
The Role of Lipids in the Process of Hair Ageing


 ,
,  and
and
Abstract
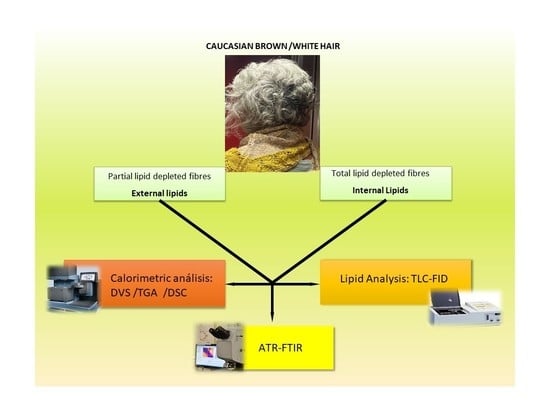
1. Introduction
2. Materials and Methods
2.1. Hair
2.2. Lipid Extraction
2.3. Lipid Analyses
2.4. Calorimetric Analysis
2.5. Moisture Content
2.6. Dynamic Water Vapour Sorption
2.7. Infrared Analyses Using FTIR
3. Results and Discussion
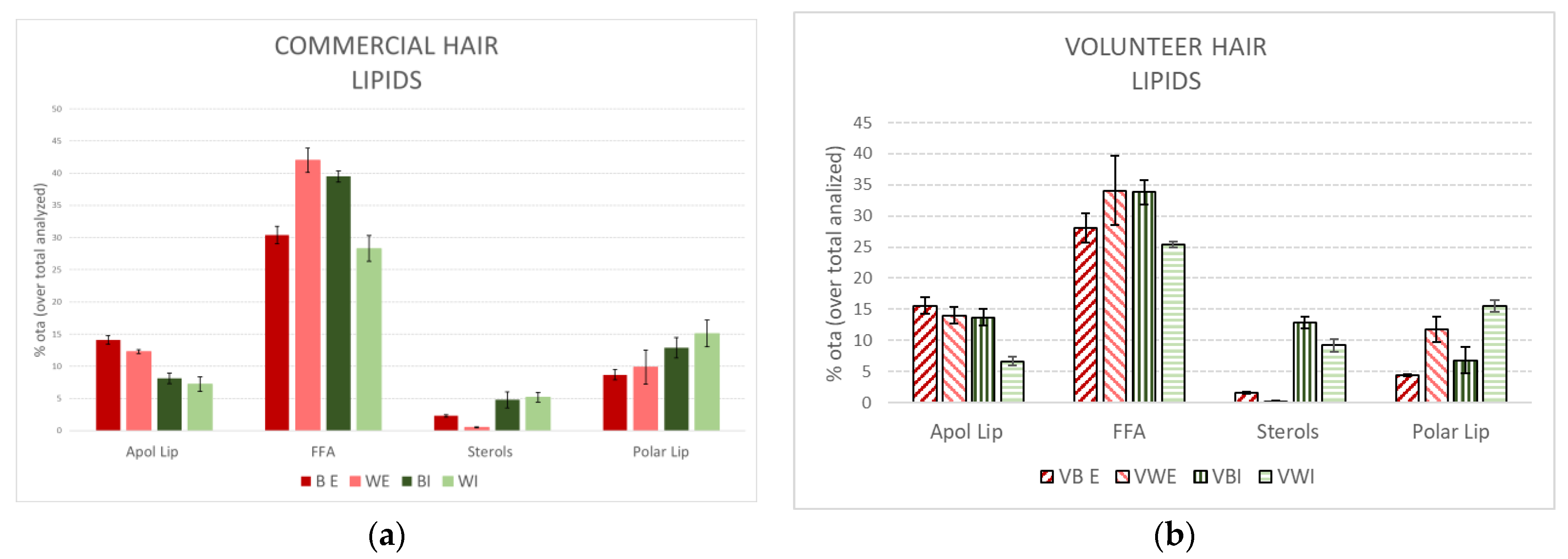
3.1. Lipid Extraction
3.2. Thermogravimetric Analysis of Extracted Lipids
3.3. Hair Moisture
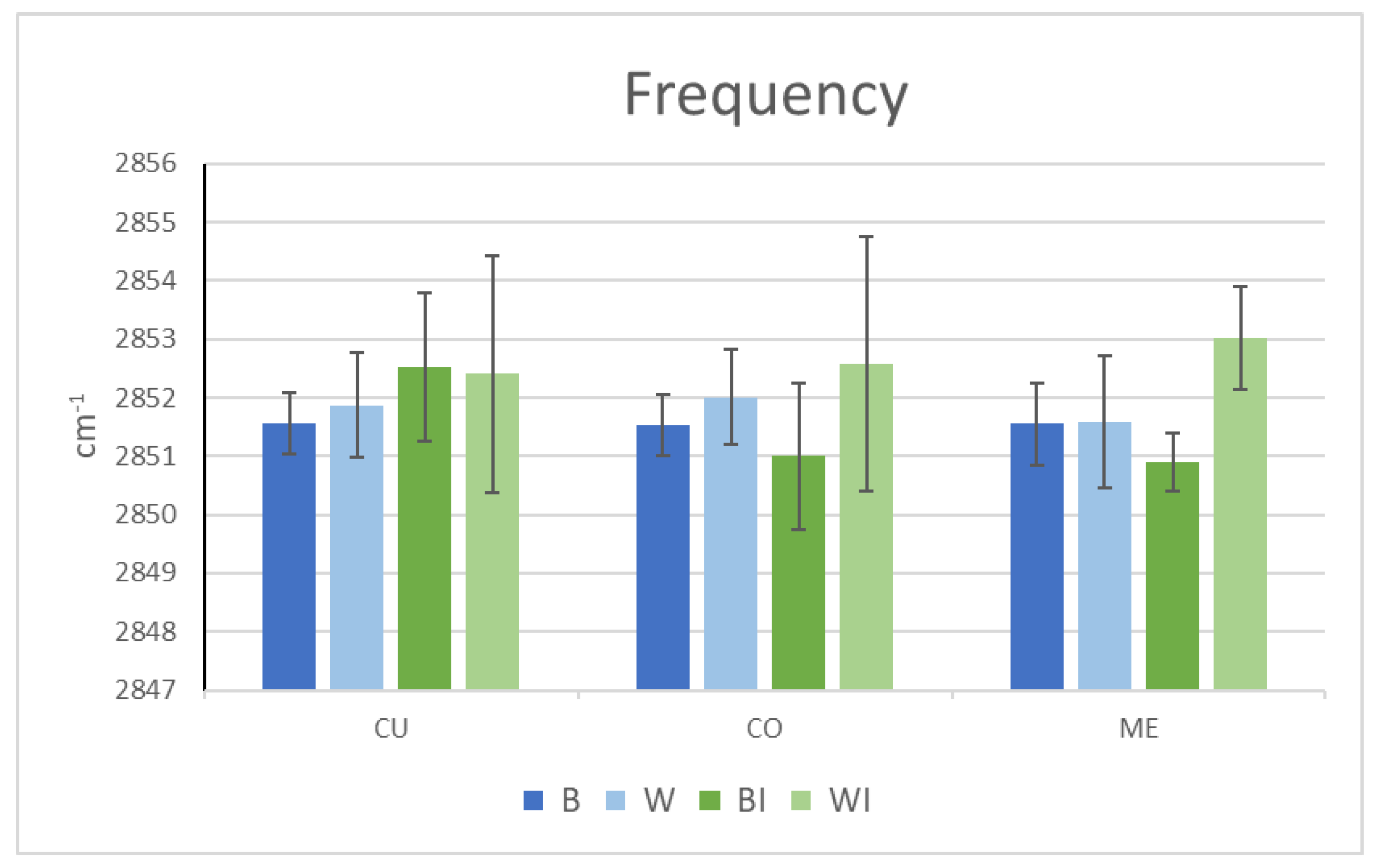
3.4. Hair IR
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Triwongwaranat, D.; Thuangtong, R.; Arunkajohnsak, S. A review of the etiologies, clinical characteristics, and treatment of canities. Int. J. Dermatol. 2019, 58, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Trueb, R.M. Aging of hair. J. Cosmet. Dermatol. 2005, 4, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.J. Human hair pigmentation–biological aspects. Int. J. Cosmet. Sci. 2008, 30, 233–257. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J.; Plonka, P.M.; Schallreuter, K.U.; Paus, R.; Tobin, D.J. Hair follicle pigmentation. J. Investig. Dermatol. 2005, 124, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Swift, J.A. Transfer of melanin granules from melanocytes to the cortical cells of human hair. Nature 1964, 203, 976–977. [Google Scholar] [CrossRef] [PubMed]
- Swift, J.A. Fine details on the surface of human hair. Int. J. Cosmet. Sci. 1991, 13, 143–159. [Google Scholar] [CrossRef]
- Coderch, L.; Oliver, M.A.; Martinez, V.; Manich, A.M.; Rubio, L.; Martí, M. Exogenous and endogenous lipids of human hair. Skin Res. Techol. 2017, 23, 479–485. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, W.; Wang, B.; Meyers, M.A. Structure and mechanical behavior of human hair. Mater. Sci. Eng. C 2017, 73, 152–163. [Google Scholar] [CrossRef]
- Kreplak, L.; Briki, F.; Duvault, Y.; Doucet, J.; Merigoux, C.; Leroy, F.; Lévêque, J.L.; Miller, L.; Carr, G.L.; Williams, G.P.; et al. Profiling lipids across Caucasian and Afro-American hair transverse cuts, using synchrotron infrared microspectrometry. Int. J. Cosmet. Sci. 2001, 23, 369–374. [Google Scholar] [CrossRef]
- Bantignies, J.-L.; Fuchs, G.; Carr, G.L.; Williams, G.P.; Lutz, D.; Marull, S. Organic reagent interaction with hair spatially characterized by infrared microspectroscopy using synchrotron radiation. Int. J. Cosmet. Sci. 1998, 20, 381–394. [Google Scholar] [CrossRef]
- Oliver, M.A.; Martí, M.; Coderch, L.; Carrer, V.; Kreuzer, M.; Barba, C. Lipid loses and barrier function modifications of the brown-to white hair transition. Skin Res. Technol. 2019, 25, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Sandt, C.; Borondics, F. A new typology of human hair medullas based on lipid composition analysis by synchrotron FTIR microspectroscopy. Analyst 2021, 146, 3942–3954. [Google Scholar] [CrossRef] [PubMed]
- Arck, P.C.; Overall, R.; Spatz, K.; Liezman, C.; Handjiski, B.; Klapp, B.F.; Birch-Machin, M.A.; Peters, E.M.J. Towards a “free radical theory of graying”: Melanocyte apoptosis in the aging human hair follicle is an indicator of oxidative stress induced tissue damage. FASEB J. 2006, 20, 1567–1569. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.J. Aging of the hair follicle pigmentation system. Int. J. Trichol. 2009, 1, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Kaur, R.; Bala, I. Therapeutics of premature hair graying: A long journey ahead. J. Cosmet. Dermatol. 2019, 18, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.M.; Decker, H.; Hartmann, H.; Chavan, B.; Rokos, H.; Spencer, J.D.; Hasse, M.J.; Thornton, M.J.; Shalbaf, M.; Paus, R.; et al. Senile hair graying: H202mediated oxidative stress affects human hair color by blunting methionine sulfoxide repair. FASEB J. 2009, 23, 2065–2075. [Google Scholar] [CrossRef] [PubMed]
- Trueb, R.M.; Rezende, H.D.; Dias, M.F.R.G. A comment on the science of hair aging. Int. J. Trichol. 2018, 10, 245–254. [Google Scholar] [CrossRef]
- O’Sullivan, J.D.; Nicu, C.; Picard, M.; Chéret, J.; Bedogni, B.; Tobin, D.J.; Paus, R. The biology of human hair greying. Biol. Rev. 2021, 96, 107–128. [Google Scholar] [CrossRef]
- Csuka, D.A.; Csuka, E.A.; Juhász, M.L.W.; Sharma, A.N.; Mesinkovska, N.A. A systematic review on the lipid composition of human hair. Int. J. Dermatol. 2022. [Google Scholar] [CrossRef]
- Martí, M.; Barba, C.; Manich, A.M.; Rubio, L.; Alonso, C.; Coderch, L. The influence of hair lipids in ethnic hair properties. Int. J. Cosmet. Sci. 2016, 38, 77–84. [Google Scholar] [CrossRef]
- Bianchini, P.D.; Kaplan, T.; Polefka, G.; Grove, S.; Daly, L.; Jumbelic, D.; Harper, M.Ç.; Nori, T.; Evans, R.; Ramaprasad, R. Grey hair: Clinical investigation into changes in hair fibres with loss of pigmentation in a photoprotected populations. Int. J. Cosmet. Sci. 2011, 33, 171–182. [Google Scholar]
- Breakspear, S.; Smith, J.R.; Luengo, G. Effect of the covalently linked fatty acid 18-MEA on the nanotribology of hair’s outermost surface. J. Struct. Biol. 2005, 149, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, Y.D.; Pi, L.-Q.; Lee, S.Y.; Hong, H.; Lee, W.-S. Comparison of hair shaft damage after chemical treatment in Asian, White European, and African hair. Int. J. Dermatol. 2014, 53, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Masukawa, Y.; Tsujimura, H.; Narita, H. Liquid chromatographymass spectrometry for comprehensive profiling of ceramide molecules in human hair. J. Lipid Res. 2006, 47, 1559–1571. [Google Scholar] [CrossRef]
- Brosche, T.; Dressler, S.; Platt, D. Age-associated changes in integral cholesterol and cholesterol sulfate concentrations in human scalp hair and finger nail clippings. Aging Clin. Exp. Res. 2001, 13, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Hollfelder, B.; Blankenburg, G.; Wolfram, L.J.; Höcker, H. Chemical and physical properties of pigmented and non-pigmented hair (‘grey hair’). Int. J. Cosmet. Sci. 1995, 17, 87–89. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, J.; He, C. Exploration of potential lipid biomarkers for premature canities by UPLC-QTOF-MS analyses of hair follicle roots. Exp. Dermatol. 2020, 29, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Richena, M.; Rezende, C.A. Structure of photo-damaged white and naturally pigmented human hair. J. Photochem. Photobiol. B Biol. 2020, 202, 111673. [Google Scholar] [CrossRef] [PubMed]
- Vyumvuhore, R.; Verzeaux, L.; Gilardeau, S.; Bordes, S.; Aymard, E.; Manfait, M.; Closs, B. Investigation of the molecular structure of greying hair shafts. Int. J. Cosmet. Sci. 2021, 43, 332–340. [Google Scholar] [CrossRef]
- Barba, C.; Oliver, M.A.; Martí, M.; Kreuzer, M.; Coderch, L. Lipid distribution of ethnic hairs by -Fourier Transform Infrared Synchrotron Spectroscopy. Skin Res. Technol. 2021, 28, 75–83. [Google Scholar] [CrossRef]
- Vickerstaff, T. The Physical Chemistry of Dyeing; Oliver and Boyd: London, UK, 1954. [Google Scholar]
- Oliver, M.A.; Coderch, L.; Carrer, V.; Barba, C.; Martí, M. Ethnic hair: Thermoanalytical and spectroscopic differences. Skin Res. Technol. 2020, 26, 617–626. [Google Scholar] [CrossRef]
- Yousef, I.; Ribó, L.; Crisol, A.; Šics, I.; Ellis, G.; Ducic, T.; Kreuzer, M.; Benseny-Cases, N.; Quispe, M.; Dumas, P.; et al. MIRAS: The Infrared Synchrotron Radiation Beamline at ALBA. Synchrotron Radiat. News 2017, 30, 4–6. [Google Scholar] [CrossRef]
- Hussler, G.; Kaba, G.; Francois, A.M.; Saint-Leger, D. Isolation and identification of human hair ceramides. Int. J. Cosmet. Sci. 1995, 17, 197–206. [Google Scholar] [CrossRef]
- Masukawa, Y.; Narita, H.; Imokawa, G. Characterization of the lipid composition at proximal root regions of human hair. J. Cosmet. Sci. 2005, 56, 1–16. [Google Scholar] [CrossRef]
- Evans, T. Measuring the water content of hair. Cosmet Toil. 2014, 129, 1–5. [Google Scholar]
- Kamath, Y.K. Quantification of human hair moisturization with cosmetic products by Dynamis Vapor Sorption. J. Cosmet. Sci. 2020, 71, 303–320. [Google Scholar]
- Timmermann, E. Multilayer sorption parameters, BET or GAB values? Colloids Surf. A Physicochem. Eng. Asp. 2003, 220, 235–260. [Google Scholar] [CrossRef]
- Barba, C.; Martí, M.; Carilla, J.; Manich, A.M.; Coderch, L. Moisture sorption/desorption of protein fibers. Thermochim. Acta 2013, 552, 70–76. [Google Scholar] [CrossRef]
- Barba, C.; Martí, M.; Manich, A.M.; Carilla, J.; Parra, J.L.; Coderch, L. Water absorption/desorption of human hair and nails. Thermochim. Acta 2010, 503–504, 33–39. [Google Scholar] [CrossRef]
- Barba, C.; Martí, M.; Semenzato, A.; Baratto, G.; Manich, A.M.; Coderch, L. Effect of lipid modification on stratum corneum permeability. J. Therm. Anal. Calorim. 2015, 120, 297–305. [Google Scholar] [CrossRef]
- Bildstein, L.; Deniset-Besseau, A.; Pasini, I.; Mazilier, C.; Keuong, Y.W.; Dazzi, A.; Baghdadli, N. Discrete nanoscale distribution of hair lipids fails to provide humidity resistance. Anal. Chem. 2020, 92, 11498–11504. [Google Scholar] [CrossRef] [PubMed]
- McMullen, R.L.; Zhang, G. Investigation of the internal structure of human hair with Atomic Force Microscopy. J. Cosmet. Sci. 2020, 71, 117–131. [Google Scholar]
- Mendelsohn, R.; Flach, C.R.; Moore, D.J. Determination of molecular conformation and permeation in skin via IR spectroscopy, microscopy, and imaging. Biochim. Biophys. Acta (BBA)-Biomembr. 2006, 1758, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, K.; Nishino, M.; Inaoka, Y.; Kitada, Y.; Fukushima, M. Interrelationship between the hair lipids and the hair moisture. J. Cosmet. Sci. Soc. Jpn. 1989, 13, 134–139. [Google Scholar]
- Fellows, A.P.; Casford, M.T.L.; Davies, P.B. Using hybrid atomic force microscopy and infrared spectroscopy (AFM-IR) to identify chemical components of hair medulla on the nanoscale. J. Microsc. 2021, 284, 189–202. [Google Scholar] [CrossRef]
- Ymazaki, J.; Maeda, K. Analysis of lipids in the medulla of Japanese hair and their function. Cosmetics 2018, 5, 27. [Google Scholar] [CrossRef]
- Kaneta, D.; Goto, M.; Hagihara, M.; Leproux, P.; Couderc, V.; Egawa, M.; Kano, H. Visualizing intra-medulla lipids in human hair using ultra-multiplex CARS, SHG, and THG microscopy. Analyst 2021, 146, 1163–1168. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BE | WE | BI | WI | |
|---|---|---|---|---|
| Total Lip Ext (% owf) | 0.92 ± 0.11 | 0.81 ± 0.15 | 2.48 ± 0.39 | 1.52 ± 0.16 |
| Apol Lip (% owf) | 0.13 ± 0.07 | 0.10 ± 0.02 | 0.20 ± 0.07 | 0.11 ± 0.04 |
| FFA (% owf) | 0.28 ± 0.08 | 0.34 ± 0.06 | 0.98 ± 0.15 | 0.43 ± 0.15 |
| Sterols (% owf) | 0.04 ± 0.02 | 0.01 ± 0.00 | 0.12 ± 0.05 | 0.08 ± 0.01 |
| Polar Lip (% owf) | 0.08 ± 0.06 | 0.08 ± 0.02 | 0.32 ± 0.06 | 0.23 ± 0.08 |
| BE | WE | BI | WI | |||||
|---|---|---|---|---|---|---|---|---|
| DSC | °C | J/g | °C | J/g | °C | J/g | °C | J/g |
| 10.5 | −24.7 | 19.4 | −37.3 | 12 | −16.7 | 17.5 | −26.2 | |
| 30.3 | −11.5 | 41.7 | −4.4 | 43.8 | −4.6 | 38.0 | −2.5 | |
| 54.9 | −6.4 | --- | --- | --- | --- | 50.2 | −0.8 | |
| TGA | °C | % | °C | % | °C | % | °C | % |
| 242.8 | 36.1 | 247.1 | 34.1 | 249.0 | 64.6 | 241.3 | 63.0 | |
| --- | --- | --- | --- | 340.6 | 25.6 | 338.2 | 19.1 | |
| 457.8 | 23.9 | 463.0 | 26.3 | 461.5 | 3.3 | 445.2 | 5.5 | |
| B | BE | BI | W | WE | WI | |
|---|---|---|---|---|---|---|
| Moisture at 50% RH Grav | 9.61 | 9.97 | 10.99 | 8.13 | 9.83 | 10.76 |
| Regain at 95% RH (%) | 26.25 | 25.61 | 26.18 | 24.85 | 24.15 | 25.50 |
| Wm (%) | 0.0772 | 0.0732 | 0.0795 | 0.0789 | 0.0729 | 0.0793 |
| Cg | 5.915 | 6.548 | 5.667 | 5.813 | 6.460 | 5.455 |
| K | 0.7367 | 0.7480 | 0.7334 | 0.7209 | 0.7396 | 0.7260 |
| R2 | 0.9980 | 0.9977 | 0.9989 | 24.85 | 24.15 | 25.50 |
| Da absor (min−1 × 10−3) | 16.41 | 17.41 | 14.85 | 17.27 | 18.14 | 16.57 |
| Da desor 95% (min−1 × 10−3) | 23.67 | 24.12 | 22.55 | 24.13 | 24.22 | 23.84 |
| Da (min−1 × 10−3) | 20.46 | 21.17 | 19.05 | 21.04 | 21.52 | 20.53 |
| B | BE | BI | W | WE | WI | |
|---|---|---|---|---|---|---|
| λCH2 St. Asym. cm−1 | 2918.9 ± 0.1 | 2925.9 ± 0.3 | 2927.6 ± 0.2 | 2920.9 ± 0.6 | 2928.7 ± 2.4 | Undetectable |
| Intensity Reduction % | - | 18.5 | 23.9 | - | 16.6 | |
| λCH2 St. Sym. cm−1 | 2850.1 ± 0.3 | 2851.5 ± 0.4 | 2851.7 ± 0.3 | 2851.0 ± 0.8 | 2851.0 ± 0.8 | Undetectable |
| Intensity Reduction % | - | 20.0 | 29.3 | - | 20.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coderch, L.; Lorenzo, R.D.; Mussone, M.; Alonso, C.; Martí, M. The Role of Lipids in the Process of Hair Ageing. Cosmetics 2022, 9, 124. https://doi.org/10.3390/cosmetics9060124
Coderch L, Lorenzo RD, Mussone M, Alonso C, Martí M. The Role of Lipids in the Process of Hair Ageing. Cosmetics. 2022; 9(6):124. https://doi.org/10.3390/cosmetics9060124
Chicago/Turabian StyleCoderch, Luisa, Ritamaria Di Lorenzo, Marika Mussone, Cristina Alonso, and Meritxell Martí. 2022. "The Role of Lipids in the Process of Hair Ageing" Cosmetics 9, no. 6: 124. https://doi.org/10.3390/cosmetics9060124
APA StyleCoderch, L., Lorenzo, R. D., Mussone, M., Alonso, C., & Martí, M. (2022). The Role of Lipids in the Process of Hair Ageing. Cosmetics, 9(6), 124. https://doi.org/10.3390/cosmetics9060124