Extracellular Vesicles in the Development of the Non-Alcoholic Fatty Liver Disease: An Update



Abstract
1. Introduction
2. Extracellular Vesicles (EVs): Biogenesis and Classes
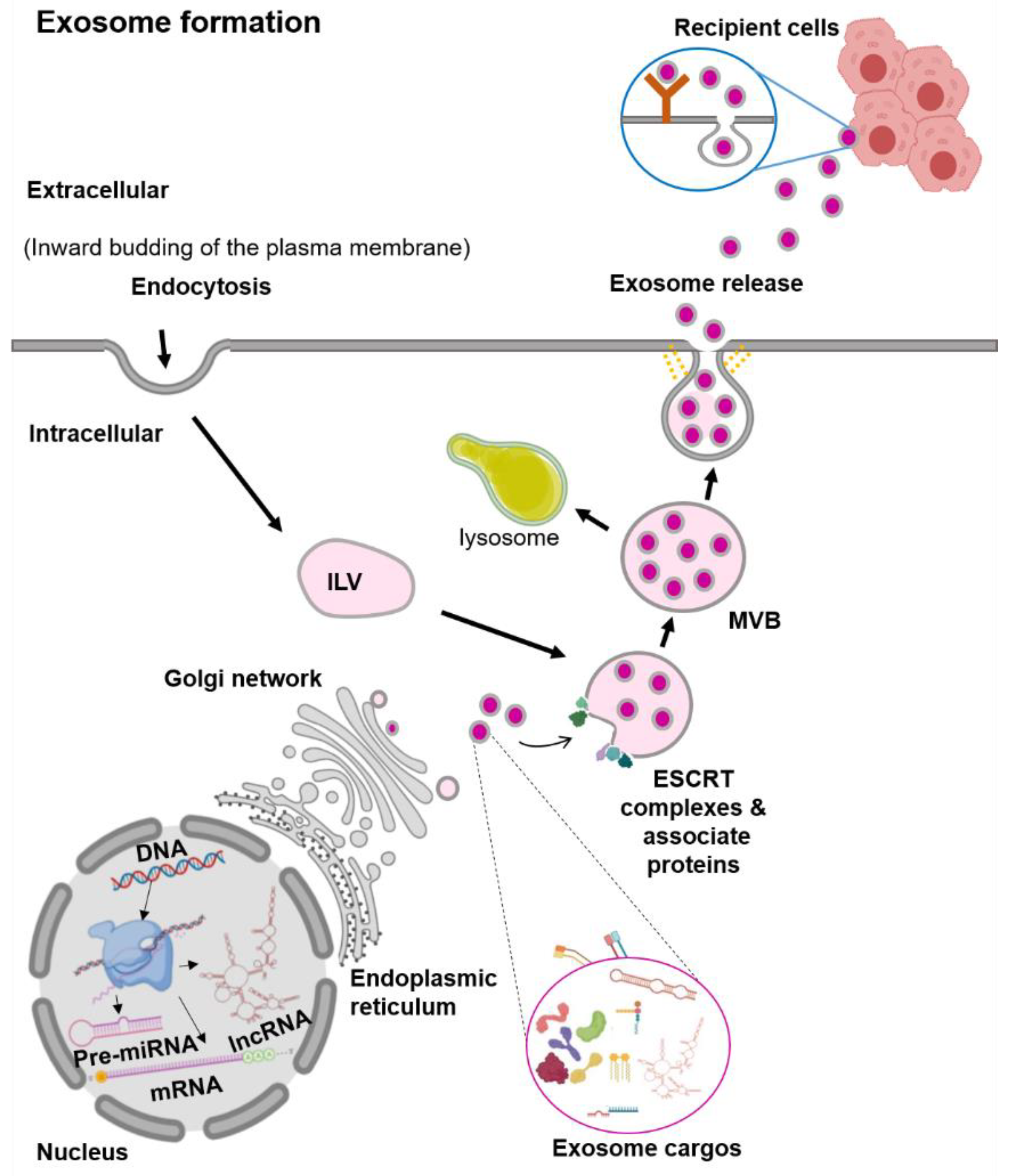
2.1. Extracellular Vesicles (EVs): Exosomes
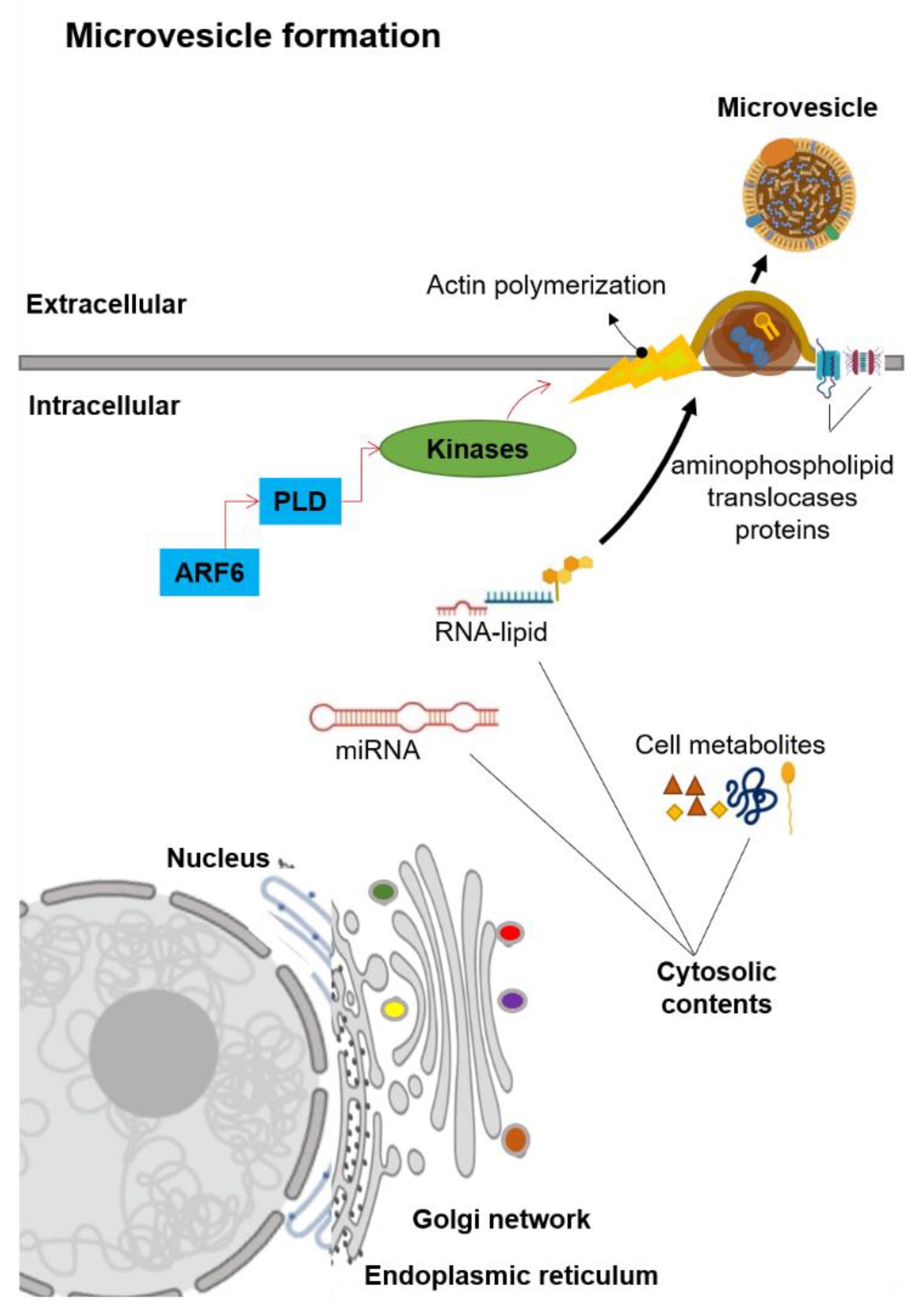
2.2. Extracellular Vesicles (EVs): Microvesicles
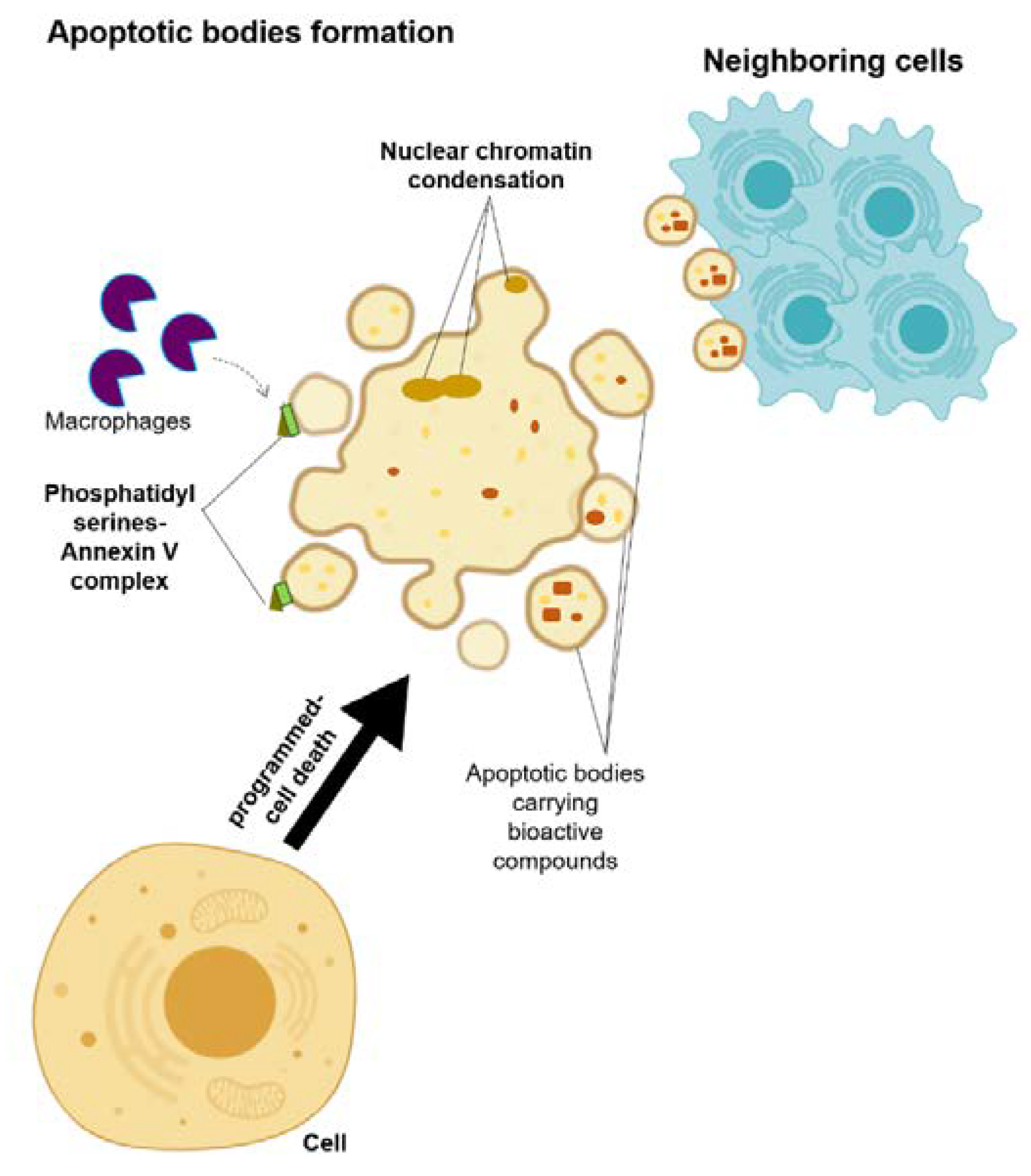
2.3. Extracellular Vesicles (EVs): Apoptotic Bodies
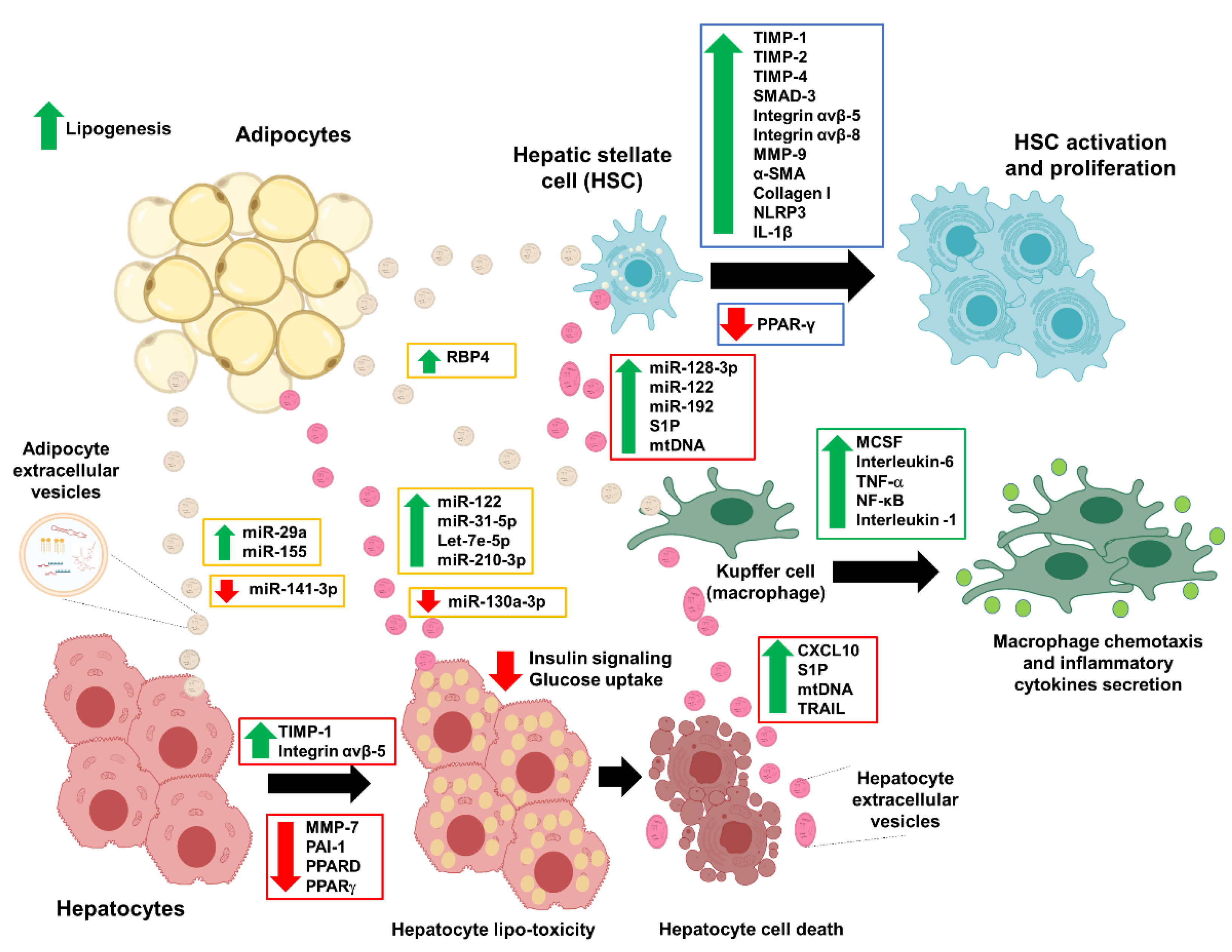
3. Extracellular Vesicles (EVs): Adipocyte-Derived EVs in NAFLD
4. Extracellular Vesicles (EVs): Damaged Hepatocytes Roles in NAFLD
5. EVs from the Mesenchymal Stem Cells as a Treatment Option
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of nafld and nash: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Araújo, A.R.; Rosso, N.; Bedogni, G.; Tiribelli, C.; Bellentani, S. Global epidemiology of non-alcoholic fatty liver disease/non-alcoholic steatohepatitis: What we need in the future. Liver Int. 2018, 38 (Suppl. 1), 47–51. [Google Scholar]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Cobbina, E.; Akhlaghi, F. Non-alcoholic fatty liver disease (nafld)-pathogenesis, classification, and effect on drug metabolizing enzymes and transporters. Drug Metab. Rev. 2017, 49, 197–211. [Google Scholar] [CrossRef] [PubMed]
- LaBrecque, D.R.; Abbas, Z.; Anania, F.; Ferenci, P.; Khan, A.G.; Goh, K.L.; Hamid, S.S.; Isakov, V.; Lizarzabal, M.; Peñaranda, M.M.; et al. World gastroenterology organisation global guidelines: Nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. J. Clin. Gastroenterol. 2014, 48, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Watt, M.J.; Miotto, P.M.; De Nardo, W.; Montgomery, M.K. The liver as an endocrine organ—Linking nafld and insulin resistance. Endocr. Rev. 2019, 40, 1367–1393. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, S.A.; Muhsin, N.I.A.; Jamal, R. Regulatory non-coding rnas network in non-alcoholic fatty liver disease. Front. Physiol. 2019, 10, 279. [Google Scholar] [CrossRef]
- Haas, J.T.; Francque, S.; Staels, B. Pathophysiology and mechanisms of nonalcoholic fatty liver disease. Annu. Rev. Physiol. 2016, 78, 181–205. [Google Scholar] [CrossRef]
- Li, C.-J.; Fang, Q.-H.; Liu, M.-L.; Lin, J.-N. Current understanding of the role of adipose-derived extracellular vesicles in metabolic homeostasis and diseases: Communication from the distance between cells/tissues. Theranostics 2020, 10, 7422–7435. [Google Scholar] [CrossRef]
- Castaño, C.; Kalko, S.; Novials, A.; Párrizas, M. Obesity-associated exosomal mirnas modulate glucose and lipid metabolism in mice. Proc. Natl. Acad. Sci. USA 2018, 115, 12158–12163. [Google Scholar] [CrossRef]
- Hirsova, P.; Ibrahim, S.H.; Krishnan, A.; Verma, V.K.; Bronk, S.F.; Werneburg, N.W.; Charlton, M.R.; Shah, V.H.; Malhi, H.; Gores, G.J. Lipid-induced signaling causes release of inflammatory extracellular vesicles from hepatocytes. Gastroenterology 2016, 150, 956–967. [Google Scholar] [PubMed]
- Eguchi, A.; Feldstein, A.E. Extracellular vesicles in non-alcoholic and alcoholic fatty liver diseases. Liver Res. 2018, 2, 30–34. [Google Scholar] [PubMed]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of extracellular vesicles (ev): Exosomes; microvesicles; retrovirus-like vesicles; and apoptotic bodies. J. Neurooncol. 2013, 113, 1–11. [Google Scholar] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology; function; and biomedical applications of exosomes. Science 2020, 367. [Google Scholar] [CrossRef]
- Chen, Y.; Li, G.; Liu, M.-L. Microvesicles as emerging biomarkers and therapeutic targets in cardiometabolic diseases. Genom. Proteom. Bioinform. 2018, 16, 50–62. [Google Scholar]
- Hon, K.W.; Ab-Mutalib, N.S.; Abdullah, N.M.A.; Jamal, R.; Abu, N. Extracellular vesicle–derived circular rnas confers chemoresistance in colorectal cancer. Sci. Rep. 2019, 9, 16497. [Google Scholar]
- Ab Razak, N.S.; Ab Mutalib, N.S.; Mohtar, M.A.; Abu, N. Impact of chemotherapy on extracellular vesicles: Understanding the chemo-evs. Front. Oncol. 2019, 9, 1113. [Google Scholar]
- Juan, T.; Fürthauer, M. Biogenesis and function of escrt-dependent extracellular vesicles. Semin. Cell Dev. Biol. 2018, 74, 66–77. [Google Scholar]
- McCullough, J.; Fisher, R.D.; Whitby, F.G.; Sundquist, W.I.; Hill, C.P. Alix-chmp4 interactions in the human escrt pathway. Proc. Natl. Acad. Sci. USA 2008, 105, 7687–7691. [Google Scholar]
- Babst, M. Mvb vesicle formation: Escrt-dependent; escrt-independent and everything in between. Curr. Opin. Cell Biol. 2011, 23, 452–457. [Google Scholar]
- Janas, T.; Janas, M.M.; Sapoń, K.; Janas, T. Mechanisms of rna loading into exosomes. FEBS Lett. 2015, 589, 1391–1398. [Google Scholar] [CrossRef] [PubMed]
- Phuyal, S.; Hessvik, N.P.; Skotland, T.; Sandvig, K.; Llorente, A. Regulation of exosome release by glycosphingolipids and flotillins. FEBS J. 2014, 281, 2214–2227. [Google Scholar] [CrossRef] [PubMed]
- Elsherbini, A.; Bieberich, E. Ceramide and exosomes: A novel target in cancer biology and therapy. Adv. Cancer Res. 2018, 140, 121–154. [Google Scholar] [PubMed]
- Kajimoto, T.; Okada, T.; Miya, S.; Zhang, L.; Nakamura, S. Ongoing activation of sphingosine 1-phosphate receptors mediates maturation of exosomal multivesicular endosomes. Nat. Commun. 2013, 4, 2712. [Google Scholar] [CrossRef] [PubMed]
- Stuffers, S.; Sem Wegner, C.; Stenmark, H.; Brech, A. Multivesicular endosome biogenesis in the absence of escrts. Traffic 2009, 10, 925–937. [Google Scholar] [CrossRef]
- Sebastian, T.T.; Baldridge, R.D.; Xu, P.; Graham, T.R. Phospholipid flippases: Building asymmetric membranes and transport vesicles. Biochim. Biophys. Acta 2012, 1821, 1068–1077. [Google Scholar] [CrossRef]
- Leventis, P.A.; Grinstein, S. The distribution and function of phosphatidylserine in cellular membranes. Annu. Rev. Biophys. 2010, 39, 407–427. [Google Scholar] [CrossRef]
- Panatala, R.; Hennrich, H.; Holthuis, J.C. Inner workings and biological impact of phospholipid flippases. J. Cell Sci. 2015, 128, 2021–2032. [Google Scholar] [CrossRef]
- Zhou, W.-B.; Gong, J.-S.; Hou, H.-J.; Li, H.; Lu, Z.-M.; Xu, H.-Y.; Xu, Z.-H.; Shi, J.-S. Mining of a phospholipase d and its application in enzymatic preparation of phosphatidylserine. Bioengineered 2018, 9, 80–89. [Google Scholar] [CrossRef]
- Van Acker, T.; Tavernier, J.; Peelman, F. The small gtpase arf6: An overview of its mechanisms of action and of its role in host-pathogen interactions and innate immunity. Int. J. Mol. Sci. 2019, 20, 2209. [Google Scholar] [CrossRef]
- Muralidharan-Chari, V.; Clancy, J.; Plou, C.; Romao, M.; Chavrier, P.; Raposo, G.; D’Souza-Schorey, C. Arf6-regulated shedding of tumor cell-derived plasma membrane microvesicles. Curr. Biol. 2009, 19, 1875–1885. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lai, Y.; Hua, Z.-C. Apoptosis and apoptotic body: Disease message and therapeutic target potentials. Biosci. Rep. 2019, 39, BSR20180992. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.C.; Freyssinet, J.M. Deciphering the plasma membrane hallmarks of apoptotic cells: Phosphatidylserine transverse redistribution and calcium entry. BMC Cell Biol. 2001, 2, 20. [Google Scholar]
- Deng, Z.B.; Poliakov, A.; Hardy, R.W.; Clements, R.; Liu, C.; Liu, Y.; Wang, J.; Xiang, X.; Zhang, S.; Zhuang, X.; et al. Adipose tissue exosome-like vesicles mediate activation of macrophage-induced insulin resistance. Diabetes 2009, 58, 2498–2505. [Google Scholar] [CrossRef] [PubMed]
- Jayabalan, N.; Lai, A.; Ormazabal, V.; Adam, S.; Guanzon, D.; Palma, C.; Scholz-Romero, K.; Lim, R.; Jansson, T.; McIntyre, H.D.; et al. Adipose tissue exosomal proteomic profile reveals a role on placenta glucose metabolism in gestational diabetes mellitus. J. Clin. Endocrinol. Metab. 2019, 104, 1735–1752. [Google Scholar] [CrossRef]
- Ferrante, S.C.; Nadler, E.P.; Pillai, D.K.; Hubal, M.J.; Wang, Z.; Wang, J.M.; Gordish-Dressman, H.; Koeck, E.; Sevilla, S.; Wiles, A.A.; et al. Adipocyte-derived exosomal mirnas: A novel mechanism for obesity-related disease. Pediatr. Res. 2015, 77, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Koeck, E.S.; Iordanskaia, T.; Sevilla, S.; Ferrante, S.C.; Hubal, M.J.; Freishtat, R.J.; Nadler, E.P. Adipocyte exosomes induce transforming growth factor beta pathway dysregulation in hepatocytes: A novel paradigm for obesity-related liver disease. J. Surg. Res. 2014, 192, 268–275. [Google Scholar]
- Ying, W.; Riopel, M.; Bandyopadhyay, G.; Dong, Y.; Birmingham, A.; Seo, J.B.; Ofrecio, J.M.; Wollam, J.; Hernandez-Carretero, A.; Fu, W.; et al. Adipose tissue macrophage-derived exosomal mirnas can modulate in vivo and in vitro insulin sensitivity. Cell 2017, 171, 372–384. [Google Scholar] [CrossRef]
- Liu, T.; Sun, Y.C.; Cheng, P.; Shao, H.G. Adipose tissue macrophage-derived exosomal mir-29a regulates obesity-associated insulin resistance. Biochem. Biophys. Res. Commun. 2019, 515, 352–358. [Google Scholar] [CrossRef]
- Dang, S.Y.; Leng, Y.; Wang, Z.X.; Xiao, X.; Zhang, X.; Wen, T.; Gong, H.Z.; Hong, A.; Ma, Y. Exosomal transfer of obesity adipose tissue for decreased mir-141-3p mediate insulin resistance of hepatocytes. Int. J. Biol. Sci. 2019, 15, 351–368. [Google Scholar] [CrossRef]
- Hubal, M.J.; Nadler, E.P.; Ferrante, S.C.; Barberio, M.D.; Suh, J.H.; Wang, J.; Dohm, G.L.; Pories, W.J.; Mietus-Snyder, M.; Freishtat, R.J. Circulating adipocyte-derived exosomal micrornas associated with decreased insulin resistance after gastric bypass. Obesity (Silver Spring) 2017, 25, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Kranendonk, M.E.; de Kleijn, D.P.; Kalkhoven, E.; Kanhai, D.A.; Uiterwaal, C.S.; van der Graaf, Y.; Pasterkamp, G.; Visseren, F.L. Extracellular vesicle markers in relation to obesity and metabolic complications in patients with manifest cardiovascular disease. Cardiovasc. Diabetol. 2014, 13, 37. [Google Scholar] [CrossRef] [PubMed]
- de Mendonça, M.; Rocha, K.C.; de Sousa, É.; Pereira, B.M.V.; Oyama, L.M.; Rodrigues, A.C. Aerobic exercise training regulates serum extracellular vesicle mirnas linked to obesity to promote their beneficial effects in mice. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E579–E591. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, A.; Mulya, A.; Lazic, M.; Radhakrishnan, D.; Berk, M.P.; Povero, D.; Gornicka, A.; Feldstein, A.E. Microparticles release by adipocytes act as “find-me” signals to promote macrophage migration. PLoS ONE 2015, 10, e0123110. [Google Scholar] [CrossRef]
- Li, J.; Liu, H.; Mauer, A.S.; Lucien, F.; Raiter, A.; Bandla, H.; Mounajjed, T.; Yin, Z.; Glaser, K.J.; Yin, M.; et al. Characterization of cellular sources and circulating levels of extracellular vesicles in a dietary murine model of nonalcoholic steatohepatitis. Hepatol. Commun. 2019, 3, 1235–1249. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Salomon, C.; Freeman, D.J. Extracellular vesicles from adipose tissue-a potential role in obesity and type 2 diabetes? Front. Endocrinol. 2017, 8, 202. [Google Scholar] [CrossRef] [PubMed]
- DeClercq, V.; d’Eon, B.; McLeod, R.S. Fatty acids increase adiponectin secretion through both classical and exosome pathways. Biochim. Biophys. Acta 2015, 1851, 1123–1133. [Google Scholar] [CrossRef]
- Liao, C.Y.; Song, M.J.; Gao, Y.; Mauer, A.S.; Revzin, A.; Malhi, H. Hepatocyte-derived lipotoxic extracellular vesicle sphingosine 1-phosphate induces macrophage chemotaxis. Front. Immunol. 2018, 9, 2980. [Google Scholar] [CrossRef]
- Tosello-Trampont, A.C.; Landes, S.G.; Nguyen, V.; Novobrantseva, T.I.; Hahn, Y.S. Kuppfer cells trigger nonalcoholic steatohepatitis development in diet-induced mouse model through tumor necrosis factor-α production. J. Biol. Chem. 2012, 287, 40161–40172. [Google Scholar] [CrossRef]
- Wu, J.; Dong, T.; Chen, T.; Sun, J.; Luo, J.; He, J.; Wei, L.; Zeng, B.; Zhang, H.; Li, W.; et al. Hepatic exosome-derived mir-130a-3p attenuates glucose intolerance via suppressing phlpp2 gene in adipocyte. Metabolism 2020, 103, 154006. [Google Scholar] [CrossRef]
- Lee, Y.S.; Kim, S.Y.; Ko, E.; Lee, J.H.; Yi, H.S.; Yoo, Y.J.; Je, J.; Suh, S.J.; Jung, Y.K.; Kim, J.H.; et al. Exosomes derived from palmitic acid-treated hepatocytes induce fibrotic activation of hepatic stellate cells. Sci. Rep. 2017, 7, 3710. [Google Scholar] [CrossRef] [PubMed]
- Cannito, S.; Morello, E.; Bocca, C.; Foglia, B.; Benetti, E.; Novo, E.; Chiazza, F.; Rogazzo, M.; Fantozzi, R.; Povero, D. Microvesicles released from fat-laden cells promote activation of hepatocellular nlrp3 inflammasome: A pro-inflammatory link between lipotoxicity and non-alcoholic steatohepatitis. PLoS ONE 2017, 12, e0172575. [Google Scholar] [CrossRef]
- Povero, D.; Panera, N.; Eguchi, A.; Johnson, C.D.; Papouchado, B.G.; de Araujo Horcel, L.; Pinatel, E.M.; Alisi, A.; Nobili, V.; Feldstein, A.E. Lipid-induced hepatocyte-derived extracellular vesicles regulate hepatic stellate cell via micrornas targeting ppar-γ. Cell Mol. Gastroenterol. Hepatol. 2015, 1, 646–663.e4. [Google Scholar] [CrossRef]
- Ibrahim, S.H.; Hirsova, P.; Tomita, K.; Bronk, S.F.; Werneburg, N.W.; Harrison, S.A.; Goodfellow, V.S.; Malhi, H.; Gores, G.J. Mixed lineage kinase 3 mediates release of c-x-c motif ligand 10-bearing chemotactic extracellular vesicles from lipotoxic hepatocytes. Hepatology 2016, 63, 731–744. [Google Scholar] [CrossRef]
- Mauer, A.S.; Hirsova, P.; Maiers, J.L.; Shah, V.H.; Malhi, H. Inhibition of sphingosine 1-phosphate signaling ameliorates murine nonalcoholic steatohepatitis. Am. J. Physiol. Gastrointest Liver Physiol. 2017, 312, G300–G313. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, M.F.; Jiang, S.; Wu, J.; Liu, J.; Yuan, X.W.; Shen, D.; Zhang, J.Z.; Zhou, N.; He, J.; et al. Liver governs adipose remodelling via extracellular vesicles in response to lipid overload. Nat. Commun. 2020, 11, 719. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.; Reyes, D.; Geng, Y.; Arab, J.P.; Cabrera, D.; Sepulveda, R.; Solis, N.; Buist-Homan, M.; Arrese, M.; Moshage, H. Extracellular vesicles derived from fat-laden hepatocytes undergoing chemical hypoxia promote a pro-fibrotic phenotype in hepatic stellate cells. Biochim. Biophys. Acta Mol. Basis. Dis. 2020, 1866, 165857. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.; Geng, Y.; Sepúlveda, R.; Solís, N.; Torres, J.; Arab, J.P.; Barrera, F.; Cabrera, D.; Moshage, H.; Arrese, M. Chemical hypoxia induces pro-inflammatory signals in fat-laden hepatocytes and contributes to cellular crosstalk with kupffer cells through extracellular vesicles. Biochim. Biophys. Acta Mol. Basis. Dis. 2020, 1866, 165753. [Google Scholar] [CrossRef]
- Jiang, F.; Chen, Q.; Wang, W.; Ling, Y.; Yan, Y.; Xia, P. Hepatocyte-derived extracellular vesicles promote endothelial inflammation and atherogenesis via microrna-1. J. Hepatol. 2020, 72, 156–166. [Google Scholar] [CrossRef]
- Szabo, G.; Momen-Heravi, F. Extracellular vesicles in liver disease and potential as biomarkers and therapeutic targets. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 455–466. [Google Scholar] [CrossRef]
- Kakazu, E.; Mauer, A.S.; Yin, M.; Malhi, H. Hepatocytes release ceramide-enriched pro-inflammatory extracellular vesicles in an ire1α-dependent manner. J. Lipid. Res. 2016, 57, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Martinez, I.; Santoro, N.; Chen, Y.; Hoque, R.; Ouyang, X.; Caprio, S.; Shlomchik, MJ.; Coffman, RL.; Candia, A.; Mehal, W.Z. Hepatocyte mitochondrial DNA drives nonalcoholic steatohepatitis by activation of tlr9. J. Clin. Investig. 2016, 126, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Sasai, M.; Linehan, M.M.; Iwasaki, A. Bifurcation of toll-like receptor 9 signaling by adaptor protein 3. Science 2010, 329, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Mardpour, S.; Ghanian, M.H.; Sadeghi-Abandansari, H.; Mardpour, S.; Nazari, A.; Shekari, F.; Baharvand, H. Hydrogel-mediated sustained systemic delivery of mesenchymal stem cell-derived extracellular vesicles improves hepatic regeneration in chronic liver failure. ACS Appl. Mater. Interfaces 2019, 11, 37421–37433. [Google Scholar] [CrossRef]
- Povero, D.; Eguchi, A.; Niesman, I.R.; Andronikou, N.; de Mollerat du Jeu, X.; Mulya, A.; Berk, M.; Lazic, M.; Thapaliya, S.; Parola, M.; et al. Lipid-induced toxicity stimulates hepatocytes to release angiogenic microparticles that require vanin-1 for uptake by endothelial cells. Sci. Signal. 2013, 6, ra88. [Google Scholar] [CrossRef]
- Baglio, S.R.; Rooijers, K.; Koppers-Lalic, D.; Verweij, F.J.; Pérez Lanzón, M.; Zini, N.; Naaijkens, B.; Perut, F.; Niessen, H.W.; Baldini, N.; et al. Human bone marrow- and adipose-mesenchymal stem cells secrete exosomes enriched in distinctive mirna trna species. Stem Cell Res. 2015, 6, 127. [Google Scholar] [CrossRef]
- Kalinina, N.; Kharlampieva, D.; Loguinova, M.; Butenko, I.; Pobeguts, O.; Efimenko, A.; Ageeva, L.; Sharonov, G.; Ischenko, D.; Alekseev, D.; et al. Characterization of secretomes provides evidence for adipose-derived mesenchymal stromal cells subtypes. Stem Cell Res. 2015, 6, 221. [Google Scholar] [CrossRef]
- Togliatto, G.; Dentelli, P.; Gili, M.; Gallo, S.; Deregibus, C.; Biglieri, E.; Iavello, A.; Santini, E.; Rossi, C.; Solini, A.; et al. Obesity reduces the pro-angiogenic potential of adipose tissue stem cell-derived extracellular vesicles (evs) by impairing mir-126 content: Impact on clinical applications. Int. J. Obes. 2016, 40, 102–111. [Google Scholar] [CrossRef]
- Lopatina, T.; Bruno, S.; Tetta, C.; Kalinina, N.; Porta, M.; Camussi, G. Platelet-derived growth factor regulates the secretion of extracellular vesicles by adipose mesenchymal stem cells and enhances their angiogenic potential. Cell Commun. Signal. 2014, 12, 26. [Google Scholar] [CrossRef]
- Liu, R.; Shen, H.; Ma, J.; Sun, L.; Wei, M. Extracellular vesicles derived from adipose mesenchymal stem cells regulate the phenotype of smooth muscle cells to limit intimal hyperplasia. Cardiovasc. Drugs 2016, 30, 111–118. [Google Scholar] [CrossRef]
- Eirin, A.; Zhu, X.-Y.; Puranik, A.S.; Tang, H.; McGurren, K.A.; van Wijnen, A.J.; Lerman, A.; Lerman, L.O. Mesenchymal stem cell-derived extracellular vesicles attenuate kidney inflammation. Kidney Int. 2017, 92, 114–124. [Google Scholar] [CrossRef]
- Zhao, H.; Shang, Q.; Pan, Z.; Bai, Y.; Li, Z.; Zhang, H.; Zhang, Q.; Guo, C.; Zhang, L.; Wang, Q. Exosomes from adipose-derived stem cells attenuate adipose inflammation and obesity through polarizing m2 macrophages and beiging in white adipose tissue. Diabetes 2018, 67, 235–247. [Google Scholar] [CrossRef]
- Li, T.; Yan, Y.; Wang, B.; Qian, H.; Zhang, X.; Shen, L.; Wang, M.; Zhou, Y.; Zhu, W.; Li, W.; et al. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate liver fibrosis. Stem Cells Dev. 2013, 22, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.Y.; Lai, R.C.; Wong, W.; Dan, Y.Y.; Lim, S.K.; Ho, H.K. Mesenchymal stem cell-derived exosomes promote hepatic regeneration in drug-induced liver injury models. Stem Cell Res. 2014, 5, 76. [Google Scholar] [CrossRef] [PubMed]
- Haga, H.; Yan, I.K.; Takahashi, K.; Matsuda, A.; Patel, T. Extracellular vesicles from bone marrow-derived mesenchymal stem cells improve survival from lethal hepatic failure in mice. Stem Cells Transl. Med. 2017, 6, 1262–1272. [Google Scholar] [CrossRef]
- Damania, A.; Jaiman, D.; Teotia, A.K.; Kumar, A. Mesenchymal stromal cell-derived exosome-rich fractionated secretome confers a hepatoprotective effect in liver injury. Stem Cell Res. 2018, 9, 31. [Google Scholar] [CrossRef]
- Haga, H.; Yan, I.K.; Borrelli, D.A.; Matsuda, A.; Parasramka, M.; Shukla, N.; Lee, D.D.; Patel, T. Extracellular vesicles from bone marrow-derived mesenchymal stem cells protect against murine hepatic ischemia/reperfusion injury. Liver Transpl. 2017, 23, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Anger, F.; Camara, M.; Ellinger, E.; Germer, C.T.; Schlegel, N.; Otto, C.; Klein, I. Human mesenchymal stromal cell-derived extracellular vesicles improve liver regeneration after ischemia reperfusion injury in mice. Stem Cells Dev. 2019, 28, 1451–1462. [Google Scholar] [CrossRef]
- Rong, X.; Liu, J.; Yao, X.; Jiang, T.; Wang, Y.; Xie, F. Human bone marrow mesenchymal stem cells-derived exosomes alleviate liver fibrosis through the wnt/β-catenin pathway. Stem Cell Res. 2019, 10, 98. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, Y.; Pu, Z. Bone marrow mesenchymal stem cell-derived exosomes attenuate d-gain/lps-induced hepatocyte apoptosis by activating autophagy in vitro. Drug Des. Dev. Ther. 2019, 13, 2887–2897. [Google Scholar] [CrossRef]
- Yan, Y.; Jiang, W.; Tan, Y.; Zou, S.; Zhang, H.; Mao, F.; Gong, A.; Qian, H.; Xu, W. Hucmsc exosome-derived gpx1 is required for the recovery of hepatic oxidant injury. Mol Ther. 2017, 25, 465–479. [Google Scholar] [CrossRef]
- Yao, J.; Zheng, J.; Cai, J.; Zeng, K.; Zhou, C.; Zhang, J.; Li, S.; Li, H.; Chen, L.; He, L.; et al. Extracellular vesicles derived from human umbilical cord mesenchymal stem cells alleviate rat hepatic ischemia-reperfusion injury by suppressing oxidative stress and neutrophil inflammatory response. FASEB J. 2019, 33, 1695–1710. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, S.; Hu, H.; Yang, J.; Wang, X.; Ma, Y.; Jiang, J.; Wang, J.; Zhong, L.; Chen, M.; et al. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate acute liver failure by reducing the activity of the nlrp3 inflammasome in macrophages. Biochem. Biophys. Res. Commun. 2019, 508, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Herrera, M.B.; Fonsato, V.; Gatti, S.; Deregibus, M.C.; Sordi, A.; Cantarella, D.; Calogero, R.; Bussolati, B.; Tetta, C.; Camussi, G. Human liver stem cell-derived microvesicles accelerate hepatic regeneration in hepatectomized rats. J. Cell Mol. Med. 2010, 14, 1605–1618. [Google Scholar] [CrossRef]
- Herrera Sanchez, M.B.; Previdi, S.; Bruno, S.; Fonsato, V.; Deregibus, M.C.; Kholia, S.; Petrillo, S.; Tolosano, E.; Critelli, R.; Spada, M.; et al. Extracellular vesicles from human liver stem cells restore argininosuccinate synthase deficiency. Stem Cell Res. 2017, 8, 176. [Google Scholar] [CrossRef]
- Du, Y.; Li, D.; Han, C.; Wu, H.; Xu, L.; Zhang, M.; Zhang, J.; Chen, X. Exosomes from human-induced pluripotent stem cell-derived mesenchymal stromal cells (hipsc-mscs) protect liver against hepatic ischemia/reperfusion injury via activating sphingosine kinase and sphingosine-1-phosphate signaling pathway. Cell Physiol. Biochem. 2017, 43, 611–625. [Google Scholar] [CrossRef]
- Chen, L.; Xiang, B.; Wang, X.; Xiang, C. Exosomes derived from human menstrual blood-derived stem cells alleviate fulminant hepatic failure. Stem Cell Res. 2017, 8, 9. [Google Scholar] [CrossRef]
- Lee, S.C.; Kim, K.H.; Kim, O.H.; Lee, S.K.; Hong, H.E.; Won, S.S.; Jeon, S.J.; Choi, B.J.; Jeong, W.; Kim, S.J. Determination of optimized oxygen partial pressure to maximize the liver regenerative potential of the secretome obtained from adipose-derived stem cells. Stem Cell Res. 2017, 8, 181. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lou, G.; Li, A.; Zhang, T.; Qi, J.; Ye, D.; Zheng, M.; Chen, Z. Amsc-derived exosomes alleviate lipopolysaccharide/d-galactosamine-induced acute liver failure by mir-17-mediated reduction of txnip/nlrp3 inflammasome activation in macrophages. EBioMedicine 2018, 36, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Wang, J.; Li, H.; Gao, S.; Shi, R.; Yang, D.; Wang, X.; Wang, X.; Zhu, L.; Wang, X.; et al. Extracellular vesicles secreted by human adipose-derived stem cells (hascs) improve survival rate of rats with acute liver failure by releasing lncrna h19. EBioMedicine 2018, 34, 231–242. [Google Scholar] [CrossRef]
- Liau, L.L.; Makpol, S.; Azurah, A.G.N.; Chua, K.H. Human adipose-derived mesenchymal stem cells promote recovery of injured hepg2 cell line and show sign of early hepatogenic differentiation. Cytotechnology 2018, 70, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.; Wang, S.; Kim, J.; Kim, G.J.; Jung, Y. Microrna125b-mediated hedgehog signaling influences liver regeneration by chorionic plate-derived mesenchymal stem cells. Sci. Rep. 2015, 5, 14135. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Zhang, Q.; Cai, X.; Li, F.; Ma, Z.; Xu, M.; Lu, L. Exosomes derived from mir-181-5p-modified adipose-derived mesenchymal stem cells prevent liver fibrosis via autophagy activation. J. Cell Mol. Med. 2017, 21, 2491–2502. [Google Scholar] [CrossRef]
- Lou, G.; Yang, Y.; Liu, F.; Ye, B.; Chen, Z.; Zheng, M.; Liu, Y. Mir-122 modification enhances the therapeutic efficacy of adipose tissue-derived mesenchymal stem cells against liver fibrosis. J. Cell Mol. Med. 2017, 21, 2963–2973. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Tsuchiya, A.; Takeuchi, S.; Nojiri, S.; Yoshida, T.; Ogawa, M.; Itoh, M.; Takamura, M.; Suganami, T.; Ogawa, Y.; et al. Development of a non-alcoholic steatohepatitis model with rapid accumulation of fibrosis; and its treatment using mesenchymal stem cells and their small extracellular vesicles. Regen 2020, 14, 252–261. [Google Scholar] [CrossRef]
- Povero, D.; Pinatel, E.M.; Leszczynska, A.; Goyal, N.P.; Nishio, T.; Kim, J.; Kneiber, D.; de Araujo Horcel, L.; Eguchi, A.; Ordonez, P.M.; et al. Human induced pluripotent stem cell-derived extracellular vesicles reduce hepatic stellate cell activation and liver fibrosis. JCI Insight 2019, 5, e125652. [Google Scholar] [CrossRef]
- Ohara, M.; Ohnishi, S.; Hosono, H.; Yamamoto, K.; Yuyama, K.; Nakamura, H.; Fu, Q.; Maehara, O.; Suda, G.; Sakamoto, N. Extracellular vesicles from amnion-derived mesenchymal stem cells ameliorate hepatic inflammation and fibrosis in rats. Stem Cells Int. 2018, 2018, 3212643. [Google Scholar] [CrossRef]
- Bruno, S.; Pasquino, C.; Herrera Sanchez, M.B.; Tapparo, M.; Figliolini, F.; Grange, C.; Chiabotto, G.; Cedrino, M.; Deregibus, M.C.; Tetta, C.; et al. HLSC-derived extracellular vesicles attenuate liver fibrosis and inflammation in a murine model of non-alcoholic steatohepatitis. Mol. Ther. 2020, 28, 479–489. [Google Scholar] [CrossRef]
- Dong, L.; Pu, Y.; Chen, X.; Qi, X.; Zhang, L.; Xu, L.; Li, W.; Ma, Y.; Zhou, S.; Zhu, J. Hucmsc-extracellular vesicles downregulated hepatic stellate cell activation and reduced liver injury in S. japonicum-infected mice. Stem Cell Res. 2020, 11, 21. [Google Scholar]
- Zhang, Y.; Cai, W.; Huang, Q.; Gu, Y.; Shi, Y.; Huang, J.; Zhao, F.; Liu, Q.; Wei, X.; Jin, M. Mesenchymal stem cells alleviate bacteria-induced liver injury in mice by inducing regulatory dendritic cells. Hepatology 2014, 59, 671–682. [Google Scholar] [CrossRef]
- Quintanilha, L.F.; Takami, T.; Hirose, Y.; Fujisawa, K.; Murata, Y.; Yamamoto, N.; Goldenberg, R.C.; Terai, S.; Sakaida, I. Canine mesenchymal stem cells show antioxidant properties against thioacetamide-induced liver injury in vitro and in vivo. Hepatol. Res. 2014, 44, E206–E217. [Google Scholar] [PubMed]
- Fu, J.; Zhang, H.; Zhuang, Y.; Liu, H.; Shi, Q.; Li, D.; Ju, X. The role of n-acetyltransferase 8 in mesenchymal stem cell-based therapy for liver ischemia/reperfusion injury in rats. PLoS ONE 2014, 9, e103355. [Google Scholar]
- Zhang, J.; Zhou, S.; Zhou, Y.; Feng, F.; Wang, Q.; Zhu, X.; Ai, H.; Huang, X.; Zhang, X. Hepatocyte growth factor gene-modified adipose-derived mesenchymal stem cells ameliorate radiation induced liver damage in a rat model. PLoS ONE 2014, 9, e114670. [Google Scholar]
- van Poll, D.; Parekkadan, B.; Cho, C.H.; Berthiaume, F.; Nahmias, Y.; Tilles, A.W.; Yarmush, M.L. Mesenchymal stem cell-derived molecules directly modulate hepatocellular death and regeneration in vitro and in vivo. Hepatology 2008, 47, 1634–1643. [Google Scholar]
- Driscoll, J.; Patel, T. The mesenchymal stem cell secretome as an acellular regenerative therapy for liver disease. J. Gastroenterol. 2019, 54, 763–773. [Google Scholar]
- Hernández, A.; Arab, J.P.; Reyes, D.; Lapitz, A.; Moshage, H.; Bañales, J.M.; Arrese, M. Extracellular vesicles in nafld/ald: From pathobiology to therapy. Cells 2020, 9, 817. [Google Scholar]
- Zhao, L.; Chen, S.; Shi, X.; Cao, H.; Li, L. A pooled analysis of mesenchymal stem cell-based therapy for liver disease. Stem Cell Res. Ther. 2018, 9, 72. [Google Scholar]
- Tsuchiya, A.; Takeuchi, S.; Watanabe, T.; Yoshida, T.; Nojiri, S.; Ogawa, M.; Terai, S. Mesenchymal stem cell therapies for liver cirrhosis: Mscs as “conducting cells” for improvement of liver fibrosis and regeneration. Inflamm. Regen. 2019, 39, 18. [Google Scholar]
- Hassan, M.N.F.B.; Yazid, M.D.; Yunus, M.H.M.; Chowdhury, S.R.; Lokanathan, Y.; Idrus, R.B.H.; Ng, A.M.H.; Law, J.X. Large-scale expansion of human mesenchymal stem cells. Stem Cells Int. 2020, 2020, 9529465. [Google Scholar]
- Zhang, S.; Hou, Y.; Yang, J.; Xie, D.; Jiang, L.; Hu, H.; Hu, J.; Luo, C.; Zhang, Q. Application of mesenchymal stem cell exosomes and their drug-loading systems in acute liver failure. J. Cell Mol. Med. 2020, 24, 7082–7093. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracellular Vesicles | Cell Source | EVs-Derived Disease Model | Molecular Mediators in the EVs Cargos | Recipient Targets | Interaction | NAFLD Relevance | Reference |
|---|---|---|---|---|---|---|---|
| Exosome | Visceral adipose tissue (VAT) | Leptin-deficient (ob/ob) B6 mice, B6 mice fed high-fat diets | RBP4 | Bone marrow-derived macrophages (BMDM) | Increased production of MCSF, IL-6, and TNF-α | Activation of BMDM macrophages induced insulin resistance | [34] |
| Exosome | VAT | Human, Females with BMI > 30 kg/m2 | MicroRNAs | TGF-β and Wnt/β-catenin signaling | TGF-β signaling and Wnt/β-catenin signaling among the top significant pathways | MicroRNAs in the exosomes derived from the obese visceral adipocytes are predicted to regulate inflammatory and fibrotic signaling pathways | [36] |
| Exosomes | VAT | Human, Females with BMI 35–46 (obese) | - | Hepatocytes and Hepatic stellate cells (HSCs) | Induced the expressions of TIMP-1, TIMP-4, SMAD-3, MMP-9, integrins ανβ-5 and -8 | Dysfunctional ECM regulation in the liver cells due to obese adipocyte exosomes | [37] |
| Exosome | Adipose tissue macrophages (ATM) | C57BL6 mice fed high-fat diets (in vivo), 3T3-L1 adipocytes (in vitro) | MicroRNAs (specifically miR-155) | L6 muscle cells and primary hepatocytes | Enriched miR-155 in the obese ATM-derived exosomes suppressed the expression of its target gene, PPARγ, and the downstream pathways | MicroRNAs cargos of secreted ATM-derived exosomes induced insulin resistance and glucose intolerance | [38] |
| Exosome | ATM | C57/BL6 mice fed high-fat diets | MicroRNAs (specifically miR-29a) | PPARD | MiR-29a interacts with PPARD to promote obesity-induced insulin resistance | ATM-derived exosomal miR-29a impairs insulin sensitivity in vitro and in vivo | [39] |
| Exosome | Adipose tissue | C57BL/6J (B6) mice fed high-fat diets and B6 ob/ob mice | miR-141-3p | AML12 liver cells | Decreased miR-141-3p expression caused impaired insulin signaling and glucose uptake in the hepatocytes | Exosomes from obese adipose tissues induced hepatocyte insulin resistance | [40] |
| Exosomes | Adipocytes | Human, Females with BMI 51.2±8.8 kg/m2 | MicroRNAs | Insulin receptor signaling pathway | Circulating adipocyte-derived exosomes are modified following gastric bypass surgery and correlated with improved post-surgery insulin sensitivity | Bypass surgery intervention changed the properties of the exosomes derived from the adipocyte tissues | [41] |
| Exosomes | Hepatocytes | C57BL/6 mice fed high-fat diets | Sphingosine-1-phosphate (S1P) | BMDM | Hepatocytes EVs with S1P-enriched activated macrophage chemotaxis via the S1P1 receptor | Lipotoxic hepatocytes-derived EVs induce macrophage chemotaxis | [48] |
| Exosomes | Hepatocytes | C56Bl/6J mice fed high-fat diets | Pro-inflammatory lipids (C16:0 ceramide) | Macrophages | Lipotoxic hepatocyte-EVs stimulated macrophage chemotaxis via S1P generation | Lipotoxic hepatocytes-derived EVs induce macrophage chemotaxis | [49] |
| Exosomes | Hepatocytes | C56Bl/6J mice fed high-fat diets | miR-130a-3p | Adipocytes, PHLPP2 | High expression of miR-130a-3p suppressed PHLPP2 expression to activate AKT-AS160–GLUT4 signaling pathway in adipocytes | miR-130a-3p regulates glucose metabolism by increasing glucose uptake | [50] |
| Exosomes | Hepatocytes | Huh7 cells treated with palmitate | MicroRNAs (especially miR-122 and miR-192) | HSCs | Hepatocyte-EVs increased the expression of pro-fibrotic markers such as α-SMA, TGF-β, and COL1A1 in HSCs. | Activation of fibrosis molecules | [51] |
| Microvesicle | Hepatocytes | HepG2 cells treated with palmitate | - | HSCs and hepatocytes | Lipotoxic hepatocyte-microvesicle internalization activated NLRP3 inflammasome via NF-kB, pro-caspase-1 and pro-interleukin-1, IL-1β | Activation of inflammatory phenotype in macrophages | [52] |
| Extracellular vesicles | Adipocytes | Patients with vascular disease | Cystatin-C | Monocytes, endothelial cells, platelets | The elevated level of EVs-cystatin C associated with metabolic complications of obesity | Low HDL cholesterol was significantly related to higher EV-cystatin C levels | [42] |
| Extracellular vesicles | Hepatocytes | C57BL/6 mice with choline-deficient amino acid diet | MicroRNAs (especially miR-128-3p) | HSCs | miR-128-3p suppressed the expression of PPARγ in HSCs | Activation of the HSCs | [53] |
| Extracellular vesicles | Hepatocytes | C57BL/6 mice model of NASH | TRAIL | IL-1β and IL6 in BMDM | Lipotoxic hepatocytes induced releases of pro-inflammatory EVs that activated macrophage via the death receptor 5 (DR5)-dependent manner | Activation of inflammatory phenotype in macrophages due to excess lipids in the liver cells | [11] |
| Extracellular vesicles | Hepatocytes | Primary hepatocytes and Huh7 cells treated with palmitate | CXCL10 | BMDM | Lipotoxic EVs have enriched of CXCL10, a chemotaxis inducer for macrophages | Lipotoxic hepatocytes-EVs activated macrophage chemotaxis | [54] |
| Extracellular vesicles | Hepatocytes, macrophage, neutrophil, platelet | C56BL/6J mice fed high-fat diets | - | Changes in liver condition (onset of NASH) | Quantitative evolution of hepatocyte-, macrophage- and neutrophil-derived EVs correlated well with the histology of NASH | Circulating EVs derived from different cells are enriched at a specific time, according to NASH development | [45] |
| Extracellular vesicles | Serum | C56BL/6J mice fed high-fat diets and underwent aerobic training | MicroRNAs (especially miR-122, miR-192, and miR-22) | Hepatocytes, adipocytes | Serum EVs miR-22 expression was associated with adipogenesis and insulin sensitivity markers in adipocytes. Liver PPARγ expression was negatively correlated with serum miR-122 level | Aerobic training prevented obesity-induced steatohepatitis | [43] |
| Extracellular vesicles | Plasma, hepatocytes | C56BL/6J male mice fed high-fat diets | S1P | BMDM and HSCs | Circulating EVs were enriched in mice with high-fat diets | Activation of inflammatory phenotype in macrophages | [55] |
| Extracellular vesicles | Hepatocytes | C57BL/6J mice fed high-fat diets | MicroRNAs (especially miR-122, let-7e-5p, miR-31-5p and miR-210-3p) | Adipocytes | Increased miR-122, let-7e-5p, miR-31-5p and miR-210-3p expression in adipocytes | Hepatocyte-EVs increased fat accumulation and the expression of lipogenesis genes | [56] |
| Extracellular vesicles | Hepatocytes | HepG2 cells treated with cobalt chloride (CoCl2) or excess fatty acids | - | HSCs | Hepatocyte-EVs increased the expression of the pro-fibrotic markers of TGFβ-1, CTGF, COL1A1, and α-SMA in HSCs | Activation of the fibrosis and HSCs | [57] |
| Extracellular vesicles | Hepatocytes | HepG2 cells treated with cobalt chloride (CoCl2) or excess fatty acids | - | Kupffer cells | Hepatocyte-EVs have enrichment of the pro-inflammatory cytokines and inflammasomes (interleukin-1β, NLRP3, and ASC). Hepatocyte-EVs induced chemotaxis in Kupffer cells | Lipotoxic hepatocytes-EVs activated Kupffer cells chemotaxis | [58] |
| Extracellular vesicles | Hepatocytes | Hepatocytes treated with palmitate | MicroRNAs (especially miR-1) | Human umbilical vein endothelial cells (HUVECs) | miR-1 suppressed expression of KLF-4 and increased the NF-κB activity | Hepatocyte-EVs induced endothelial cell inflammation | [59] |
| Extracellular Vesicles | Cell Source | Molecular Mediators in the EVs Cargos | Recipient Targets Model | Interaction | Clinical Relevance | Reference |
|---|---|---|---|---|---|---|
| Exosomes | Human umbilical cord MSCs (hucMSC) | mRNA, surface adhesion molecules | Acute liver injury mice model (CCl4 treatment) | hucMSC exosomes recovered AST activity, reduced COL1A1, COL3A1, and TGF-β1 expressions | Alleviation of liver fibrosis | [73] |
| Exosomes | hucMSC | GPX1 | Acute liver injury mice model (CCl4 treatment) | Reduction of hepatic ROS and apoptosis by increasing the ERK1/2 and BCL-2 and decreasing the IKKB/NFkB/Casp-9/-3 pathway | The recovery of hepatic oxidant injury | [81] |
| Exosomes | hucMSC | - | Acute liver injury mice model (LPS and D-galactosamine treatment), RAW264.7 macrophages | Reduction of NLRP3, Casp-1, IL-1β, IL-6 expressions in the macrophage, liver ALT and AST levels, and the restoration of damaged liver tissue | Reduced inflammation and liver damage is repaired | [83] |
| Exosome | Chorionic plate-derived MSCs (CP-MSCs) | miR-125b | Acute liver injury mice model (CCl4 treatment), hepatic stellate cells (HSCs) | miR-125b suppressed the activation of Hh signaling that promotes fibrosis | Suppression of the HSCs activation and proliferation | [92] |
| Exosomes | MSCs | - | Acute liver injury mice model (CCl4 treatment), hepatocytes | MSCs exosomes activated proliferation genes and prevented apoptosis | MSC-derived exosomes have hepatoprotective effects against acute-liver injury | [74] |
| Exosomes | Adipose tissue-derived MSCs (AMSCs) | miR-17 | Acute liver injury mice model (LPS and D-galactosamine treatment), Kupffer cells | miR-17 reduced TXNIP expression and suppressed the NLRP3 inflammasome activation in Kupffer cells | Reduction of inflammatory activation in Kupffer cells | [89] |
| Exosomes | AMSCs | miR-181-5p | Acute liver injury mice model (CCl4 treatment), HSCs | miR-181-5p increased autophagy and reduced liver fibrosis by inhibiting the STAT3/BCL-2/Beclin-1 pathway HSCs COL1A1, VIMENTIN, α-SMA, and FN1 expressions were reduced | AMSCs exosomal miR-181-5p has an anti-fibrotic role | [93] |
| Exosomes | AMSCs | miR-122 | Acute liver injury mice model (CCl4 treatment), HSCs | miR-122 reduced the expression of IGF1R, CCNG1, and P4HA1 in HSCs | Suppression of the HSCs proliferation and collagen maturation | [94] |
| Exosome | Adipose-derived stem cells (ADSC) | STAT3 | Mice fed high-fat diets, macrophages | ADSC exosomes improved insulin sensitivity, reduced obesity, and alleviated hepatic steatosis, by inducing the anti-inflammatory phenotypes in M2 macrophages via the transactivation of arginase-1 by exosome-STAT3 | Improvement of insulin regulation and hepatic steatosis | [72] |
| Exosomes | Bone-marrow-derived MSC (BMSCs) | - | Acute liver injury mice model (CCl4 treatment), hepatocytes (Acetaminophen or hydrogen peroxide treatment) | Reduced ROS production and prevented oxidative stress, as well as improved liver regeneration and recovery | The recovery of hepatic oxidant injury | [76] |
| Exosomes | BMSCs | - | Hepatocytes (LPS and D-galactosamine treatment) | BMSCs exosomes reduced the pro-apoptotic proteins BAX, and cleaved Casp-3, and increased the expression of the anti-apoptotic BCL-2 | Induce autophagy and protect hepatic cells from damage caused by various stresses by mediating autophagy | [80] |
| Exosome | BMSCs | - | Acute liver injury mice model (CCl4 treatment), HSCs | BMSCs exosomes alleviated liver fibrosis and inflammation, as well as reduced the expression of Wnt/β-catenin pathway components (PPARγ, Wnt3a, Wnt10b, β-catenin, WISP1, CCND1, α-SMA, and COL1A1) in HSCs and liver tissue | Alleviation of liver fibrosis via the inhibition of Wnt/β-catenin signaling | [79] |
| Exosomes | Human-induced pluripotent stem cell-derived mesenchymal stromal cells (hiPSC-MSCs) | - | Liver injury mice model (ischemia/reperfusion surgery), hepatocytes | hiPSC-MSCs exosomes reduced AST and ALT levels and increased primary hepatocyte proliferation and synthesis of S1P | Protection against hepatic ischemia/reperfusion injury | [86] |
| Exosomes | Human menstrual blood-derived stem cells (MenSCs) | ICAM-1, angiopoietin-2, Axl, angiogenin, IGFBP-6, osteoprotegerin, IL-6, and IL-8 | Acute liver injury mice model (LPS and D-galactosamine treatment), AML12 macrophage cells | MenSCs exosomes improved liver function and inhibited apoptosis with a reduction of active Casp-3 | Inhibition of cell apoptosis and enhanced survival | [87] |
| Microvesicles (MVs) | Human liver stem cells (HLSC) | mRNAs | Hepatocytes | HLSC MVs activated cell proliferation and liver regeneration | Liver regeneration | [84] |
| Extracellular vesicles | HLSC | ASS1 protein and mRNA | Hepatocytes derived from ASS1 deficient HLSC | HLSC EVs restored ASS1 activity and urea production | Restoration of ASS1 function in deficient cells | [85] |
| Extracellular vesicles | HLSC | NASH mice model (choline-deficient amino acid diet) | HLSC EVs reduced fibrosis and inflammation markers (α-SMA), COL1A1, TGF-β1, TNF-α, IL-1β, and LTBP1 | Reduction of inflammation and fibrogenesis | [98] | |
| Extracellular vesicles | hucMSC | MnSOD enzyme | Liver injury mice model (ischemia/reperfusion surgery) | hucMSC EVs reduced neutrophils infiltration and alleviated hepatic oxidative stress | Inhibition of the oxidative stress and neutrophil inflammatory response | [82] |
| Extracellular vesicles | hucMSC | - | Liver injury mice model (S. japonicum infection), HSCs | hUCMSC EVs ameliorated liver injury and reduced the expression of α-SMA, COL1A1, and COL3A1, as well as HSCs proliferation | Suppression of HSCs proliferation and improved liver condition | [99] |
| Extracellular vesicles | Amnion-derived mesenchymal stem | - | NASH mice model (high-fat diets), Acute liver injury mice model (CCl4 treatment), HSCs and Kupffer cells | AMSC EVs reduced the expression of pro-inflammatory cytokines (TNF-α, IL-1β, IL-6, and TGF-β), fibrosis, Kupffer cell numbers, and HSC activation | Reduction of inflammation and fibrogenesis | [97] |
| Extracellular vesicles | BMSCs | Y-RNA-1 | Liver failure mice model (D-galactosamine/TNF-α treatment), hepatocytes | BMSCs EVs reduced hepatic injury and apoptosis | Protective effect against hepatocyte apoptosis | [75] |
| Extracellular vesicles | BMSCs | - | Liver injury mice model (ischemia/reperfusion surgery), hepatocytes | BMSCs EVs reduced tissue necrosis, apoptosis, serum ALT, and increased expression of NLRP12 and CXCL1, as well as increased the expression of IL-6 | Reduction of tissue necrosis, inflammation, and apoptosis | [77] |
| Extracellular vesicles | Human mesenchymal stromal cell (hMSCs) | - | Liver injury mice model (ischemia/reperfusion surgery) | hMSCs EVs reduced hepatic necrosis and inflammatory genes (HMBG-1, ICAM-1, HO-1, and IL-1β) | Reduction of tissue necrosis and inflammation | [78] |
| Extracellular vesicles | Human embryonic stem cell-derived mesenchymal stroma cells | - | Liver injury mice model (thioacetamide treatment) | EVs reduced fibrosis, apoptosis, and regenerated liver cells | Regeneration of liver | [64] |
| Extracellular vesicles | Human adipose-derived stem cells (hASCs) | lncRNA H19 | Acute liver injury mice model (D-galactosamine treatment) | hASCs EVs reduced the expression of inflammatory mediators and chemotactic factors | Inhibition of the liver inflammation | [90] |
| Extracellular vesicles | hASCs | - | NASH mice model (high-fat diets) with acute liver injury (LPS treatment) | hASCs EVs reduced serum ALT levels and inflammatory markers and macrophages | Inhibition of the liver inflammation | [95] |
| Extracellular vesicles | Human induced pluripotent stem cell (iPSCs) | MicroRNAs (specifically miR-92a-3p) | HSCs | iPSCs EVs reduced pro-fibrogenic markers (α–SMA, COL1A1, FN1, and TIMP-1), and HSC proliferation | Inhibition of fibrosis and HSCs proliferation | [96] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dorairaj, V.; Sulaiman, S.A.; Abu, N.; Abdul Murad, N.A. Extracellular Vesicles in the Development of the Non-Alcoholic Fatty Liver Disease: An Update. Biomolecules 2020, 10, 1494. https://doi.org/10.3390/biom10111494
Dorairaj V, Sulaiman SA, Abu N, Abdul Murad NA. Extracellular Vesicles in the Development of the Non-Alcoholic Fatty Liver Disease: An Update. Biomolecules. 2020; 10(11):1494. https://doi.org/10.3390/biom10111494
Chicago/Turabian StyleDorairaj, Vicneswarry, Siti Aishah Sulaiman, Nadiah Abu, and Nor Azian Abdul Murad. 2020. "Extracellular Vesicles in the Development of the Non-Alcoholic Fatty Liver Disease: An Update" Biomolecules 10, no. 11: 1494. https://doi.org/10.3390/biom10111494
APA StyleDorairaj, V., Sulaiman, S. A., Abu, N., & Abdul Murad, N. A. (2020). Extracellular Vesicles in the Development of the Non-Alcoholic Fatty Liver Disease: An Update. Biomolecules, 10(11), 1494. https://doi.org/10.3390/biom10111494