Mutation S115T in IMP-Type Metallo-β-Lactamases Compensates for Decreased Expression Levels Caused by Mutation S119G

 , ,
, , 
Abstract
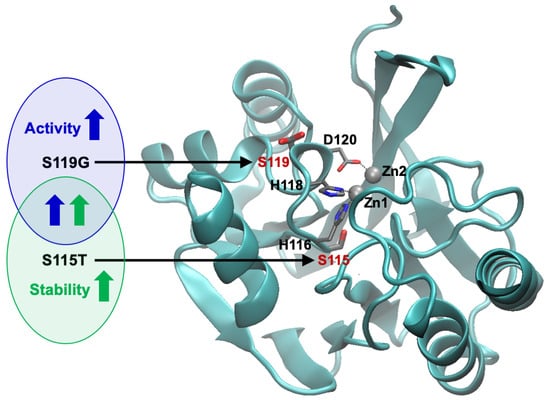
1. Introduction
2. Materials and Methods
2.1. Site-Directed Mutagenesis and Subcloning
2.2. Antibiotic Susceptibility Assays
2.2.1. Agar Disc Diffusion Assay
2.2.2. Minimum Inhibitory Concentration Assay
2.3. In-Cell Protein Expression Analysis
2.4. Protein Overexpression and Purification
2.5. Biophysical Characterization of Enzyme Variants
2.5.1. Mass Spectrometry
2.5.2. Determination of Zn(II) Content
2.5.3. Circular Dichroism Experiments
2.6. Enzyme Kinetic Assays
3. Results
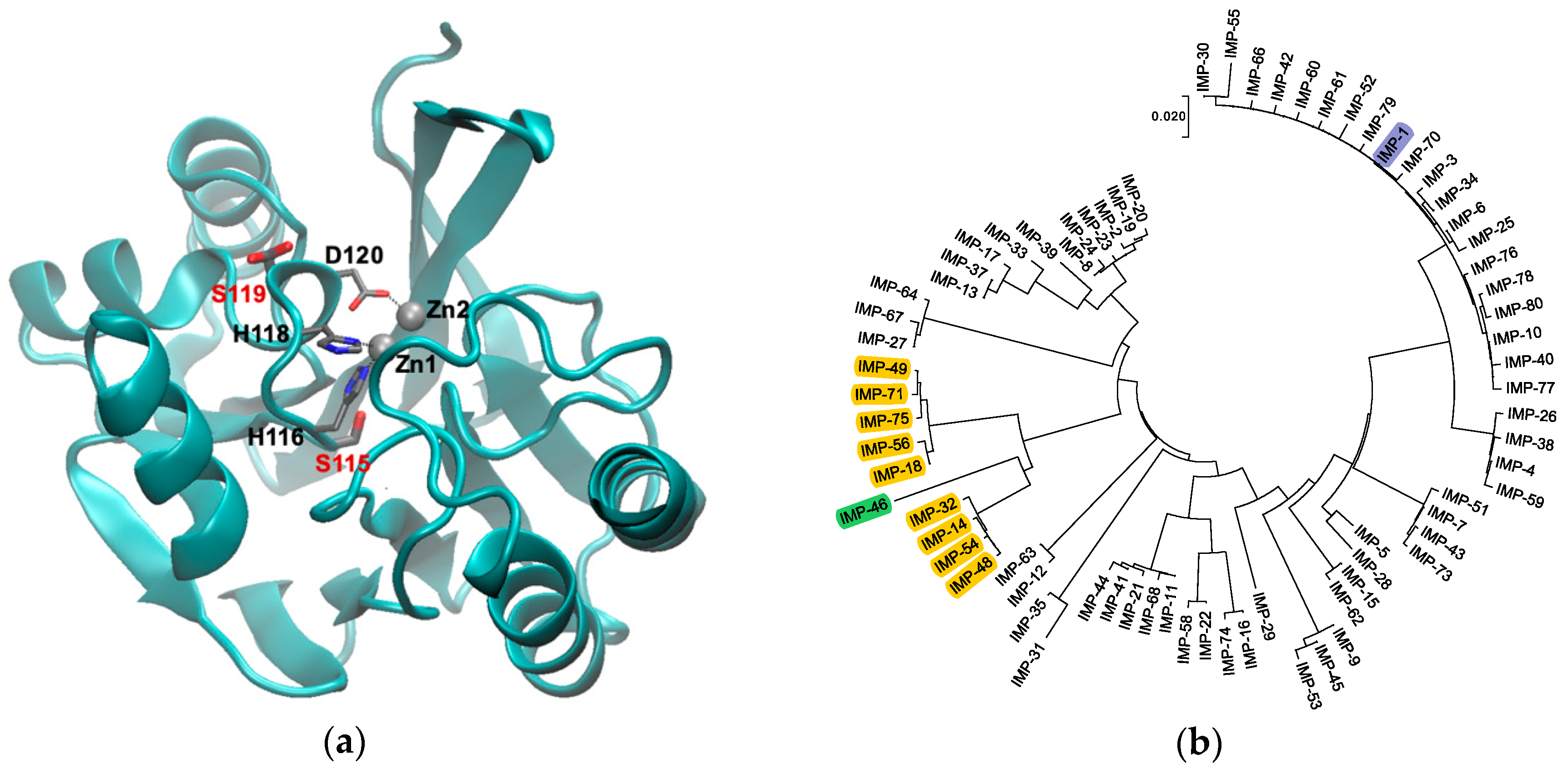
3.1. Site-Directed Mutagenesis and Subcloning
3.2. Antibiotic Susceptibility Assays
3.2.1. Agar Disc Diffusion Assay
3.2.2. Minimum Inhibitory Concentration Assay
3.3. In-Cell Expression Levels
3.4. Enzyme Overexpression, Purification, and Biophysical Characterization
3.4.1. Enzyme Overexpression and Purification
3.4.2. Mass Spectrometry
3.4.3. Zn(II) Content
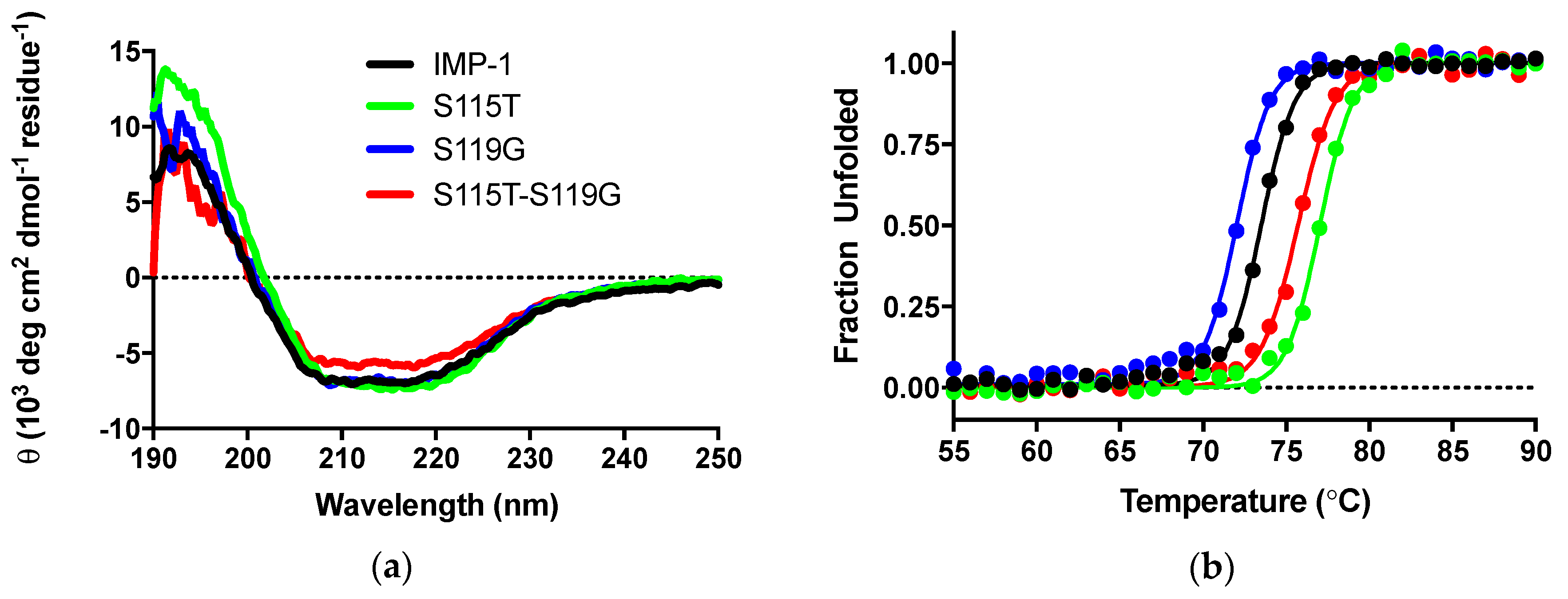
3.4.4. Circular Dichroism Experiments
3.5. Enzyme Kinetic Constants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oelschlaeger, P.; Ai, N.; Duprez, K.T.; Welsh, W.J.; Toney, J.H. Evolving carbapenemases: Can medicinal chemists advance one step ahead of the coming storm? J. Med. Chem. 2010, 53, 3013–3027. [Google Scholar] [CrossRef] [PubMed]
- Cornaglia, G.; Giamarellou, H.; Rossolini, G.M. Metallo-β-lactamases: A last frontier for β-lactams? Lancet Infect. Dis. 2011, 11, 381–393. [Google Scholar] [CrossRef]
- Walsh, T.R.; Weeks, J.; Livermore, D.M.; Toleman, M.A. Dissemination of NDM-1 positive bacteria in the New Delhi environment and its implications for human health: An environmental point prevalence study. Lancet Infect. Dis. 2011, 11, 355–362. [Google Scholar] [CrossRef]
- Zhanel, G.G.; Golden, A.R.; Zelenitsky, S.; Wiebe, K.; Lawrence, C.K.; Adam, H.J.; Idowu, T.; Domalaon, R.; Schweizer, F.; Zhanel, M.A.; et al. Cefiderocol: A Siderophore Cephalosporin with Activity Against Carbapenem-Resistant and Multidrug-Resistant Gram-Negative Bacilli. Drugs 2019, 79, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Krajnc, A.; Brem, J.; Hinchliffe, P.; Calvopina, K.; Panduwawala, T.D.; Lang, P.A.; Kamps, J.; Tyrrell, J.M.; Widlake, E.; Saward, B.G.; et al. Bicyclic Boronate VNRX-5133 Inhibits Metallo- and Serine-β-Lactamases. J. Med. Chem. 2019, 62, 8544–8556. [Google Scholar] [CrossRef]
- Lomovskaya, O. Broad-Spectrum Orally Bioavailable β-Lactamase Inhibitor QPX7728. Proceedings of ASM Microbe 2019, San Francisco, CA, USA, 21 June 2019. [Google Scholar]
- Carfi, A.; Pares, S.; Duee, E.; Galleni, M.; Duez, C.; Frere, J.M.; Dideberg, O. The 3-D structure of a zinc metallo-β-lactamase from Bacillus cereus reveals a new type of protein fold. EMBO J. 1995, 14, 4914–4921. [Google Scholar] [CrossRef]
- Concha, N.O.; Janson, C.A.; Rowling, P.; Pearson, S.; Cheever, C.A.; Clarke, B.P.; Lewis, C.; Galleni, M.; Frere, J.M.; Payne, D.J.; et al. Crystal structure of the IMP-1 metallo β-lactamase from Pseudomonas aeruginosa and its complex with a mercaptocarboxylate inhibitor: Binding determinants of a potent, broad-spectrum inhibitor. Biochemistry 2000, 39, 4288–4298. [Google Scholar] [CrossRef]
- Llarrull, L.I.; Tioni, M.F.; Kowalski, J.; Bennett, B.; Vila, A.J. Evidence for a dinuclear active site in the metallo-β-lactamase BcII with substoichiometric Co(II). A new model for metal uptake. J. Biol. Chem. 2007, 282, 30586–30595. [Google Scholar] [CrossRef]
- Garau, G.; Garcia-Saez, I.; Bebrone, C.; Anne, C.; Mercuri, P.; Galleni, M.; Frere, J.M.; Dideberg, O. Update of the standard numbering scheme for class B β-lactamases. Antimicrob. Agents Chemothe.R 2004, 48, 2347–2349. [Google Scholar] [CrossRef]
- Wang, Z.; Fast, W.; Benkovic, S.J. On the mechanism of the metallo-β-lactamase from Bacteroides fragilis. Biochemistry 1999, 38, 10013–10023. [Google Scholar] [CrossRef]
- Crowder, M.W.; Spencer, J.; Vila, A.J. Metallo-β-lactamases: Novel weaponry for antibiotic resistance in bacteria. ACC Chem. Res. 2006, 39, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Lisa, M.N.; Palacios, A.R.; Aitha, M.; Gonzalez, M.M.; Moreno, D.M.; Crowder, M.W.; Bonomo, R.A.; Spencer, J.; Tierney, D.L.; Llarrull, L.I.; et al. A general reaction mechanism for carbapenem hydrolysis by mononuclear and binuclear metallo-β-lactamases. Nat. Commun. 2017, 8, 538. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.C.; Bethel, C.R.; VanPelt, J.; Bergstrom, A.; Cheng, Z.; Miller, C.G.; Williams, C.; Poth, R.; Morris, M.; Lahey, O.; et al. Clinical Variants of New Delhi Metallo-β-Lactamase Are Evolving to Overcome Zinc Scarcity. ACS Infect. Dis. 2017, 3, 927–940. [Google Scholar] [CrossRef] [PubMed]
- Materon, I.C.; Beharry, Z.; Huang, W.; Perez, C.; Palzkill, T. Analysis of the context dependent sequence requirements of active site residues in the metallo-β-lactamase IMP-1. J. Mol. Biol. 2004, 344, 653–663. [Google Scholar] [CrossRef]
- Oelschlaeger, P.; Mayo, S.L.; Pleiss, J. Impact of remote mutations on metallo-β-lactamase substrate specificity: Implications for the evolution of antibiotic resistance. Protein Sci. 2005, 14, 765–774. [Google Scholar] [CrossRef]
- Tomatis, P.E.; Rasia, R.M.; Segovia, L.; Vila, A.J. Mimicking natural evolution in metallo-β-lactamases through second-shell ligand mutations. Proc. Natl. Acad. Sci. USA 2005, 102, 13761–13766. [Google Scholar] [CrossRef]
- Liu, E.M.; Pegg, K.M.; Oelschlaeger, P. The sequence-activity relationship between metallo-β-lactamases IMP-1, IMP-6, and IMP-25 suggests an evolutionary adaptation to meropenem exposure. Antimicrob. Agents Chemother. 2012, 56, 6403–6406. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. M100-S22. Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2013. [Google Scholar]
- Pegg, K.M.; Liu, E.M.; George, A.C.; LaCuran, A.E.; Bethel, C.R.; Bonomo, R.A.; Oelschlaeger, P. Understanding the determinants of substrate specificity in IMP family metallo-β-lactamases: The importance of residue 262. Protein Sci. 2014, 23, 1451–1460. [Google Scholar] [CrossRef]
- LaCuran, A.E.; Pegg, K.M.; Liu, E.M.; Bethel, C.R.; Ai, N.; Welsh, W.J.; Bonomo, R.A.; Oelschlaeger, P. Elucidating the Role of Residue 67 in IMP-Type Metallo-β-Lactamase Evolution. Antimicrob. Agents Chemother. 2015, 59, 7299–7307. [Google Scholar] [CrossRef]
- Goto, M.; Yasuzawa, H.; Higashi, T.; Yamaguchi, Y.; Kawanami, A.; Mifune, S.; Mori, H.; Nakayama, H.; Harada, K.; Arakawa, Y. Dependence of hydrolysis of β-lactams with a zinc(II)-β-lactamase produced from Serratia marcescens (IMP-1) on pH and concentration of zinc(II) ion: Dissociation of Zn(II) from IMP-1 in acidic medium. Biol. Pharm. Bull. 2003, 26, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Amos, B. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Humana Press Inc.: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Hunt, J.B.; Neece, S.H.; Ginsburg, A. The use of 4-(2-pyridylazo)resorcinol in studies of zinc release from Escherichia coli aspartate transcarbamoylase. Anal. Biochem. 1985, 146, 150–157. [Google Scholar] [CrossRef]
- Whitmore, L.; Wallace, B.A. DICHROWEB, an online server for protein secondary structure analyses from circular dichroism spectroscopic data. Nucleic Acids Res. 2004, 32, 668–673. [Google Scholar] [CrossRef]
- Whitmore, L.; Wallace, B.A. Protein secondary structure analyses from circular dichroism spectroscopy: Methods and reference databases. Biopolymers 2008, 89, 392–400. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Wang, W.M.; Oelschlaeger, P.; Chen, C.; Lei, J.E.; Lv, M.; Yang, K.W. Real-Time Monitoring of NDM-1 Activity in Live Bacterial Cells by Isothermal Titration Calorimetry: A New Approach to Measure Inhibition of Antibiotic-Resistant Bacteria. ACS Infect. Dis. 2018, 4, 1671–1678. [Google Scholar] [CrossRef]
- Osano, E.; Arakawa, Y.; Wacharotayankun, R.; Ohta, M.; Horii, T.; Ito, H.; Yoshimura, F.; Kato, N. Molecular characterization of an enterobacterial metallo β-lactamase found in a clinical isolate of Serratia marcescens that shows imipenem resistance. Antimicrob. Agents Chemother. 1994, 38, 71–78. [Google Scholar] [CrossRef]
- Toney, J.H.; Hammond, G.G.; Fitzgerald, P.M.; Sharma, N.; Balkovec, J.M.; Rouen, G.P.; Olson, S.H.; Hammond, M.L.; Greenlee, M.L.; Gao, Y.D. Succinic acids as potent inhibitors of plasmid-borne IMP-1 metallo-β-lactamase. J. Biol. Chem. 2001, 276, 31913–31918. [Google Scholar] [CrossRef]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-PdbViewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef]
- Malakauskas, S.M.; Mayo, S.L. Design, structure and stability of a hyperthermophilic protein variant. Nat. Struct. Biol. 1998, 5, 470–475. [Google Scholar] [CrossRef]
- Tomatis, P.E.; Fabiane, S.M.; Simona, F.; Carloni, P.; Sutton, B.J.; Vila, A.J. Adaptive protein evolution grants organismal fitness by improving catalysis and flexibility. Proc. Natl. Acad. Sci. USA 2008, 105, 20605–20610. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Huang, W.; Palzkill, T. A natural polymorphism in β-lactamase is a global suppressor. Proc. Natl. Acad. Sci. USA 1997, 94, 8801–8806. [Google Scholar] [CrossRef]
- Orencia, M.C.; Yoon, J.S.; Ness, J.E.; Stemmer, W.P.; Stevens, R.C. Predicting the emergence of antibiotic resistance by directed evolution and structural analysis. Nat. Struct. Biol. 2001, 8, 238–242. [Google Scholar] [CrossRef]
- Winkler, M.L.; Bonomo, R.A. SHV-129: A Gateway to Global Suppressors in the SHV β-Lactamase Family? Mol. Biol. Evol. 2016, 33, 429–441. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| β-Lactam | pBCSK(+) blaIMP-1 | pBCSK(+) blaIMP-1-S115T | pBCSK(+) blaIMP-1-S119G | pBCSK(+) blaIMP-1-S115T-S119G | No Vector | Empty pBCSK(+) |
|---|---|---|---|---|---|---|
| Benzylpenicillin | 128 | 64 | 128 | 256 | 32 | 32 |
| Ampicillin | 512 | 128 | 256 | ≥1024 | 4 | 4 |
| Cephalothin | 1024 | 1024 | 1024 | 512 | 16 | 16 |
| Cefoxitin | ≥1024 | ≥1024 | ≥1024 | ≥1024 | 8 | 8 |
| Cefotaxime | 128 | 128 | 64 | 128 | 8 | 8 |
| Ceftazidime | 512 | 128 | 128 | 256 | 0.063 | 0.063 |
| Imipenem | 8 | 8 | 8 | 8 | 0.25 | 0.25 |
| Meropenem | 16 | 8 | 4 | 16 | 0.125 | 0.125 |
| Doripenem | 16 | 16 | 4 | 32 | 0.125 | 0.125 |
| Aztreonam | 0.125 | 0.063 | 0.063 | 0.063 | 0.25 | 0.25 |
| Characteristic\Enzyme | IMP-1 | IMP-1-S115T | IMP-1-S119T | IMP-1-S115T-S119G |
|---|---|---|---|---|
| Relative In-Cell Expression Level (%) | 100 | 95 | 54 | 83 |
| Concentration After Purification (μM) | 93.7 | 80.4 | 47.3 | 52.9 |
| Yield of Purified Enzyme from 1 L Culture (mg) | 5.2 | 4.2 | 1.3 | 3.6 |
| Molecular Mass (measured) | 25,112 | 25,125 | 25,082 | 25,096 |
| Molecular Mass (calculated) 1 | 25,112.7 | 25,126.7 | 25,082.7 | 25,096.7 |
| Zinc Content | 2.2 ± 0.2 | 2.1 ± 0.2 | 2.1 ± 0.2 | 2.3 ± 0.1 |
| Secondary Structure Elements: 2 | ||||
| α Helix (%) | 37 | 37 | 37 | 37 |
| β Sheet (%) | 26 | 26 | 26 | 26 |
| Random Coil (%) | 38 | 38 | 38 | 38 |
| Melting Temperature (°C) 3 | 73.5 ± 0.1 | 77.0 ± 0.1 | 72.1 ± 0.1 | 75.7 ± 0.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.J.; Faheem, M.; Dang, P.; Morris, M.N.; Kumar, P.; Oelschlaeger, P. Mutation S115T in IMP-Type Metallo-β-Lactamases Compensates for Decreased Expression Levels Caused by Mutation S119G. Biomolecules 2019, 9, 724. https://doi.org/10.3390/biom9110724
Zhang CJ, Faheem M, Dang P, Morris MN, Kumar P, Oelschlaeger P. Mutation S115T in IMP-Type Metallo-β-Lactamases Compensates for Decreased Expression Levels Caused by Mutation S119G. Biomolecules. 2019; 9(11):724. https://doi.org/10.3390/biom9110724
Chicago/Turabian StyleZhang, Charles J., Mohammad Faheem, Paulie Dang, Monica N. Morris, Pooja Kumar, and Peter Oelschlaeger. 2019. "Mutation S115T in IMP-Type Metallo-β-Lactamases Compensates for Decreased Expression Levels Caused by Mutation S119G" Biomolecules 9, no. 11: 724. https://doi.org/10.3390/biom9110724
APA StyleZhang, C. J., Faheem, M., Dang, P., Morris, M. N., Kumar, P., & Oelschlaeger, P. (2019). Mutation S115T in IMP-Type Metallo-β-Lactamases Compensates for Decreased Expression Levels Caused by Mutation S119G. Biomolecules, 9(11), 724. https://doi.org/10.3390/biom9110724