boron toxicity and deficiency in agricultural plants. Int. J. Mol. Sci. 2020, 21, 1424. [Google Scholar] [CrossRef]
- He, L.; Yu, L.; Du, N.; Guo, S. The effect of exogenous calcium on cucumber fruit quality, photosynthesis, chlorophyll fluorescence, and fast chlorophyll fluorescence during the fruiting period under hypoxic stress. BMC Plant Biol. 2018, 18, 1–10. [Google Scholar] [CrossRef]
- Rehman, A.; Farooq, M.; Rashid, A.; Nadeem, F.; Stuerz, S.; Asch, F.; Bell, R.W.; Siddique, K.H.M. Boron nutrition of rice in different production systems: A review. Agron. Para O Desenvolv. Sustent. 2018, 38, 1–24. [Google Scholar]
- Weaver, M.L.; Timm, H.; Nag, H.; Burke, D.W.; Silbernagel, M.J.; Foster, K. Pod retention and seed yield of beans in response to chemical foliar applications. HortScience 1985, 20, 429–431. [Google Scholar] [CrossRef]
- Bevilaqua, G.A.P.; Silva Filho, P.M.; Possenti, J.C. Aplicação foliar de cálcio e boro e componentes de rendimento e qualidade de sementes de soja. Ciência Rural 2002, 32, 31–34. [Google Scholar] [CrossRef]
- Souza, L.C.D.; Sá, M.E.; Carvalho, M.A.C.; Simudi, H.M. Produtividade de quatro cultivares de soja em função da aplicação de fertilizantes mineral foliar a base de cálcio e boro. Rev. Biol. E Ciências Terra 2008, 8, 37–44. [Google Scholar]
- Hepler, P.K. Calcium: A central regulator of plant growth and development. Célula Veg. 2005, 17, 2142–2155. [Google Scholar] [CrossRef]
- van Raij, B.; Cantarella, H.; Quaggio, J.A.; Furlani, A.M.C. Recomendação de Adubação e Calagem para o Estado de São Paulo, 2nd ed.; Instituto Agronômico/Fundação IAC: Campinas, Brazil, 1997; 285p.
- Brown, P.H.; Bellaloui, N.; Wimmer, M.A.; Bassil, E.S.; Ruiz, J.; Hu, H.; Pfeffer, H.; Dannel, F.; Römheld, V. Boron in plant biology. Plant Biol. 2002, 4, 205–223. [Google Scholar] [CrossRef]
- Taylor, S.H.; Orr, D.J.; Silva, E.C.; Long, S.P. During photosynthetic induction, biochemical and stomatal limitations differ between Brassica crops. Plant Cell Environ. 2020, 43, 2623–2636. [Google Scholar] [CrossRef]
- Fernández, V.; Sotiropoulos, T.; Brown, P. Foliar Fertilization:Scientific Principles and Field Practices; International Fertilizes Industry Association: Paris, France, 2015; 144p. [Google Scholar]
- Princi, M.P.; Lupini, A.; Araniti, F.; Longo, C.; Mauceri, A.; Sunseri, F.; Abenavoli, M.R. Boron toxicity and tolerance in Plants: Recent Advances and Future Perspectives. In Plant Metal Interaction; Ahmad, P., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 115–147. [Google Scholar]
- Oikonomou, A.; Ladikou, E.V.; Chatziperou, G.; Margaritopoulou, T.; Landi, M.; Sotiropoulos, T.; Araniti, F.; Papadakis, I.E. Boron excess imbalances root/shoot allometry, photosynthetic and chlorophyll fluorescence parameters and sugar metabolism in apple plants. Agronomy 2019, 9, 731. [Google Scholar] [CrossRef]
- Bender, R.R.; Haegele, J.W.; Below, F.E. Nutrient uptake, partitioning, and remobilization in modern soybean varieties. Agron. J. 2015, 107, 563–573. [Google Scholar] [CrossRef]
- Barth, G.; Franscisco, E.; Tamie Suvama, J. Nutrient Uptake Illustrated for Modern, High-Yielding Soybean. Better Crop Plant Food 2018, 102, 11–14. [Google Scholar] [CrossRef]
- Moreira, A.; Moraes, L.A.C.; Schroth, G.; Becker, F.J.; Mandarino, J.M.G. Soybean yield and nutritional status response to nitrogen sources and rates of foliar fertilization. Agron. J. 2017, 109, 629–635. [Google Scholar] [CrossRef]
- de Souza, L.C.D. Efeito da Aplicação de Fertilizante Mineral via Foliar Sobre a Produção e Qualidade Fisiológica de Sementes de Soja; Dissertação (Mestrado)—Agronomia—Unesp: Ilha Solteira, Brazil, 2007. [Google Scholar]
- Naeem, M.; Naeem, M.H. Foliar-applied calcium induces drought stress tolerance in maize by manipulating osmolyte accumulation and antioxidative responses. Pak. J. Bot. 2017, 49, 427–434. [Google Scholar]
- Song, Q.; Liu, Y.; Pang, J.; Yong, J.W.H.; Chen, Y.; Bai, C.; Gille, C.; Shi, Q.; Wu, D.; Han, X.; et al. Supplementary calcium restores peanut (Arachis hypogaea) growth and photosynthetic capacity under low nocturnal temperature. Front. Plant Sci. 2020, 10, 1637. [Google Scholar] [CrossRef]
- Zahra, N.; Wahid, A.; Shaukat, K.; Rasheed, T. Role of Seed Priming and Foliar Spray of Calcium in Improving Flag Leaf Growth, Grain Filling and Yield Characteristics in Wheat (Triticum aestivum)—A Field Appraisal. Int. J. Agric. Biol. 2020, 24, 1591–1600. [Google Scholar] [CrossRef]
- Calonego, J.C.; Ocani, K.; Ocani, M.; Santos, C.H. Adubação boratada foliar na cultura da soja. Colloq. Agrar. 2020, 6, 20–26. [Google Scholar] [CrossRef]
- Tcherkez, G.; Limami, A.M. Net Photosynthetic CO2 Assimilation: More than Just CO2 and O2 Reduction Cycles. New Phytol. 2019, 223, 520–529. [Google Scholar] [CrossRef]
- Oliveira, S.L.; Crusciol, C.A.C.; Rodrigues, V.A.; Galeriani, T.M.; Portugal, J.R.; Bossolani, J.W.; Moretti, L.C.; Calonego, J.C.; Cantarella, H. Molybdenum foliar fertilization improves photosynthetic metabolism and grain yields of field-grown soybean and maize. Front. Plant Sci. 2022, 13, 887682. [Google Scholar] [CrossRef]
- Carmo-Silva, E.; Scales, J.C.; Madgwick, P.J.; Parry, M.A.J. Optimizing Rubisco and its regularion for greater resource use efficiency. Plant Cell Environ. 2014, 38, 1817–1892. [Google Scholar] [CrossRef]
- Rodrigues, V.R.; Crusciol, C.A.C.; Bossolani, J.W.; Moretti, L.G.; Portugal, J.R.; Mundt, T.T.; Oliveira, S.L.; Garcia, A.; Calonego, J.C.; Lollato, R.P. Magnesium foliar supplementation increases grain yield of soybean and maize by improving photosynthetic carbon metabolism and antioxidant metabolism. Plants 2021, 10, 797. [Google Scholar] [CrossRef]
- Rodrigues, V.A.; Crusciol, C.A.C.; Bossolani, J.W.; Portugal, J.R.; Moretti, L.G.; Bernart, L.; Vilela, R.G.; Galeriani, T.; Lollato, R.P. Foliar nitrogen as stimulant fertilization alters carbon metaboslim, reactive oxygen species scavenging, and enhances grain yield in a soybean-maize rotation. Crop Sci. 2021, 61, 3687–3701. [Google Scholar] [CrossRef]
- Garcia-González, M.; Mateo, P.; Bonilla, I. Photosynthesis and reductant sources and their relationship with nitrogenase activity in Anabaena PCC 7119. Plant Physiol. 1990, 93, 325–560. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, S.; Wan, S.; Li, X. The significance of calcium in photosynthesis. Int. J. Mol. Sci. 2019, 20, 1353. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.N.; Ramchandra, T. Water relations and photosynthesis in mustard plants subjected to boron deficiency. Indian J. Plant Physiol. 1990, 33, 150–154. [Google Scholar]
- Cakmak, I.; Römheld, V. Boron deficiency-induced impairments of cellular functions in plants. Plant Soil 1997, 193, 71–83. [Google Scholar] [CrossRef]
- Furlani, A.M.C.; Carvalho, C.P.; Freitas, J.P.; Verdial, M.F. Wheat cultivar tolerance to boron deficiency and toxicity in nutrient solution. Sci. Agric. 2003, 60, 359–370. [Google Scholar] [CrossRef]
- Goldbach, H.; Huang, L.; Wimmer, M. Boron functions in plants and animals: Recent advances in boron research and open questions. In Advances in Plant and Animal Boron Nutrition; Xu, F., Goldbach, H.E., Brown, P.H., Bell, R.W., Fujiwara, T., Hunt, C.D., Goldberg, S., Shi, L., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 3–25. [Google Scholar]
- Pandey, D.K.; Pandey, N. Screening of wheat genotypes for their susceptibility to boron deficiency. Res. Environ. Life Sci. 2008, 1, 37–42. [Google Scholar]
- Ardic, M.; Sekmen, A.H.; Tokur, S.; Ozdemir, F.; Turkan, I. Antioxidant responses of chickpea plants subjected to boron toxicity. Plant Biol. 2009, 11, 328–338. [Google Scholar] [CrossRef]
- Li, M.; Zhao, Z.; Zhang, Z.; Zhang, W.; Zhou, J.; Xu, F.; Liu, X. Effect of boron deficiency on anatomical structure and chemical composition of petioles and photosynthesis of leaves in cotton (Gossypium hirsutum L.). Sci. Rep. 2017, 7, 4420. [Google Scholar] [CrossRef]
- Landi, M.; Margaritopoulou, T.; Papadakis, I.; Araniti, F. Boron toxicity in higher 340 plants: An update. Plants 2019, 250, 1011–1032. [Google Scholar] [CrossRef] [PubMed]
- Ohto, M.; Onai, K.; Furukawa, Y.; Aoki, E.; Araki, T.; Nakamura, K. Effects of sugar on vegetative development and floral transition in Arabidopsis. Plant Physiol. 2001, 127, 252–261. [Google Scholar] [CrossRef]
- Liu, F.; Jensen, C.R.; Anderson, M.N. Pod set related to photosynthetic rate and endogenous ABA in soybeans subjected to different water regimes and exogenous ABA and BA at early reproductive stages. Ann. Bot. Lond. 2004, 94, 405–411. [Google Scholar] [CrossRef]
- White, P.J. Ion uptake mechanisms of individual cells and roots: Short-distance transport. In Marschner’s Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 7–47. [Google Scholar]
- Bogiani, J.C.; Amaro, A.C.E.; Rosolem, C.A. Carbohydrate production and transport in cotton cultivars grown under boron deficiency. Sci. Agric. 2013, 70, 442–448. [Google Scholar] [CrossRef][Green Version]
- USDA. Keys to Soil Taxonomy, 12th ed.; USDA—Serviço de Conservação de Recursos Naturais: Washington, DC, USA, 2014.
- Koppen, W.; Geiger, R. Klimate der Erde; Verlag Justus Perthes: Gotha, Germany, 1928. [Google Scholar]
- Unicamp. Center of Meteorological and Climatic Research Applied to Agriculture; Municipalities Climate of São Paulo: Botucatu, Brazil, 2020; Available online: https://www.cpa.unicamp.br/outras (accessed on 5 June 2021).
- Rolim, G.; Sentelhas, P.; Barbiere, V. Planilhas no ambiente excel para os cálculos de balanço hídricos: Normal, sequencial, de cultura e de produtividade real e potencial. Rev. Bras. De Agrometeorol. 1998, 6, 133–137. [Google Scholar]
- Quaggio, J.A.; van Raij, B. Correção da descrição do solo. In Recomendações de Adubações e Calagem para o Estado de São Paulo; Van Raij, B., Cantarella, H., Quaggio, J.A., Furlani, A.M.C., Eds.; Instituto Agronômico: Campinas, Brazil, 1997; pp. 14–19. [Google Scholar]
- Fehr, W.R.; Caviness, C.E. Iowa State University Cooperative Extension Service, Special Report 80; Iowa State University of Science and Technology: Ames, IA, USA, 1977. [Google Scholar]
- Cosmo, B.M.N.; Galeriani, T.M.; Zanetti, W.A.L. Aplicação prática da escala fenológica na cultura da soja. In Congresso Online Para Aumento De Produtividade Do Milho e Soja, 2nd ed.; COMSOJA: Santa Maria, CA, USA, 2019. [Google Scholar]
- Ambrosano, E.J.; Tanaka, R.T.; Mascarenhas, H.A.A.; van Raij, B.; Quaggio, J.A.; Cantarella, H. Leguminosas e oleaginosas. In Recomendações de Adubação e Calagem para o Estado de São Paulo, 2nd ed.; van Raij, B., Cantarella, H., Quaggio, J.A., Furlani, A.M.C., Eds.; IAC: Campinas, Brazil, 1997; pp. 189–191. [Google Scholar]
- Malavolta, E.; Vitti, G.; Oliveira, S.A. Evolution of Nutritional Status of Plants: Principles and Applications, 2nd ed.; Piracicaba: Potafós, Brazil, 1997. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Reid, C.D.; Tecido, D.T.; Fiscus, E.L.; Strain, B.R. Comparison of spectrophotometric and radioisotopic methods for the assay of rubisco in ozone-treated plants. Physiol. Plant. 1997, 101, 398–404. [Google Scholar] [CrossRef]
- SoilControl. Medidor Portátil de Umidade tipo Caneca. 2021. Available online: https://www.soilcontrol.com.br/produto/92906/medidor-portatil-de-umidade-tipo-caneca.aspx (accessed on 23 May 2021).
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Levene, H. Robust tests for equality of variances. In Contribuições para Probabilidade e Estatística: Ensaios em Honra de Ingram; Olkin, L., Ghurye, S.G., Hoefding, W., Madow, W.G., Mann, H.B., Eds.; Stanford University Press: Palo Alto, CA, USA, 1960; pp. 278–292. [Google Scholar]
- R Core Team. R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https//www.r.project.org/ (accessed on 10 April 2019).
- SAS Institute. Procedure Guide for Personal Computers; Version 5; SAS Inst.: Cary, NC, USA, 1999. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galeriani, T.M.; Neves, G.O.; Santos Ferreira, J.H.; Oliveira, R.N.; Oliveira, S.L.; Calonego, J.C.; Crusciol, C.A.C. Calcium and Boron Fertilization Improves Soybean Photosynthetic Efficiency and Grain Yield. Plants 2022, 11, 2937. https://doi.org/10.3390/plants11212937
Galeriani TM, Neves GO, Santos Ferreira JH, Oliveira RN, Oliveira SL, Calonego JC, Crusciol CAC. Calcium and Boron Fertilization Improves Soybean Photosynthetic Efficiency and Grain Yield. Plants. 2022; 11(21):2937. https://doi.org/10.3390/plants11212937
Chicago/Turabian StyleGaleriani, Tatiani Mayara, Gabriel Oliveira Neves, João Henrique Santos Ferreira, Rafael Neres Oliveira, Sirlene Lopes Oliveira, Juliano Carlos Calonego, and Carlos Alexandre Costa Crusciol. 2022. "Calcium and Boron Fertilization Improves Soybean Photosynthetic Efficiency and Grain Yield" Plants 11, no. 21: 2937. https://doi.org/10.3390/plants11212937
APA StyleGaleriani, T. M., Neves, G. O., Santos Ferreira, J. H., Oliveira, R. N., Oliveira, S. L., Calonego, J. C., & Crusciol, C. A. C. (2022). Calcium and Boron Fertilization Improves Soybean Photosynthetic Efficiency and Grain Yield. Plants, 11(21), 2937. https://doi.org/10.3390/plants11212937