

Triggering Pyroptosis in Cancer

Abstract
1. Introduction to Pyroptosis
2. Pyroptosis Signaling Pathways
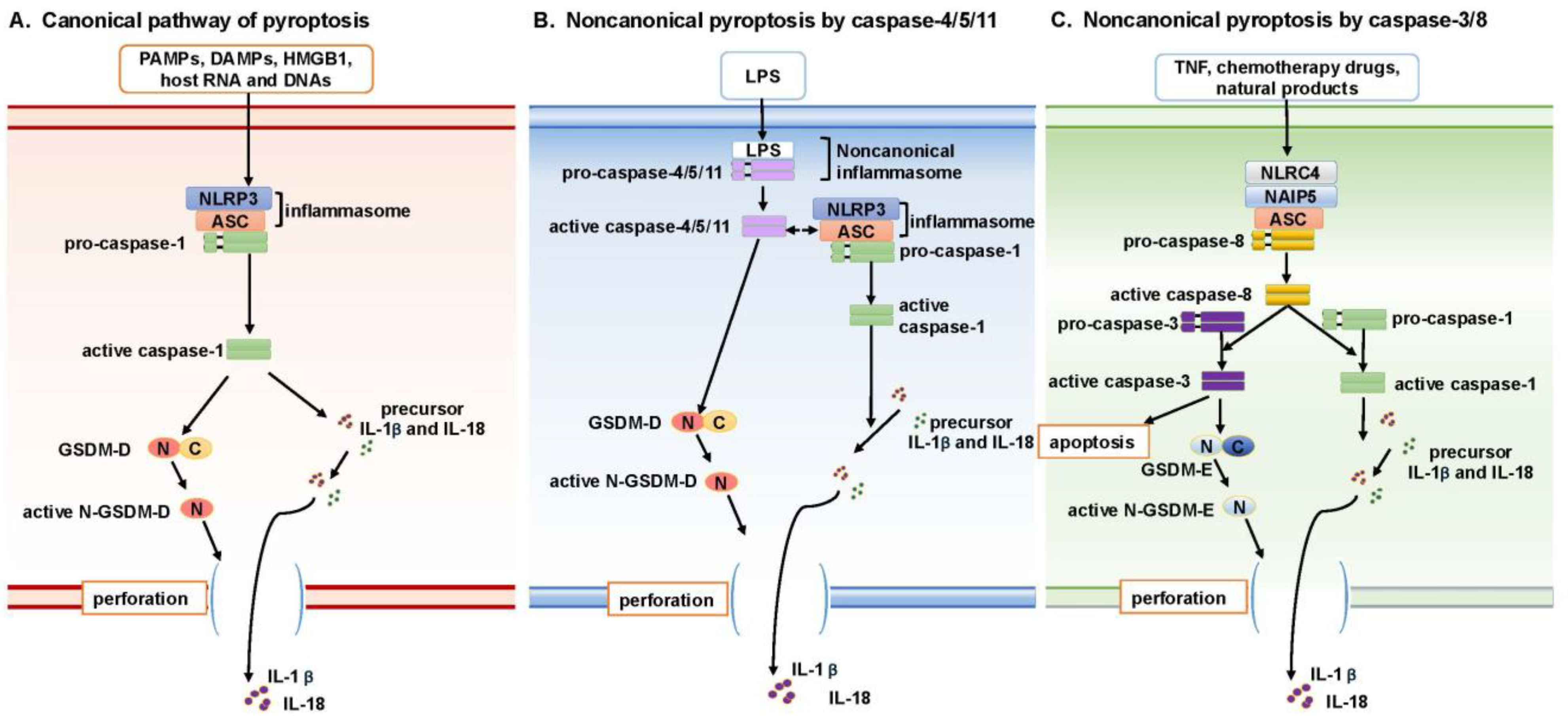
2.1. Canonical Pathway of Pyroptosis
2.2. Noncanonical Pyroptosis Mediated by Caspase-4/5/11
2.3. Noncanonical Pyroptosis Mediated by Caspase-3/8
3. Inducing Pyroptosis in Cancer
3.1. Chemotherapy Drugs That Induce Pyroptosis
3.2. Metformin
3.3. Small Molecule Inhibitors That Induce Pyroptosis
3.3.1. DPP8/9 Inhibitors
3.3.2. BRD4 Inhibitor
3.3.3. Other Inhibitors
3.4. Natural Products That Induce Pyroptosis
3.4.1. Curcumin
3.4.2. Iron
3.4.3. Cucurbitacin B
3.5. Pathogen or Pathogen-Derived Compounds That Induce Pyroptosis
Lipopolysaccharide
3.6. Endogenous Metabolites That Induce Pyroptosis
4. Challenges and Opportunities
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cookson, B.T.; Brennan, M.A. Pro-inflammatory programmed cell death. Trends Microbiol. 2001, 9, 113–114. [Google Scholar] [CrossRef] [PubMed]
- Hersh, D.; Monack, D.M.; Smith, M.R.; Ghori, N.; Falkow, S.; Zychlinsky, A. The Salmonella invasin SipB induces macrophage apoptosis by binding to caspase-1. Proc. Natl. Acad. Sci. USA 1999, 96, 2396–2401. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.A.; Cookson, B.T. Salmonella induces macrophage death by caspase-1-dependent necrosis. Mol. Microbiol. 2000, 38, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Arend, W.P.; Palmer, G.; Gabay, C. IL-1, IL-18, and IL-33 families of cytokines. Immunol. Rev. 2008, 223, 20–38. [Google Scholar] [CrossRef]
- Zychlinsky, A.; Prevost, M.C.; Sansonetti, P.J. Shigella flexneri induces apoptosis in infected macrophages. Nature 1992, 358, 167–169. [Google Scholar] [CrossRef]
- Man, S.M.; Hopkins, L.J.; Nugent, E.; Cox, S.; Gluck, I.M.; Tourlomousis, P.; Wright, J.A.; Cicuta, P.; Monie, T.P.; Bryant, C.E. Inflammasome activation causes dual recruitment of NLRC4 and NLRP3 to the same macromolecular complex. Proc. Natl. Acad. Sci. USA 2014, 111, 7403–7408. [Google Scholar] [CrossRef]
- Oh, C.; Spears, T.J.; Aachoui, Y. Inflammasome-mediated pyroptosis in defense against pathogenic bacteria. Immunol. Rev. 2024, 329, e13408. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, J.; Yu, S.; Li, Y.; Zhu, J.; Zhang, K.; Zhang, R. Cell pyroptosis in health and inflammatory diseases. Cell Death Discov. 2022, 8, 191. [Google Scholar] [CrossRef]
- Zhaolin, Z.; Guohua, L.; Shiyuan, W.; Zuo, W. Role of pyroptosis in cardiovascular disease. Cell Prolif. 2019, 52, e12563. [Google Scholar] [CrossRef]
- Dinarello, C.A. Immunological and inflammatory functions of the interleukin-1 family. Annu. Rev. Immunol. 2009, 27, 519–550. [Google Scholar] [CrossRef]
- Jorgensen, I.; Lopez, J.P.; Laufer, S.A.; Miao, E.A. IL-1beta, IL-18, and eicosanoids promote neutrophil recruitment to pore-induced intracellular traps following pyroptosis. Eur. J. Immunol. 2016, 46, 2761–2766. [Google Scholar] [CrossRef] [PubMed]
- Cordoba-Rodriguez, R.; Fang, H.; Lankford, C.S.; Frucht, D.M. Anthrax lethal toxin rapidly activates caspase-1/ICE and induces extracellular release of interleukin (IL)-1beta and IL-18. J. Biol. Chem. 2004, 279, 20563–20566. [Google Scholar] [CrossRef] [PubMed]
- Boyden, E.D.; Dietrich, W.F. Nalp1b controls mouse macrophage susceptibility to anthrax lethal toxin. Nat. Genet. 2006, 38, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M.; Kanneganti, T.D. Innate immune sensing of cell death in disease and therapeutics. Nat. Cell Biol. 2024, 26, 1420–1433. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Coyne, C.B.; Zeh, H.J.; Lotze, M.T. PAMPs and DAMPs: Signal 0s that spur autophagy and immunity. Immunol. Rev. 2012, 249, 158–175. [Google Scholar] [CrossRef]
- Wei, X.; Xie, F.; Zhou, X.; Wu, Y.; Yan, H.; Liu, T.; Huang, J.; Wang, F.; Zhou, F.; Zhang, L. Role of pyroptosis in inflammation and cancer. Cell Mol. Immunol. 2022, 19, 971–992. [Google Scholar] [CrossRef]
- Jin, C.; Flavell, R.A. Molecular mechanism of NLRP3 inflammasome activation. J. Clin. Immunol. 2010, 30, 628–631. [Google Scholar] [CrossRef]
- Ding, J.; Wang, K.; Liu, W.; She, Y.; Sun, Q.; Shi, J.; Sun, H.; Wang, D.C.; Shao, F. Pore-forming activity and structural autoinhibition of the gasdermin family. Nature 2016, 535, 111–116. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Z.; Ruan, J.; Pan, Y.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 2016, 535, 153–158. [Google Scholar] [CrossRef]
- Chen, K.W.; Broz, P. Gasdermins as evolutionarily conserved executors of inflammation and cell death. Nat. Cell Biol. 2024, 26, 1394–1406. [Google Scholar] [CrossRef]
- Dinarello, C.A. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef]
- Evavold, C.L.; Ruan, J.; Tan, Y.; Xia, S.; Wu, H.; Kagan, J.C. The Pore-Forming Protein Gasdermin D Regulates Interleukin-1 Secretion from Living Macrophages. Immunity 2018, 48, 35–44 e36. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Control of adaptive immunity by the innate immune system. Nat. Immunol. 2015, 16, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Van Den Eeckhout, B.; Tavernier, J.; Gerlo, S. Interleukin-1 as Innate Mediator of T Cell Immunity. Front. Immunol. 2020, 11, 621931. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, Y.; Wang, Y.; Gao, W.; Ding, J.; Li, P.; Hu, L.; Shao, F. Inflammatory caspases are innate immune receptors for intracellular LPS. Nature 2014, 514, 187–192. [Google Scholar] [CrossRef]
- Kayagaki, N.; Warming, S.; Lamkanfi, M.; Vande Walle, L.; Louie, S.; Dong, J.; Newton, K.; Qu, Y.; Liu, J.; Heldens, S.; et al. Non-canonical inflammasome activation targets caspase-11. Nature 2011, 479, 117–121. [Google Scholar] [CrossRef]
- Kayagaki, N.; Stowe, I.B.; Lee, B.L.; O’Rourke, K.; Anderson, K.; Warming, S.; Cuellar, T.; Haley, B.; Roose-Girma, M.; Phung, Q.T.; et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 2015, 526, 666–671. [Google Scholar] [CrossRef]
- Zhu, C.; Xu, S.; Jiang, R.; Yu, Y.; Bian, J.; Zou, Z. The gasdermin family: Emerging therapeutic targets in diseases. Signal Transduct. Target. Ther. 2024, 9, 87. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, W.; Shi, X.; Ding, J.; Liu, W.; He, H.; Wang, K.; Shao, F. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature 2017, 547, 99–103. [Google Scholar] [CrossRef]
- Mascarenhas, D.P.A.; Cerqueira, D.M.; Pereira, M.S.F.; Castanheira, F.V.S.; Fernandes, T.D.; Manin, G.Z.; Cunha, L.D.; Zamboni, D.S. Inhibition of caspase-1 or gasdermin-D enable caspase-8 activation in the Naip5/NLRC4/ASC inflammasome. PLoS Pathog. 2017, 13, e1006502. [Google Scholar] [CrossRef]
- Fritsch, M.; Gunther, S.D.; Schwarzer, R.; Albert, M.C.; Schorn, F.; Werthenbach, J.P.; Schiffmann, L.M.; Stair, N.; Stocks, H.; Seeger, J.M.; et al. Caspase-8 is the molecular switch for apoptosis, necroptosis and pyroptosis. Nature 2019, 575, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Orning, P.; Weng, D.; Starheim, K.; Ratner, D.; Best, Z.; Lee, B.; Brooks, A.; Xia, S.; Wu, H.; Kelliher, M.A.; et al. Pathogen blockade of TAK1 triggers caspase-8-dependent cleavage of gasdermin D and cell death. Science 2018, 362, 1064–1069. [Google Scholar] [CrossRef] [PubMed]
- Pazhouhesh Far, N.; Hajiheidari Varnousafaderani, M.; Faghihkhorasani, F.; Etemad, S.; Abdulwahid, A.R.R.; Bakhtiarinia, N.; Mousaei, A.; Dortaj, E.; Karimi, S.; Ebrahimi, N.; et al. Breaking the barriers: Overcoming cancer resistance by targeting the NLRP3 inflammasome. Br. J. Pharmacol. 2024, 182, 3–25. [Google Scholar] [CrossRef]
- Wu, X.; Cao, J.; Wan, X.; Du, S. Programmed cell death in hepatocellular carcinoma: Mechanisms and therapeutic prospects. Cell Death Discov. 2024, 10, 356. [Google Scholar] [CrossRef]
- Yan, H.; Luo, B.; Wu, X.; Guan, F.; Yu, X.; Zhao, L.; Ke, X.; Wu, J.; Yuan, J. Cisplatin Induces Pyroptosis via Activation of MEG3/NLRP3/caspase-1/GSDMD Pathway in Triple-Negative Breast Cancer. Int. J. Biol. Sci. 2021, 17, 2606–2621. [Google Scholar] [CrossRef] [PubMed]
- Li, R.Y.; Zheng, Z.Y.; Li, Z.M.; Heng, J.H.; Zheng, Y.Q.; Deng, D.X.; Xu, X.E.; Liao, L.D.; Lin, W.; Xu, H.Y.; et al. Cisplatin-induced pyroptosis is mediated via the CAPN1/CAPN2-BAK/BAX-caspase-9-caspase-3-GSDME axis in esophageal cancer. Chem. Biol. Interact. 2022, 361, 109967. [Google Scholar] [CrossRef]
- Zhang, C.C.; Li, C.G.; Wang, Y.F.; Xu, L.H.; He, X.H.; Zeng, Q.Z.; Zeng, C.Y.; Mai, F.Y.; Hu, B.; Ouyang, D.Y. Chemotherapeutic paclitaxel and cisplatin differentially induce pyroptosis in A549 lung cancer cells via caspase-3/GSDME activation. Apoptosis 2019, 24, 312–325. [Google Scholar] [CrossRef]
- Yu, J.; Li, S.; Qi, J.; Chen, Z.; Wu, Y.; Guo, J.; Wang, K.; Sun, X.; Zheng, J. Cleavage of GSDME by caspase-3 determines lobaplatin-induced pyroptosis in colon cancer cells. Cell Death Dis. 2019, 10, 193. [Google Scholar] [CrossRef]
- Chen, J.; Ge, L.; Shi, X.; Liu, J.; Ruan, H.; Heng, D.; Ye, C. Lobaplatin Induces Pyroptosis in Cervical Cancer Cells via the Caspase-3/GSDME Pathway. Anticancer. Agents Med. Chem. 2022, 22, 2091–2097. [Google Scholar] [CrossRef]
- Peng, Z.; Wang, P.; Song, W.; Yao, Q.; Li, Y.; Liu, L.; Li, Y.; Zhou, S. GSDME enhances Cisplatin sensitivity to regress non-small cell lung carcinoma by mediating pyroptosis to trigger antitumor immunocyte infiltration. Signal Transduct. Target. Ther. 2020, 5, 159. [Google Scholar] [CrossRef]
- Wang, Y.; Yin, B.; Li, D.; Wang, G.; Han, X.; Sun, X. GSDME mediates caspase-3-dependent pyroptosis in gastric cancer. Biochem. Biophys. Res. Commun. 2018, 495, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Guo, S.; Tang, X.; Rong, Y.; Bo, H.; Shen, H.; Zhao, Z.; Qiao, A.; Shen, J.; Wang, J. Combination of ruthenium (II) polypyridyl complex Delta-Ru1 and Taxol enhances the anti-cancer effect on Taxol-resistant cancer cells through Caspase-1/GSDMD-mediated pyroptosis. J. Inorg. Biochem. 2022, 230, 111749. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Ying, C.; Chen, Y.; Huang, F.; Wang, B.; Fang, H.; Guo, W.; Liu, T.; Zhou, X.; Huang, B.; et al. Doxorubicin hydrochloride enhanced antitumour effect of CEA-regulated oncolytic virotherapy in live cancer cells and a mouse model. J. Cell Mol. Med. 2020, 24, 13431–13439. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Ji, L.; Han, M.; Xie, J.; Zhong, F.; Zhang, X.; Su, Q.; Yang, Z.; Liu, Z.; Gao, H.; et al. Pyroptosis is involved in the inhibitory effect of FL118 on growth and metastasis in colorectal cancer. Life Sci. 2020, 257, 118065. [Google Scholar] [CrossRef]
- Wang, L.; Li, K.; Lin, X.; Yao, Z.; Wang, S.; Xiong, X.; Ning, Z.; Wang, J.; Xu, X.; Jiang, Y.; et al. Metformin induces human esophageal carcinoma cell pyroptosis by targeting the miR-497/PELP1 axis. Cancer Lett. 2019, 450, 22–31. [Google Scholar] [CrossRef]
- Zheng, Z.; Bian, Y.; Zhang, Y.; Ren, G.; Li, G. Metformin activates AMPK/SIRT1/NF-kappaB pathway and induces mitochondrial dysfunction to drive caspase3/GSDME-mediated cancer cell pyroptosis. Cell Cycle 2020, 19, 1089–1104. [Google Scholar] [CrossRef]
- Okondo, M.C.; Johnson, D.C.; Sridharan, R.; Go, E.B.; Chui, A.J.; Wang, M.S.; Poplawski, S.E.; Wu, W.; Liu, Y.; Lai, J.H.; et al. DPP8 and DPP9 inhibition induces pro-caspase-1-dependent monocyte and macrophage pyroptosis. Nat. Chem. Biol. 2017, 13, 46–53. [Google Scholar] [CrossRef]
- Karakaya, T.; Slaufova, M.; Di Filippo, M.; Hennig, P.; Kundig, T.; Beer, H.D. CARD8: A Novel Inflammasome Sensor with Well-Known Anti-Inflammatory and Anti-Apoptotic Activity. Cells 2024, 13, 1032. [Google Scholar] [CrossRef]
- de Vasconcelos, N.M.; Vliegen, G.; Goncalves, A.; De Hert, E.; Martin-Perez, R.; Van Opdenbosch, N.; Jallapally, A.; Geiss-Friedlander, R.; Lambeir, A.M.; Augustyns, K.; et al. DPP8/DPP9 inhibition elicits canonical Nlrp1b inflammasome hallmarks in murine macrophages. Life Sci. Alliance 2019, 2, 313. [Google Scholar] [CrossRef]
- Coutts, S.J.; Kelly, T.A.; Snow, R.J.; Kennedy, C.A.; Barton, R.W.; Adams, J.; Krolikowski, D.A.; Freeman, D.M.; Campbell, S.J.; Ksiazek, J.F.; et al. Structure-activity relationships of boronic acid inhibitors of dipeptidyl peptidase IV. 1. Variation of the P2 position of Xaa-boroPro dipeptides. J. Med. Chem. 1996, 39, 2087–2094. [Google Scholar] [CrossRef]
- Lankas, G.R.; Leiting, B.; Roy, R.S.; Eiermann, G.J.; Beconi, M.G.; Biftu, T.; Chan, C.C.; Edmondson, S.; Feeney, W.P.; He, H.; et al. Dipeptidyl peptidase IV inhibition for the treatment of type 2 diabetes: Potential importance of selectivity over dipeptidyl peptidases 8 and 9. Diabetes 2005, 54, 2988–2994. [Google Scholar] [CrossRef] [PubMed]
- Van Goethem, S.; Van der Veken, P.; Dubois, V.; Soroka, A.; Lambeir, A.M.; Chen, X.; Haemers, A.; Scharpe, S.; De Meester, I.; Augustyns, K. Inhibitors of dipeptidyl peptidase 8 and dipeptidyl peptidase 9. Part 2: Isoindoline containing inhibitors. Bioorg. Med. Chem. Lett. 2008, 18, 4159–4162. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.D.; Chen, Q.; Wang, Q.; Orth-He, E.L.; Saoi, M.; Griswold, A.R.; Bhattacharjee, A.; Ball, D.P.; Huang, H.C.; Chui, A.J.; et al. M24B aminopeptidase inhibitors selectively activate the CARD8 inflammasome. Nat. Chem. Biol. 2022, 18, 565–574. [Google Scholar] [CrossRef]
- Johnson, D.C.; Taabazuing, C.Y.; Okondo, M.C.; Chui, A.J.; Rao, S.D.; Brown, F.C.; Reed, C.; Peguero, E.; de Stanchina, E.; Kentsis, A.; et al. DPP8/DPP9 inhibitor-induced pyroptosis for treatment of acute myeloid leukemia. Nat. Med. 2018, 24, 1151–1156. [Google Scholar] [CrossRef]
- Tan, Y.F.; Wang, M.; Chen, Z.Y.; Wang, L.; Liu, X.H. Inhibition of BRD4 prevents proliferation and epithelial-mesenchymal transition in renal cell carcinoma via NLRP3 inflammasome-induced pyroptosis. Cell Death Dis. 2020, 11, 239. [Google Scholar] [CrossRef]
- Dong, X.; Hu, X.; Bao, Y.; Li, G.; Yang, X.D.; Slauch, J.M.; Chen, L.F. Brd4 regulates NLRC4 inflammasome activation by facilitating IRF8-mediated transcription of Naips. J. Cell Biol. 2021, 220, e202005148. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, S.; Wu, J.; Chen, M.; Cai, M.C.; Fu, Y.; Li, W.; Wang, J.; Zhao, X.; Yu, Z.; et al. Molecular Targeted Therapies Elicit Concurrent Apoptotic and GSDME-Dependent Pyroptotic Tumor Cell Death. Clin. Cancer Res. 2018, 24, 6066–6077. [Google Scholar] [CrossRef]
- Erkes, D.A.; Cai, W.; Sanchez, I.M.; Purwin, T.J.; Rogers, C.; Field, C.O.; Berger, A.C.; Hartsough, E.J.; Rodeck, U.; Alnemri, E.S.; et al. Mutant BRAF and MEK Inhibitors Regulate the Tumor Immune Microenvironment via Pyroptosis. Cancer Discov. 2020, 10, 254–269. [Google Scholar] [CrossRef]
- Wu, M.; Wang, Y.; Yang, D.; Gong, Y.; Rao, F.; Liu, R.; Danna, Y.; Li, J.; Fan, J.; Chen, J.; et al. A PLK1 kinase inhibitor enhances the chemosensitivity of cisplatin by inducing pyroptosis in oesophageal squamous cell carcinoma. EBioMedicine 2019, 41, 244–255. [Google Scholar] [CrossRef]
- Huo, J.; Shen, Y.; Zhang, Y.; Shen, L. BI 2536 induces gasdermin E-dependent pyroptosis in ovarian cancer. Front. Oncol. 2022, 12, 963928. [Google Scholar] [CrossRef]
- Qiao, L.; Wu, X.; Zhang, J.; Liu, L.; Sui, X.; Zhang, R.; Liu, W.; Shen, F.; Sun, Y.; Xi, X. alpha-NETA induces pyroptosis of epithelial ovarian cancer cells through the GSDMD/caspase-4 pathway. FASEB J. 2019, 33, 12760–12767. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.B.; Jiao, B.P.; Liu, Y.J.; Li, Y.R.; Wang, G.J. BIX-01294 enhanced chemotherapy effect in gastric cancer by inducing GSDME-mediated pyroptosis. Cell Biol. Int. 2020, 44, 1890–1899. [Google Scholar] [CrossRef] [PubMed]
- Long, K.; Gu, L.; Li, L.; Zhang, Z.; Li, E.; Zhang, Y.; He, L.; Pan, F.; Guo, Z.; Hu, Z. Small-molecule inhibition of APE1 induces apoptosis, pyroptosis, and necroptosis in non-small cell lung cancer. Cell Death Dis. 2021, 12, 503. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Fan, P.; Liu, M.; Weng, C.; Fan, G.; Zhang, T.; Duan, X.; Wu, Y.; Tang, L.; Yang, G.; et al. Famotidine promotes inflammation by triggering cell pyroptosis in gastric cancer cells. BMC Pharmacol. Toxicol. 2021, 22, 62. [Google Scholar] [CrossRef]
- Kobayashi, T.; Mitsuhashi, A.; Hongying, P.; Shioya, M.; Kojima, K.; Nishikimi, K.; Yahiro, K.; Shozu, M. Bexarotene-induced cell death in ovarian cancer cells through Caspase-4-gasdermin E mediated pyroptosis. Sci. Rep. 2022, 12, 11123. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, L.; Zhang, Y.; Jiang, Z.; Lu, H.; Xie, Y.; Han, W.; Zhao, W.; He, J.; Shi, Z.; et al. The CDK inhibitor AT7519 inhibits human glioblastoma cell growth by inducing apoptosis, pyroptosis and cell cycle arrest. Cell Death Dis. 2023, 14, 11. [Google Scholar] [CrossRef]
- Huntington, K.E.; Louie, A.D.; Srinivasan, P.R.; Schorl, C.; Lu, S.; Silverberg, D.; Newhouse, D.; Wu, Z.; Zhou, L.; Borden, B.A.; et al. GSK-3 inhibitor elraglusib enhances tumor-infiltrating immune cell activation in tumor biopsies and synergizes with anti-PD-L1 in a murine model of colorectal cancer. bioRxiv 2023. [Google Scholar] [CrossRef]
- Prasad, S.; Tyagi, A.K.; Aggarwal, B.B. Recent developments in delivery, bioavailability, absorption and metabolism of curcumin: The golden pigment from golden spice. Cancer Res. Treat. 2014, 46, 2–18. [Google Scholar] [CrossRef]
- Alexandrow, M.G.; Song, L.J.; Altiok, S.; Gray, J.; Haura, E.B.; Kumar, N.B. Curcumin: A novel Stat3 pathway inhibitor for chemoprevention of lung cancer. Eur. J. Cancer Prev. 2012, 21, 407–412. [Google Scholar] [CrossRef]
- Panahi, Y.; Darvishi, B.; Ghanei, M.; Jowzi, N.; Beiraghdar, F.; Varnamkhasti, B.S. Molecular mechanisms of curcumins suppressing effects on tumorigenesis, angiogenesis and metastasis, focusing on NF-kappaB pathway. Cytokine Growth Factor. Rev. 2016, 28, 21–29. [Google Scholar] [CrossRef]
- Cho, J.W.; Lee, K.S.; Kim, C.W. Curcumin attenuates the expression of IL-1beta, IL-6, and TNF-alpha as well as cyclin E in TNF-alpha-treated HaCaT cells; NF-kappaB and MAPKs as potential upstream targets. Int. J. Mol. Med. 2007, 19, 469–474. [Google Scholar] [PubMed]
- Dhillon, N.; Aggarwal, B.B.; Newman, R.A.; Wolff, R.A.; Kunnumakkara, A.B.; Abbruzzese, J.L.; Ng, C.S.; Badmaev, V.; Kurzrock, R. Phase II trial of curcumin in patients with advanced pancreatic cancer. Clin. Cancer Res. 2008, 14, 4491–4499. [Google Scholar] [CrossRef] [PubMed]
- Kanai, M.; Yoshimura, K.; Asada, M.; Imaizumi, A.; Suzuki, C.; Matsumoto, S.; Nishimura, T.; Mori, Y.; Masui, T.; Kawaguchi, Y.; et al. A phase I/II study of gemcitabine-based chemotherapy plus curcumin for patients with gemcitabine-resistant pancreatic cancer. Cancer Chemother. Pharmacol. 2011, 68, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Passildas-Jahanmohan, J.; Eymard, J.C.; Pouget, M.; Kwiatkowski, F.; Van Praagh, I.; Savareux, L.; Atger, M.; Durando, X.; Abrial, C.; Richard, D.; et al. Multicenter randomized phase II study comparing docetaxel plus curcumin versus docetaxel plus placebo in first-line treatment of metastatic castration-resistant prostate cancer. Cancer Med. 2021, 10, 2332–2340. [Google Scholar] [CrossRef]
- Liang, W.F.; Gong, Y.X.; Li, H.F.; Sun, F.L.; Li, W.L.; Chen, D.Q.; Xie, D.P.; Ren, C.X.; Guo, X.Y.; Wang, Z.Y.; et al. Curcumin Activates ROS Signaling to Promote Pyroptosis in Hepatocellular Carcinoma HepG2 Cells. In Vivo 2021, 35, 249–257. [Google Scholar] [CrossRef]
- Yue, E.; Tuguzbaeva, G.; Chen, X.; Qin, Y.; Li, A.; Sun, X.; Dong, C.; Liu, Y.; Yu, Y.; Zahra, S.M.; et al. Anthocyanin is involved in the activation of pyroptosis in oral squamous cell carcinoma. Phytomedicine 2019, 56, 286–294. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, P.; An, L.; Sun, N.; Peng, L.; Tang, W.; Ma, D.; Chen, J. Miltirone induces cell death in hepatocellular carcinoma cell through GSDME-dependent pyroptosis. Acta Pharm. Sin. B 2020, 10, 1397–1413. [Google Scholar] [CrossRef]
- An, H.; Heo, J.S.; Kim, P.; Lian, Z.; Lee, S.; Park, J.; Hong, E.; Pang, K.; Park, Y.; Ooshima, A.; et al. Tetraarsenic hexoxide enhances generation of mitochondrial ROS to promote pyroptosis by inducing the activation of caspase-3/GSDME in triple-negative breast cancer cells. Cell Death Dis. 2021, 12, 159. [Google Scholar] [CrossRef]
- Cai, J.; Yi, M.; Tan, Y.; Li, X.; Li, G.; Zeng, Z.; Xiong, W.; Xiang, B. Natural product triptolide induces GSDME-mediated pyroptosis in head and neck cancer through suppressing mitochondrial hexokinase-IotaIota. J. Exp. Clin. Cancer Res. 2021, 40, 190. [Google Scholar] [CrossRef]
- Tan, Y.; Xiang, J.; Huang, Z.; Wang, L.; Huang, Y. Trichosanthin inhibits cell growth and metastasis by promoting pyroptosis in non-small cell lung cancer. J. Thorac. Dis. 2022, 14, 1193–1202. [Google Scholar] [CrossRef]
- Li, T.; Shi, L.; Liu, W.; Hu, X.; Hui, Y.; Di, M.; Xue, S.; Zheng, Y.; Yao, M.; Li, C.; et al. Aloe-Emodin Induces Mitochondrial Dysfunction and Pyroptosis by Activation of the Caspase-9/3/Gasdermin E Axis in HeLa Cells. Front. Pharmacol. 2022, 13, 854526. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Cheng, C.; Zhang, J.; Fang, J.; Yao, W.; Zhu, Y.; Xiu, Z.; Jin, N.; Lu, H.; Li, X.; et al. Myricetin activates the Caspase-3/GSDME pathway via ER stress induction of pyroptosis in lung cancer cells. Front. Pharmacol. 2022, 13, 959938. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ma, S.; Pi, D.; Wu, Y.; Zuo, Q.; Li, C.; Ouyang, M. Luteolin induces pyroptosis in HT-29 cells by activating the Caspase1/Gasdermin D signalling pathway. Front. Pharmacol. 2022, 13, 952587. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Sun, F.; Deng, K.; Lin, G.; Yin, W.; Chen, H.; Yang, D.; Liu, K.; Zhang, Y.; Huang, L. Mallotucin D, a Clerodane Diterpenoid from Croton crassifolius, Suppresses HepG2 Cell Growth via Inducing Autophagic Cell Death and Pyroptosis. Int. J. Mol. Sci. 2022, 23, 14217. [Google Scholar] [CrossRef]
- Ju, X.; Zhang, H.; Wang, J.; Sun, Z.; Guo, L.; Wang, Q. Shikonin triggers GSDME-mediated pyroptosis in tumours by regulating autophagy via the ROS-MAPK14/p38alpha axis. Phytomedicine 2023, 109, 154596. [Google Scholar] [CrossRef]
- Lee, Y.J.; Park, K.S.; Lee, S.H. Curcumin Targets Both Apoptosis and Necroptosis in Acidity-Tolerant Prostate Carcinoma Cells. Biomed. Res. Int. 2021, 2021, 8859181. [Google Scholar] [CrossRef]
- Li, R.; Zhang, J.; Zhou, Y.; Gao, Q.; Wang, R.; Fu, Y.; Zheng, L.; Yu, H. Transcriptome Investigation and In Vitro Verification of Curcumin-Induced HO-1 as a Feature of Ferroptosis in Breast Cancer Cells. Oxid. Med. Cell Longev. 2020, 2020, 3469840. [Google Scholar] [CrossRef]
- Dixon, S.J.; Stockwell, B.R. The role of iron and reactive oxygen species in cell death. Nat. Chem. Biol. 2014, 10, 9–17. [Google Scholar] [CrossRef]
- Zhou, B.; Zhang, J.Y.; Liu, X.S.; Chen, H.Z.; Ai, Y.L.; Cheng, K.; Sun, R.Y.; Zhou, D.; Han, J.; Wu, Q. Tom20 senses iron-activated ROS signaling to promote melanoma cell pyroptosis. Cell Res. 2018, 28, 1171–1185. [Google Scholar] [CrossRef]
- Yuan, R.; Zhao, W.; Wang, Q.Q.; He, J.; Han, S.; Gao, H.; Feng, Y.; Yang, S. Cucurbitacin B inhibits non-small cell lung cancer in vivo and in vitro by triggering TLR4/NLRP3/GSDMD-dependent pyroptosis. Pharmacol. Res. 2021, 170, 105748. [Google Scholar] [CrossRef]
- Lou, C.; Fang, Y.; Mei, Y.; Hu, W.; Sun, L.; Jin, C.; Chen, H.; Zheng, W. Cucurbitacin B attenuates osteoarthritis development by inhibiting NLRP3 inflammasome activation and pyroptosis through activating Nrf2/HO-1 pathway. Phytother. Res. 2024, 38, 3352–3369. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.S.; Liu, Y.; Wang, X.W.; Xu, B.; Lin, Y.L.; Song, Y.; Dong, Y.; Liu, J.L.; Wang, X.J.; Liu, S.; et al. LPS Enhances the Chemosensitivity of Oxaliplatin in HT29 Cells via GSDMD-Mediated Pyroptosis. Cancer Manag. Res. 2020, 12, 10397–10409. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ma, X.; Liu, W.; Hu, Q.; Yang, H. Targeting Pyroptosis through Lipopolysaccharide-Triggered Noncanonical Pathway for Safe and Efficient Cancer Immunotherapy. Nano Lett. 2023, 23, 8725–8733. [Google Scholar] [CrossRef] [PubMed]
- Sannino, F.; Sansone, C.; Galasso, C.; Kildgaard, S.; Tedesco, P.; Fani, R.; Marino, G.; de Pascale, D.; Ianora, A.; Parrilli, E.; et al. Pseudoalteromonas haloplanktis TAC125 produces 4-hydroxybenzoic acid that induces pyroptosis in human A459 lung adenocarcinoma cells. Sci. Rep. 2018, 8, 1190. [Google Scholar] [CrossRef]
- Zhu, X.; Wu, T.; Chi, Y.; Ge, Y.; Wu, B.; Zhou, M.; Zhu, F.; Ji, M.; Cui, L. Pyroptosis induced by enterovirus A71 infection in cultured human neuroblastoma cells. Virology 2018, 521, 69–76. [Google Scholar] [CrossRef]
- Zhang, X.; Li, C.; Chen, D.; He, X.; Zhao, Y.; Bao, L.; Wang, Q.; Zhou, J.; Xie, Y.H. pylori CagA activates the NLRP3 inflammasome to promote gastric cancer cell migration and invasion. Inflamm. Res. 2022, 71, 141–155. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Y.; Zhu, Y.; Li, N.; Li, W.; Shang, C.; Song, G.; Li, S.; Cong, J.; Li, T.; et al. Apoptin induces pyroptosis of colorectal cancer cells via the GSDME-dependent pathway. Int. J. Biol. Sci. 2022, 18, 717–730. [Google Scholar] [CrossRef]
- Wang, G.; Wei, W.; Jiang, Z.; Jiang, J.; Han, J.; Zhang, H.; Hu, J.; Zhang, P.; Li, X.; Chen, T.; et al. Talaromyces marneffei activates the AIM2-caspase-1/-4-GSDMD axis to induce pyroptosis in hepatocytes. Virulence 2022, 13, 963–979. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, T.; Tian, H.; Wu, J.; Yu, X.; Zeng, L.; Liu, F.; Liu, Q.; Huang, X. Coxsackievirus Group B3 Has Oncolytic Activity against Colon Cancer through Gasdermin E-Mediated Pyroptosis. Cancers 2022, 14, 6206. [Google Scholar] [CrossRef]
- Pizato, N.; Luzete, B.C.; Kiffer, L.; Correa, L.H.; de Oliveira Santos, I.; Assumpcao, J.A.F.; Ito, M.K.; Magalhaes, K.G. Omega-3 docosahexaenoic acid induces pyroptosis cell death in triple-negative breast cancer cells. Sci. Rep. 2018, 8, 1952. [Google Scholar] [CrossRef]
- Wang, X.; Yin, Y.; Qian, W.; Peng, C.; Shen, S.; Wang, T.; Zhao, S. Citric acid of ovarian cancer metabolite induces pyroptosis via the caspase-4/TXNIP-NLRP3-GSDMD pathway in ovarian cancer. FASEB J. 2022, 36, e22362. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Wang, J.; Xie, J.; Chen, L.; Wei, X.; Jiang, X.; Bao, M.; Qiu, Y.; Chen, Q.; Li, W.; et al. Design, synthesis, and evaluation of chalcone analogues incorporate alpha,beta-Unsaturated ketone functionality as anti-lung cancer agents via evoking ROS to induce pyroptosis. Eur. J. Med. Chem. 2018, 157, 1395–1405. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Zhou, B.; Sun, R.Y.; Ai, Y.L.; Cheng, K.; Li, F.N.; Wang, B.R.; Liu, F.J.; Jiang, Z.H.; Wang, W.J.; et al. The metabolite alpha-KG induces GSDMC-dependent pyroptosis through death receptor 6-activated caspase-8. Cell Res. 2021, 31, 980–997. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Reagents | Category | Mechanism | Cells | Reference |
|---|---|---|---|---|
| Topotecan | Topoisomerase 1 inhibitor | caspase-3/GSDM-E | SH-SY5Y, MeWo | [29] |
| Etoposide | Topoisomerase II inhibitor | caspase-3/GSDM-E | SH-SY5Y, MeWo | [29] |
| Cisplatin | Alkylating agent | caspase-3/GSDM-E | SH-SY5Y, MeWo, A549 | [37,40] |
| 5-Fu | Anti-metabolites | caspase-3/GSDM-E | SGC-7901 MKN-45 | [41] |
| Paclitaxel | Microtubules interference | caspase-3/GSDM-E | A549 | [41] |
| Ru (II) polypyridyl + Paclitaxel | caspase-1/GSDM-D | HeLa | [42] | |
| Lobaplatin | Platinum | caspase-3/GSDM-E | HT-29, HCT116 | [38] |
| Doxorubicin | Anthracycline | caspase-3/GSDM-E | HepG2, Hep 3B | [43] |
| FL118 | Camptothecin | _ | Colon cancer cells | [44] |
| Name | Target | Mechanisms | Cancer Types | Reference |
|---|---|---|---|---|
| BI2536 | PLK1 | caspase-3/GSDM-E | Esophageal | [59,60] |
| Alpha-NETA | Choline acetylcholine transferase | caspase-4/GSDM-D | Ovarian | [61] |
| BIX-01294 (BIX) | G9a | caspase-3/GSDM-E | Gastric | [62] |
| NO. 0449-0145 | APE1 | caspase-4/GSDM-D | Lung | [63] |
| Famotidine | Histamine H2-receptor | GSDM-E | gastric | [64] |
| Bexarotene | RXR | caspase-4/GSDM-E | ovarian | [65] |
| AT7519 | CDK | caspase-3/GSDM-E | Glioblastoma | [66] |
| Elraglusib | GSK-3 | GSDM-B | Colon | [67] |
| Name | Category | Mechanisms | Cancer Types | Reference |
|---|---|---|---|---|
| Curcumin | Polyphenol | caspase-3/GSDM-E | Liver | [75] |
| Anthocyanins | Flavonoid | caspase-1/GSDM-D | Oral | [76] |
| Miltirone | Abietane-type diterpenoids | caspase-3/GSDM-E | Liver | [77] |
| Tetraarsenic hexoxide | Arsenic oxide | caspase-3/GSDM-E | Breast | [78] |
| Triptolide | Diterpenoid epoxide | caspase-3/GSDM-E | Oral | [79] |
| Trichosanthin | Ribosome-inactivating protein | caspase-1/GSDM-D | Lung | [80] |
| Aloe-emodin | Anthraquinone | caspase-3/GSDM-E | Cervical | [81] |
| Myricetin | Flavonoid | caspase-3/GSDM-E | Lung | [82] |
| Luteolin | Flavonoid | caspase-1/GSDM-D | Colon | [83] |
| Mallotucin D | Flavonoid glycoside | caspase-3/GSDM-E | Liver | [84] |
| Shikonin | Naphthoquinone | caspase-3/GSDM-E | Gastric | [85] |
| Name | Category | Mechanisms | Cancer Types | Reference |
|---|---|---|---|---|
| 4-hydroxybenzoic acid | Benzoic acid derivatives | caspase-1/GSDM-D | Lung | [94] |
| EV-A71 | Enterovirus | caspase-1/GSDM-D | Neuroblastoma | [95] |
| CagA | Protein from Helicobacter pylori | caspase-1/GSDM-D | Gastric | [96] |
| Apoptin | Protein from chicken anemia virus | caspase-3/GSDM-E | Colon | [97] |
| Talaromyces marneffei | Fungus | caspase-1/GSDM-D | Liver | [98] |
| Coxsackievirus group B3 | Enterovirus | caspase-3/GSDM-E | Colon | [99] |
| Name | Category | Mechanisms | Cancer Types | Reference |
|---|---|---|---|---|
| Omega-3 docosahexaenoic acid | Polyunsaturated fatty acid | caspase-1/GSDM-D | Breast | [100] |
| Citric acid | Tricarboxylic acid | caspase-4/GSDM-D | Ovarian | [101] |
| α, β-Unsaturated ketone | Unsaturated carbonyl compound | caspase-3/GSDM-E | Lung cancer | [102] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, D.E.; Cui, Z. Triggering Pyroptosis in Cancer. Biomolecules 2025, 15, 348. https://doi.org/10.3390/biom15030348
Johnson DE, Cui Z. Triggering Pyroptosis in Cancer. Biomolecules. 2025; 15(3):348. https://doi.org/10.3390/biom15030348
Chicago/Turabian StyleJohnson, Daniel E., and Zhibin Cui. 2025. "Triggering Pyroptosis in Cancer" Biomolecules 15, no. 3: 348. https://doi.org/10.3390/biom15030348
APA StyleJohnson, D. E., & Cui, Z. (2025). Triggering Pyroptosis in Cancer. Biomolecules, 15(3), 348. https://doi.org/10.3390/biom15030348