
All That Glitters Is Not Gold: The Other Insects That Fall into the Asian Yellow-Legged Hornet Vespa velutina ‘Specific’ Traps



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
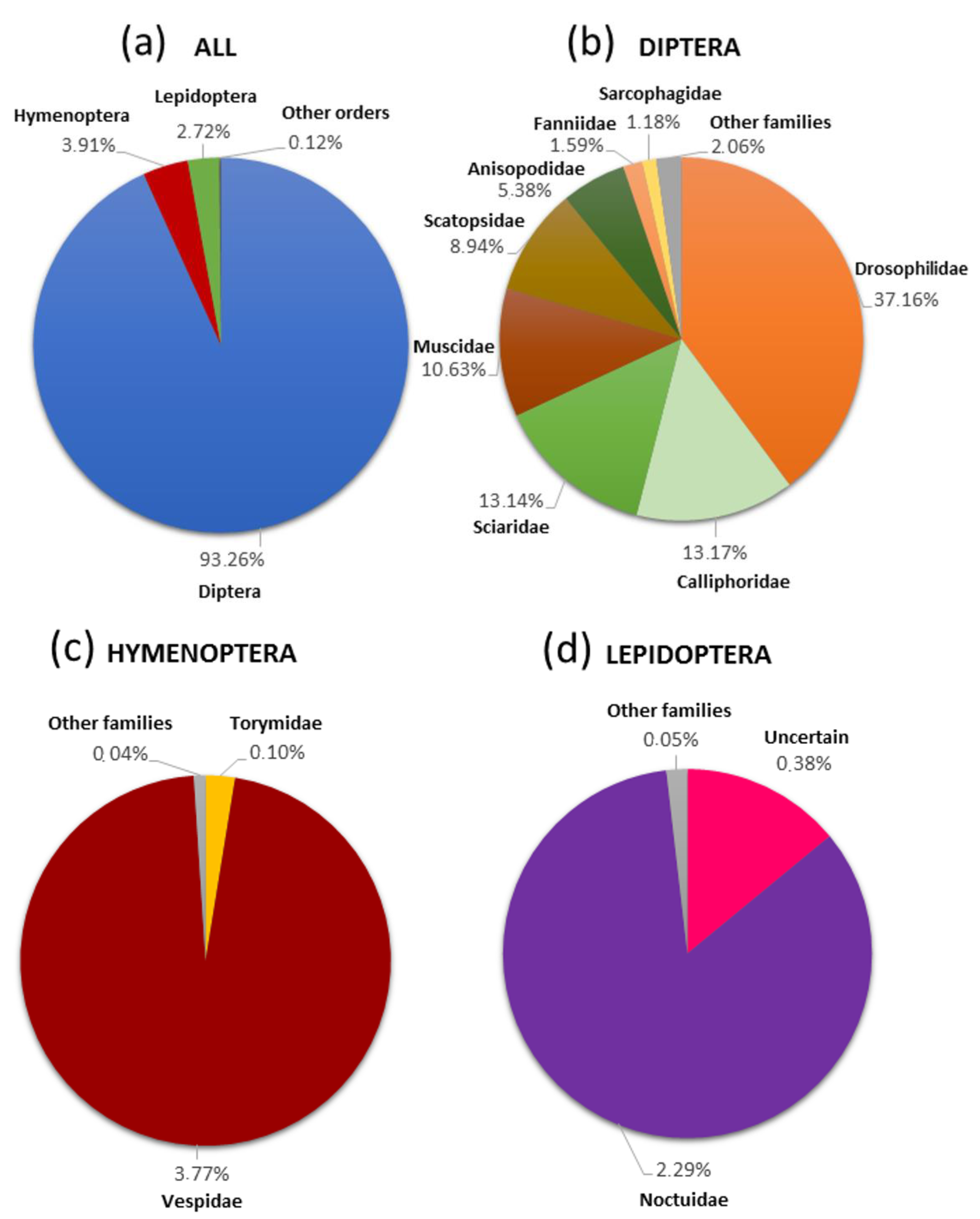
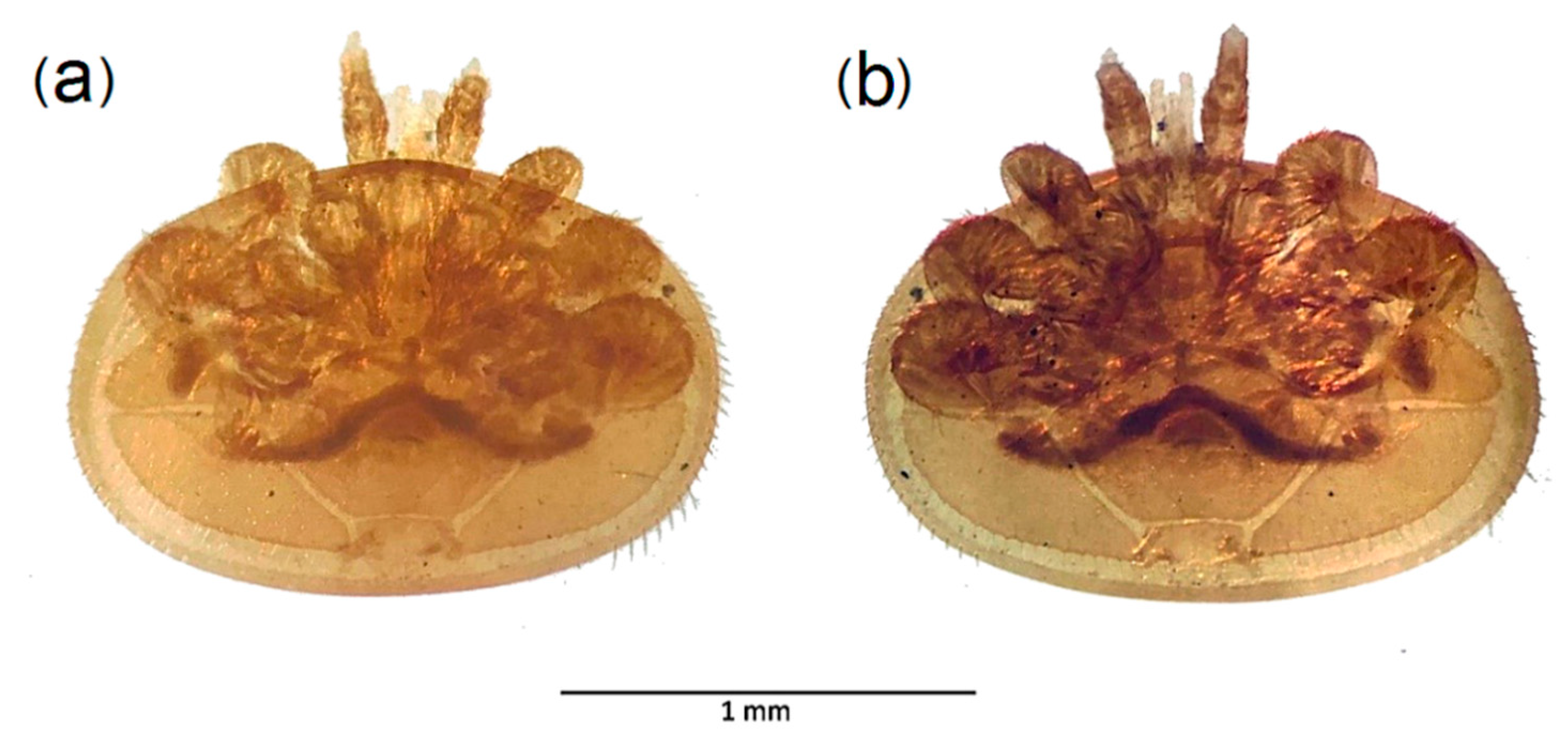
3.1. Specific Diversity of the Non-Target Insect Species
3.2. Ecosystem Services and Disservices
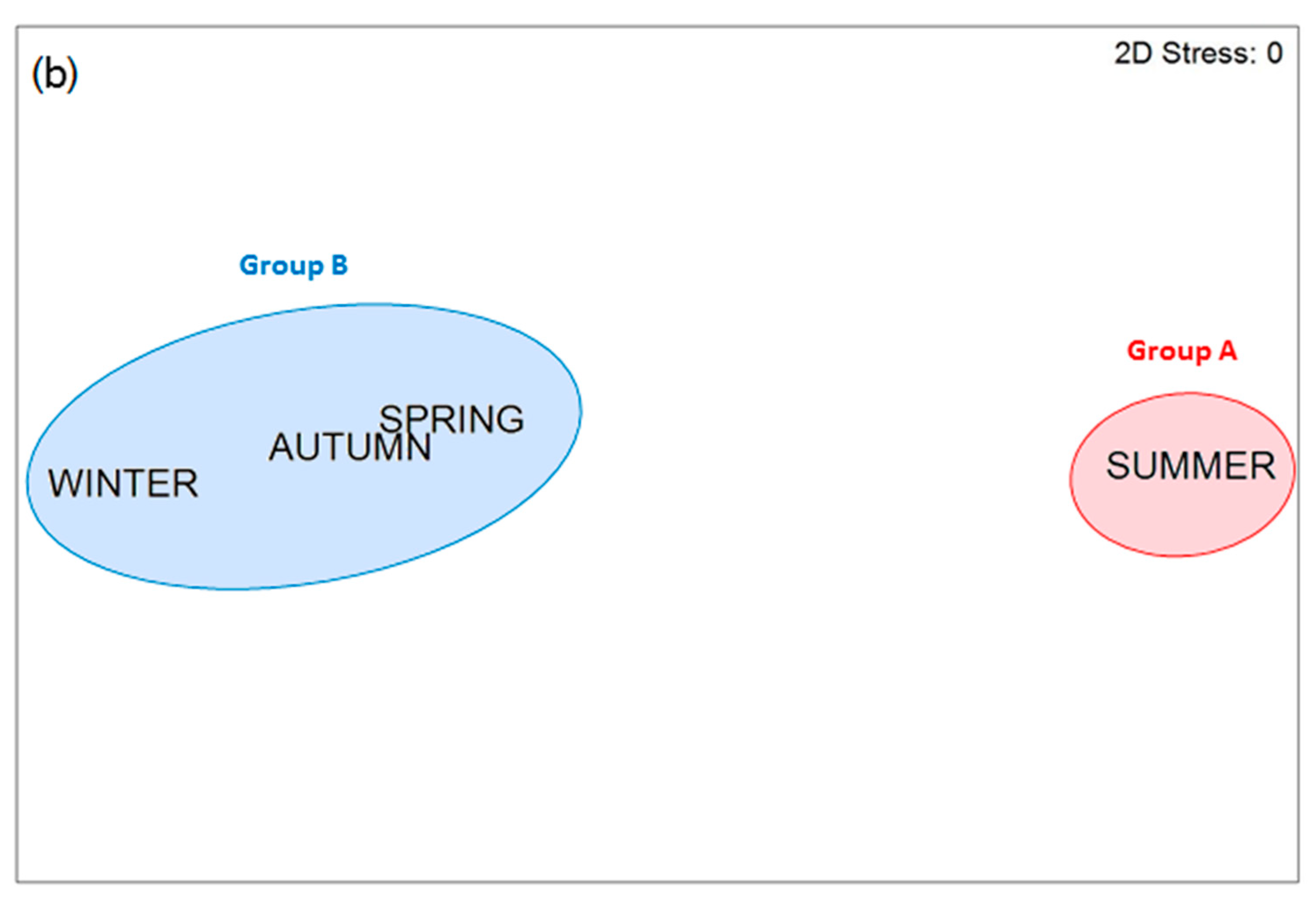
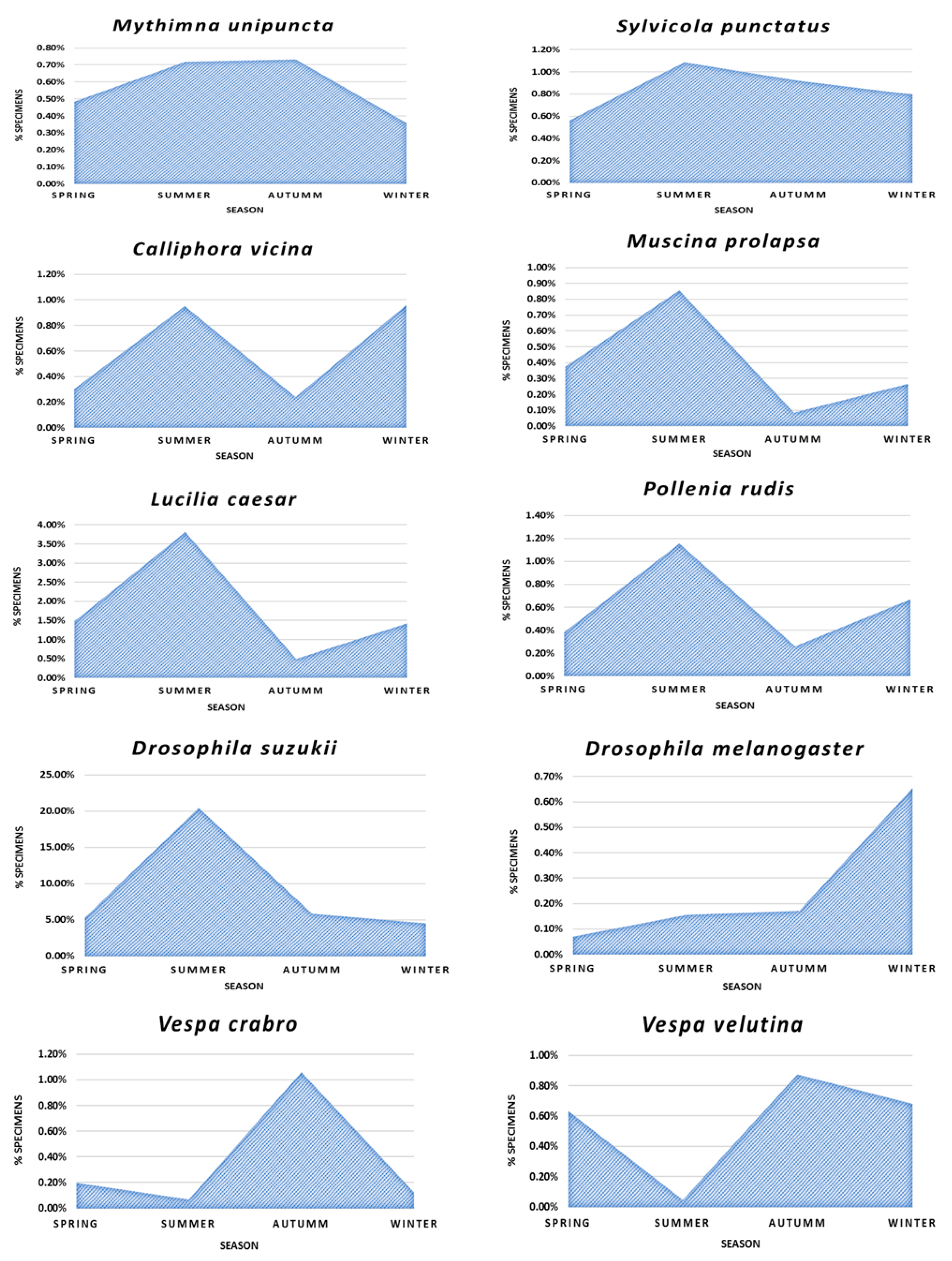
3.3. Phenological Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster/Taxa | % Contribution |
|---|---|
| Group A similarity (66.55%) | |
| Drosophila suzukii | 37.59 |
| Sciaridae sp. | 16.93 |
| Reichertella pulicaria | 7.52 |
| Lucilia caesar | 6.18 |
| Vespa velutina | 5.05 |
| Sylvicola punctatus | 4.94 |
| Mythimna unipuncta | 3.17 |
| Pollenia rudis | 2.32 |
| Calliphora vicina | 2.05 |
| Sylvicola cinctus | 1.99 |
| Fannia cf. postica | 1.73 |
| Muscina levida | 1.31 |
| Group B (Less than 2 samples in group) | |
| Group A & B dissimilarity (66.55%) | |
| Drosophila suzukii | 50.35 |
| Lucilia caesar | 8.83 |
| Reichertella pulicaria | 5.22 |
| Phaonia pallida | 4.28 |
| Sciaridae sp. | 3.49 |
| Pollenia rudis | 2.39 |
| Vespa velutina | 2.24 |
| Muscina prolapsa | 2.03 |
| Calliphora vicina | 1.55 |
| Phaonia bitincta | 1.55 |
| Dasyphora albofasciata | 1.38 |
| Vespa crabro | 1.31 |
References
- Haxaire, J.; Bouguet, J.P.; Tamisier, J.P. Vespa velutina Lepeletier, 1836, une redoutable nouveauté pour la faune de France (Hym., Vespidae). Bull. Soc. Entomol. Fr. 2006, 111, 194. [Google Scholar]
- Villemant, C.; Haxaire, J.; Streito, J.C. The discovery of the Asian hornet Vespa velutina in France. (La découverte du frelon asiatique Vespa velutina, en France). Insectes 2006, 143, 3–7. [Google Scholar]
- Caragata, C.R.; Montesinos, J.L.V. Datos ambientales preliminares del avispón asiático (Vespa velutina Lepeletier, 1836) (Hymenoptera, Vespidae) en Asturias, España. Bol. R. Soc. Esp. Hist. Nat. 2020, 114, 19–35. [Google Scholar]
- Barbet-Massin, M.; Rome, Q.; Muller, F.; Perrard, A.; Villemant, C.; Jiguet, F. Climate change increases the risk of invasion by the yellow-legged hornet. Biol. Conserv. 2013, 157, 4–10. [Google Scholar] [CrossRef]
- Castro, L.; Pagola-Carte, S. Vespa velutina Lepeletier, 1836 (Hymenoptera: Vespidae), recolectada en la Península Ibérica. Heteropterus Rev. Entomol. 2010, 10, 193–196. [Google Scholar]
- Rodríguez-Flores, M.S.; Seijo-Rodríguez, A.; Escuredo, O.; Seijo-Coello, M.C. Spreading of Vespa velutina in northwestern Spain: Influence of elevation and meteorological factors and effect of bait trapping on target and non-target living organisms. J. Pest. Sci. 2019, 92, 557–565. [Google Scholar] [CrossRef]
- Monceau, K.; Bonnard, O.; Thiéry, D. Vespa velutina: A new invasive predator of honeybees in Europe. J. Pest Sci. 2014, 87, 1–16. [Google Scholar] [CrossRef]
- Young, J.; Watt, A.; Nowicki, P.; Alard, D.; Clitherow, J.; Henle, K.; Johnson, R.; Laczko, E.; McCracken, D.; Matouch, S.; et al. Towards sustainable land use: Identifying and managing the conflicts between human activities and biodiversity conservation in Europe. Biodivers. Conserv. 2005, 14, 1641–1661. [Google Scholar] [CrossRef]
- Henle, K.; Alard, D.; Clitherow, J.; Cobb, P.; Firbank, L.; Kull, T.; McCracken, D.; Moritz, R.F.A.; Niemelä, J.; Rebane, M.; et al. Identifying and managing the conflicts between agriculture and biodiversity conservation in Europe—A review. Agric. Ecosyst. Environ. 2008, 124, 60–71. [Google Scholar] [CrossRef]
- Monceau, K.; Maher, N.; Bonnard, O.; Thiéry, D. Evaluation of competition between a native and an invasive hornet species: Do seasonal phenologies overlap? Bull. Entomol. Res. 2015, 105, 462–469. [Google Scholar] [CrossRef]
- López, S.; González, M.; Goldarazena, A. Vespa velutina lepeletier, 1836 (Hymenoptera: Vespidae): First records in Iberian Peninsula. Bull. OEPP 2011, 41, 439–441. [Google Scholar] [CrossRef]
- Haro, L.; Labadie, M.; Chanseau, P.; Cabot, C.; Blanc-Brisset, I.; Penouil, F. Medical consequences of the Asian black hornet (Vespa velutina) invasion in Southwestern France. Toxicon 2010, 55, 650–652. [Google Scholar] [CrossRef] [PubMed]
- Arca, M.; Papachristoforou, A.; Mougel, F.; Rortais, A.; Monceau, K.; Bonnard, O.; Tardy, P.; Thiéry, D.; Silvain, J.F.; Arnold, G. Defensive behaviour of Apis mellifera against Vespa velutina in France: Testing whether European honeybees can develop an effective collective defense against a new predator. Behav. Process. 2014, 106, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Budge, G.E.; Hodgetts, J.; Jones, E.P.; Ostojá-Starzewski, J.C.; Hall, J.; Tomkies, V.; Semmence, N.; Brown, M.; Wakefield, M.; Stainton, K. The invasion, provenance and diversity of Vespa velutina Lepeletier (Hymenoptera: Vespidae) in Great Britain. PLoS ONE 2017, 12, e0185172. [Google Scholar] [CrossRef] [PubMed]
- Leza, M.; Miranda, M.Á.; Colomar, V. First detection of Vespa velutina nigrithorax (Hymenoptera: Vespidae) in the Balearic Islands (Western Mediterranean): A challenging study case. Biol. Invasions 2018, 20, 1643–1649. [Google Scholar] [CrossRef]
- Lioy, S.; Manino, A.; Porporato, M.; Laurino, D.; Romano, A.; Capello, M.; Bertolino, S. Establishing surveillance areas for tackling the invasion of Vespa velutina in outbreaks and over the border of its expanding range. NeoBiota 2019, 46, 51–69. [Google Scholar] [CrossRef]
- Laurino, D.; Lioy, S.; Carisio, L.; Manino, A.; Porporato, M. Vespa velutina: An Alien Driver of HoneyBee Colony Losses. Diversity 2020, 12, 5. [Google Scholar] [CrossRef]
- Lioy, S.; Laurino, D.; Capello, M.; Romano, A.; Manino, A.; Porporato, M. Effectiveness and Selectiveness of Traps and Baits for Catching the Invasive Hornet Vespa velutina. Insects 2020, 11, 706. [Google Scholar] [CrossRef]
- Rojas-Nossa, S.V.; Novoa, N.; Serrano, A.; Calviño-Cancela, M. Performance of baited traps used as control tools for the invasive hornet Vespa velutina and their impact on non-target insects. Apidologie 2018, 49, 872–885. [Google Scholar] [CrossRef]
- Primack, R.B.; Silander, J.A. Measuring the relative importance of different pollinators to plants. Nature 1975, 255, 143–144. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Anderson, D.L.; Trueman, J.W.H. Varroa jacobsoni (Acari: Varroidae) is more than one species. Exp. Appl. Acarol. 2000, 24, 165–189. [Google Scholar] [CrossRef] [PubMed]
- Jelinski, M. Roztocz Varroa jacobsoni Oudemans, 1904 na larwach osy pospolitej Vespa (Paravespula) vulgaris L. Wiad Parazytol. 1990, 36, 55–58. [Google Scholar]
- Mordecai, G.J.; Brettell, L.E.; Pachori, P.; Villalobos, E.M.; Martin, S.J.; Jones, I.M.; Schroeder, D.C. Moku virus; a new Iflavirus found in wasps, honeybees and Varroa. Sci. Rep. 2016, 6, 34983. [Google Scholar] [CrossRef]
- Gállego, B.J. Manual de Parasitología: Morfología y Biología de Los Parásitos de Interés Sanitario; Universidad de Barcelona: Barcelona, Spain, 2006; pp. 1–517. [Google Scholar]
- Balachowsky, A.S. Entomologie Appliquée à 1’Agriculture. Tome II. Lepidópteros, Deuxième Volume; Masson, e.C., Ed.; Directeur De Publication: París, France, 1972; pp. 1345–1354. [Google Scholar]
- Scoble, M.J. The Lepidoptera. Form, Function and Diversity; Oxford University Press: Oxford, UK, 1992; pp. 1–404. [Google Scholar]
- Southwick, E.E.; Southwick, L. Estimating the economic value of honeybees (Hymenoptera: Apidae) as agricultural pollinators in the United States. J. Econ. Entomol. 1992, 85, 621–633. [Google Scholar] [CrossRef]
- Calabria, G.; Máca, J.; Bächil, G.; Serra, L.; Pascual, M. First records of the potencial pest species Drosophila suzukii (Diptera: Drosophilidae) in Europe. J. Appl. Entomol. 2012, 136, 139–147. [Google Scholar] [CrossRef]
- Bieńkowski, A.O.; Orlova-Bienkowskaja, M.J. Invasive Agricultural Pest Drosophila suzukii (Diptera, Drosophilidae) Appeared in the Russian Caucasus. Insects 2020, 11, 826. [Google Scholar] [CrossRef]
- Fiel, R.A.; Narganes, A.G.; Argüelles, M.B. Incidencia de “Drosophila suzukii” en cultivos de arándano y frambuesa en Asturias. Phytoma España 2014, 258, 49–53. [Google Scholar]
- Paradiso, F.; Martelli, F.; Cerrato, C.; Ghidotti, S.; Ramona, V.; Canterino, S.; Ferracini, C.; Bonelli, S. From Africa to the Alps: Risk assessment on an invasion by Cacyreus marshalli (Butler, 1898). J. Insect. Conserv. 2019, 23, 279–288. [Google Scholar] [CrossRef]
- Quacchia, A.; Ferracini, C.; Bonelli, S.; Balletto, E.; Alma, A. Can the Geranium Bronze, Cacyreus marshalli, become a threat for European biodiversity? Biodivers. Conserv. 2008, 17, 1429–1437. [Google Scholar] [CrossRef]
- Eitschberger, U.; Stamer, P. Cacyreus marshalli Butler, 1898, Eine neue Tagfalterart für sie Europaïsche Fauna. Lepidoptera, Lycaenidae). Atalanta 1990, 21, 101–108. [Google Scholar]
- Brou, V.A.; Brou, C.D. Mythimna unipuncta (Haworth, 1809) (Lepidoptera: Noctuidae) in Louisiana. South. Lepid. News 2020, 42, 31–33. [Google Scholar]
- Hill, D.S. Agricultural Insect Pest of the Tropics and Their Control; Alden Press: London, UK, 1983; pp. 1–749. [Google Scholar]








| Order | Family | Species | Author | N. of Specimens (Spring) | % of Insects (Spring) | N. of Specimens (Summer) | % of Insects (Summer) | N. of Specimens (Autumn) | % of Insects (Autumn) | N. of Specimens (Winter) | % of Insects (Winter) | N. of Specimens (Total) | % of Insects (Total) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diptera | Anisopodidae | Sylvicola cinctus | (Fabricius, 1787) | 32 | 0.25% | 70 | 0.55% | 34 | 0.26% | 71 | 0.55% | 207 | 1.61% |
| Diptera | Anisopodidae | Sylvicola fuscatus | (Fabricius, 1775) | 5 | 0.04% | 15 | 0.12% | 32 | 0.25% | 1 | 0.01% | 53 | 0.41% |
| Diptera | Anisopodidae | Sylvicola punctatus | (Fabricius, 1787) | 72 | 0.56% | 139 | 1.08% | 118 | 0.92% | 102 | 0.79% | 431 | 3.36% |
| Diptera | Calliphoridae | Bellardia viarum | (Robineau-Desvoidy, 1830) | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% |
| Diptera | Calliphoridae | Bellardia vulgaris | (Robineau-Desvoidy, 1830) | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% |
| Diptera | Calliphoridae | Calliphora vicina | Robineau-Desvoidy, 1830 | 39 | 0.30% | 122 | 0.95% | 31 | 0.24% | 123 | 0.96% | 315 | 2.45% |
| Diptera | Calliphoridae | Calliphora vomitoria | (Linnaeus, 1758) | 1 | 0.01% | 7 | 0.05% | 0 | 0.00% | 13 | 0.10% | 21 | 0.16% |
| Diptera | Calliphoridae | Chrysomya albiceps | (Wiedemann, 1819) | 2 | 0.02% | 3 | 0.02% | 0 | 0.00% | 2 | 0.02% | 7 | 0.05% |
| Diptera | Calliphoridae | Lucilia ampullacea | Villenueve, 1922 | 1 | 0.01% | 8 | 0.06% | 6 | 0.05% | 1 | 0.01% | 16 | 0.12% |
| Diptera | Calliphoridae | Lucilia caesar | (Linnaeus, 1758) | 189 | 1.47% | 488 | 3.80% | 63 | 0.49% | 183 | 1.43% | 923 | 7.19% |
| Diptera | Calliphoridae | Lucilia illustris | Meigen, 1826 | 4 | 0.03% | 8 | 0.06% | 3 | 0.02% | 11 | 0.09% | 26 | 0.20% |
| Diptera | Calliphoridae | Morinia doronici | (Scopoli, 1763) | 1 | 0.01% | 4 | 0.03% | 1 | 0.01% | 0 | 0.00% | 6 | 0.05% |
| Diptera | Calliphoridae | Onesia floralis | Robineau-Desvoidy, 1830 | 0 | 0.00% | 7 | 0.05% | 0 | 0.00% | 0 | 0.00% | 7 | 0.05% |
| Diptera | Calliphoridae | Pollenia griseotomentosa | (Jacentkovsky, 1944) | 6 | 0.05% | 0 | 0.00% | 0 | 0.00% | 0 | 0.00% | 6 | 0.05% |
| Diptera | Calliphoridae | Pollenia labialis | Robineau-Desvoidy, 1830 | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% |
| Diptera | Calliphoridae | Pollenia rudis | (Fabricius, 1794) | 49 | 0.38% | 148 | 1.15% | 33 | 0.26% | 86 | 0.67% | 316 | 2.46% |
| Diptera | Calliphoridae | Stomorhina lunata | (Fabricius, 1794) | 15 | 0.12% | 29 | 0.23% | 0 | 0.00% | 0 | 0.00% | 44 | 0.34% |
| Diptera | Chloropidae | Meromyza femorata | Macquart, 1835 | 0 | 0.00% | 3 | 0.02% | 0 | 0.00% | 0 | 0.00% | 3 | 0.02% |
| Diptera | Chloropidae | Thaumatomyia notata | (Meigen, 1830) | 5 | 0.04% | 3 | 0.02% | 9 | 0.07% | 24 | 0.19% | 41 | 0.32% |
| Diptera | Drosophilidae | Drosophila melanogaster | Meigen, 1830 | 9 | 0.07% | 20 | 0.16% | 22 | 0.17% | 84 | 0.65% | 135 | 1.05% |
| Diptera | Drosophilidae | Drosophila suzukii | (Matsumura, 1931) | 676 | 5.27% | 2629 | 20.48% | 744 | 5.80% | 580 | 4.52% | 4629 | 36.07% |
| Diptera | Drosophilidae | Gitona distigma | Meigen, 1830 | 0 | 0.00% | 3 | 0.02% | 0 | 0.00% | 2 | 0.02% | 5 | 0.04% |
| Diptera | Fanniidae | Fannia canicularis | (Linnaeus, 1761) | 1 | 0.01% | 10 | 0.08% | 42 | 0.33% | 36 | 0.28% | 89 | 0.69% |
| Diptera | Fanniidae | Fannia cf. postica | (Stein, 1895) | 25 | 0.19% | 4 | 0.03% | 36 | 0.28% | 50 | 0.39% | 115 | 0.90% |
| Diptera | Lauxanidae | Sapromyza opaca | Becker, 1895 | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% | 2 | 0.02% |
| Diptera | Muscidae | Dasyphora albofasciata | (Macquart in Webb & Berthelot, 1839) | 18 | 0.14% | 61 | 0.48% | 1 | 0.01% | 2 | 0.02% | 82 | 0.64% |
| Diptera | Muscidae | Mesembrina meridiana | (Linnaeus, 1758) | 2 | 0.02% | 1 | 0.01% | 1 | 0.01% | 1 | 0.01% | 5 | 0.04% |
| Diptera | Muscidae | Musca autumnalis | (De Geer, 1776) | 1 | 0.01% | 3 | 0.02% | 10 | 0.08% | 53 | 0.41% | 67 | 0.52% |
| Diptera | Muscidae | Musca domestica | Linnaeus, 1758 | 9 | 0.07% | 24 | 0.19% | 8 | 0.06% | 38 | 0.30% | 79 | 0.62% |
| Diptera | Muscidae | Musca tempestiva | Fallén, 1817 | 0 | 0.00% | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% |
| Diptera | Muscidae | Muscina levida | (Harris, 1780) | 17 | 0.13% | 44 | 0.34% | 32 | 0.25% | 37 | 0.29% | 130 | 1.01% |
| Diptera | Muscidae | Muscina pascuorum | (Meigen, 1826) | 6 | 0.05% | 22 | 0.17% | 1 | 0.01% | 0 | 0.00% | 29 | 0.23% |
| Diptera | Muscidae | Muscina prolapsa | (Harris, 1780) | 48 | 0.37% | 110 | 0.86% | 11 | 0.09% | 34 | 0.26% | 203 | 1.58% |
| Diptera | Muscidae | Muscina stabulans | (Fallén, 1817) | 18 | 0.14% | 31 | 0.24% | 0 | 0.00% | 1 | 0.01% | 50 | 0.39% |
| Diptera | Muscidae | Mydaea scutellaris | Robineau-Desvoidy, 1830 | 12 | 0.09% | 46 | 0.36% | 7 | 0.05% | 2 | 0.02% | 67 | 0.52% |
| Diptera | Muscidae | Mydaea urbana | (Meigen, 1826) | 2 | 0.02% | 3 | 0.02% | 3 | 0.02% | 0 | 0.00% | 8 | 0.06% |
| Diptera | Muscidae | Neomyia cornicina | (Fabricius, 1781) | 3 | 0.02% | 4 | 0.03% | 1 | 0.01% | 3 | 0.02% | 11 | 0.09% |
| Diptera | Muscidae | Phaonia bitincta | Róndani, 186 | 53 | 0.41% | 87 | 0.68% | 2 | 0.02% | 24 | 0.19% | 166 | 1.29% |
| Diptera | Muscidae | Phaonia pallida | (Fabricius, 1787) | 100 | 0.78% | 202 | 1.57% | 0 | 0.00% | 0 | 0.00% | 302 | 2.35% |
| Diptera | Muscidae | Polietes lardarius | Fabricius, 1781 | 3 | 0.02% | 0 | 0.00% | 8 | 0.06% | 152 | 1.18% | 163 | 1.27% |
| Diptera | Muscidae | Pyrellia vivida | Robineau-Desvoidy, 1830 | 0 | 0.00% | 0 | 0.00% | 2 | 0.02% | 0 | 0.00% | 2 | 0.02% |
| Diptera | Phoridae | Phoridae sp. | 6 | 0.05% | 27 | 0.21% | 22 | 0.17% | 59 | 0.46% | 114 | 0.89% | |
| Diptera | Sarcophagidae | Blaesoxipha cf. rossica | Villeneuve, 1912 | 1 | 0.01% | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 2 | 0.02% |
| Diptera | Sarcophagidae | Sarcophaga aff. argyrostoma | (Robineau–Desvoidy, 1830) | 6 | 0.05% | 10 | 0.08% | 0 | 0.00% | 2 | 0.02% | 18 | 0.14% |
| Diptera | Sarcophagidae | Sarcophaga cf. haemorrhoidalis | Böttcher, 1913 | 9 | 0.07% | 20 | 0.16% | 6 | 0.05% | 2 | 0.02% | 37 | 0.29% |
| Diptera | Sarcophagidae | Sarcophagidae sp. | 19 | 0.15% | 44 | 0.34% | 16 | 0.12% | 16 | 0.12% | 95 | 0.74% | |
| Diptera | Scathophagidae | Scathophaga stercolaria | (Linnaeus, 1758) | 3 | 0.02% | 0 | 0.00% | 11 | 0.09% | 61 | 0.48% | 75 | 0.58% |
| Diptera | Scatopsidae | Reichertella pulicaria | (Loew, 1846) | 141 | 1.10% | 229 | 1.78% | 115 | 0.90% | 663 | 5.17% | 1148 | 8.94% |
| Diptera | Sciaridae | Sciaridae sp. | 209 | 1.63% | 518 | 4.04% | 523 | 4.07% | 436 | 3.40% | 1686 | 13.14% | |
| Diptera | Sirphidae | Myatropa florea | (Linnaeus, 1758) | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% | 0 | 0.00% | 1 | 0.01% |
| Diptera | Tabanidae | Tabanus sudeticus | Zeller, 1842 | 0 | 0.00% | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% |
| Diptera | Trichoceridae | Trichocera annulata | Meigen, 1818 | 0 | 0.00% | 0 | 0.00% | 2 | 0.02% | 0 | 0.00% | 2 | 0.02% |
| Diptera | Ulidiidae | Physiphora alceae | (Preyssler, 1791) | 4 | 0.03% | 18 | 0.14% | 2 | 0.02% | 1 | 0.01% | 25 | 0.19% |
| Hymenoptera | Torymidae | Torymus sinensis | (Muller, 1764) | 0 | 0.00% | 0 | 0.00% | 0 | 0.00% | 5 | 0.04% | 5 | 0.04% |
| Hymenoptera | Torymidae | Torymus auratus | Kamijo, 1892 | 0 | 0.00% | 0 | 0.00% | 0 | 0.00% | 8 | 0.06% | 8 | 0.06% |
| Hymenoptera | Apidae | Apis mellifera | (Linnaeus, 1758) | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% |
| Hymenoptera | Apidae | Bombus terrestris lusitanicus | Krüger, 1956 | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% |
| Hymenoptera | Formicidae | Plagiolepsis sp. | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% | |
| Hymenoptera | Formicidae | Solenopsis sp. | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% | 0 | 0.00% | 1 | 0.01% | |
| Hymenoptera | Vespidae | Vespa crabro | Linnaeus, 1758 | 25 | 0.19% | 9 | 0.07% | 136 | 1.06% | 16 | 0.12% | 186 | 1.45% |
| Hymenoptera | Vespidae | Vespa velutina | Lepeletier, 1836 | 81 | 0.63% | 6 | 0.05% | 112 | 0.87% | 87 | 0.68% | 286 | 2.23% |
| Hymenoptera | Vespidae | Vespula germanica | (Fabricius, 1793) | 2 | 0.02% | 0 | 0.00% | 5 | 0.04% | 0 | 0.00% | 7 | 0.05% |
| Hymenoptera | Vespidae | Vespula vulgaris | (Linnaeus, 1758) | 3 | 0.02% | 0 | 0.00% | 2 | 0.02% | 0 | 0.00% | 5 | 0.04% |
| Lepidoptera | Uncertain | Lepidoptera sp. 1 | 17 | 0.13% | 31 | 0.24% | 0 | 0.00% | 0 | 0.00% | 48 | 0.37% | |
| Lepidoptera | Uncertain | Lepidoptera sp. 2 | 0 | 0.00% | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% | |
| Lepidoptera | Licenidae | Cacyreus marshalli | Butler, 1897 | 0 | 0.00% | 1 | 0.01% | 3 | 0.02% | 0 | 0.00% | 4 | 0.03% |
| Lepidoptera | Noctuidae | Mythimna unipuncta | (Haworth, 1809) | 62 | 0.48% | 92 | 0.72% | 94 | 0.73% | 46 | 0.36% | 294 | 2.29% |
| Lepidoptera | Nymphalidae | Pararge aegeria | (Linnaeus, 1758) | 2 | 0.02% | 0 | 0.00% | 0 | 0.00% | 0 | 0.00% | 2 | 0.02% |
| Blattodea | Blattidae | Blatta orientalis | Linnaeus, 1758 | 0 | 0.00% | 0 | 0.00% | 4 | 0.03% | 0 | 0.00% | 4 | 0.03% |
| Hemiptera | Pentatomidae | Nezara viridula | Linnaeus, 1758 | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% |
| Mecoptera | Panorpidae | Panorpa communis | Linnaeus, 1758 | 0 | 0.00% | 2 | 0.02% | 1 | 0.01% | 0 | 0.00% | 3 | 0.02% |
| Neuroptera | Chrysopidae | Pseudomellada clathratus | (Schneider, 1845) | 0 | 0.00% | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 1 | 0.01% |
| Neuroptera | Chrysopidae | Pseudomellada flavifrons | (Brauer, 1851) | 1 | 0.01% | 4 | 0.03% | 0 | 0.00% | 0 | 0.00% | 5 | 0.04% |
| Neuroptera | Chrysopidae | Pseudomellada marianus | (Navás, 1905) | 1 | 0.01% | 1 | 0.01% | 0 | 0.00% | 0 | 0.00% | 2 | 0.02% |
| 2024 | 15.77% | 5375 | 41.88% | 2317 | 18.05% | 3114 | 24.26% | 12,835 | 100% |
| Scientific Name | PO | DE | ADV | HDV | PC | AD | PA | PR | Status |
|---|---|---|---|---|---|---|---|---|---|
| Sylvicola cinctus | + | Native | |||||||
| Sylvicola fuscatus | + | Native | |||||||
| Sylvicola punctatus | + | Native | |||||||
| Bellardia viarum | + | + | Native | ||||||
| Bellardia vulgaris | + | + | Native | ||||||
| Calliphora vicina | + | + | + | Native | |||||
| Calliphora vomitoria | + | + | + | Native | |||||
| Chrysomya albiceps | + | + | + | Native | |||||
| Lucilia ampullacea | + | + | Native | ||||||
| Lucilia caesar | + | + | + | Native | |||||
| Lucilia illustris | + | + | + | Native | |||||
| Morinia doronici | + | + | Native | ||||||
| Onesia floralis | + | + | Native | ||||||
| Pollenia griseotomentosa | + | + | Native | ||||||
| Pollenia labialis | + | + | Native | ||||||
| Pollenia rudis | + | + | Native | ||||||
| Stomorhina lunata | + | + | + | Native | |||||
| Meromyza femorata | + | Native | |||||||
| Thaumatomyia notata | + | + | + | + | Native | ||||
| Drosophila melanogaster | + | Native | |||||||
| Drosophila suzukii | + | + | Invasive | ||||||
| Gitona distigma | + | Native | |||||||
| Fannia canicularis | + | Native | |||||||
| Fannia postica | + | Native | |||||||
| Sapromyza opaca | + | + | Native | ||||||
| Dasyphora albofasciata | + | + | Native | ||||||
| Mesembrina meridiana | + | + | + | Native | |||||
| Musca autumnalis | + | + | + | + | Native | ||||
| Musca domestica | + | + | + | + | Native | ||||
| Musca tempestiva | + | + | + | + | Native | ||||
| Muscina levida | + | + | + | + | + | Native | |||
| Muscina pascuorum | + | + | + | + | + | Native | |||
| Muscina prolapsa | + | + | + | + | + | Native | |||
| Muscina stabulans | + | + | + | + | + | Native | |||
| Mydaea Scutellaris | + | + | + | Native | |||||
| Mydaea urbana | + | + | + | Native | |||||
| Neomyia cornicina | + | + | Native | ||||||
| Phaonia bitincta | + | + | Native | ||||||
| Phaonia pallida | + | + | Native | ||||||
| Polietes lardarius | + | + | Native | ||||||
| Pyrellia vivida | + | + | Native | ||||||
| Phoridae sp.1 | + | NA | |||||||
| Blaesoxipha cf. rossica | + | + | Native | ||||||
| Sarcophaga aff. argyrostoma | + | + | Native | ||||||
| Sarcophaga cf. haemorrhoidalis | + | + | Native | ||||||
| Sarcophagidae sp. | + | + | NA | ||||||
| Scathophaga stercolaria | + | + | + | + | Native | ||||
| Reichertella pulicaria | + | Native | |||||||
| Sciaridae sp. | + | + | NA | ||||||
| Myatropa florea | + | + | Native | ||||||
| Tabanus sudeticus | + | + | + | Native | |||||
| Trichocera annulata | Native | ||||||||
| Physiphora alceae | + | Native | |||||||
| Torymus sinensis | + | + | Exotic | ||||||
| Torymus auratus | + | + | Native | ||||||
| Apis mellifera | + | Native | |||||||
| Bombus terrestris lusitanicus | + | Native | |||||||
| Plagiolepsis sp. | Native | ||||||||
| Solenopsis sp. | Native | ||||||||
| Vespa crabro | + | + | Native | ||||||
| Vespa velutina | + | + | Invasive | ||||||
| Vespula germanica | + | + | Native | ||||||
| Vespula vulgaris | + | + | Native | ||||||
| Lepidoptera sp. 1 | + | NA | |||||||
| Lepidoptera sp. 2 | + | NA | |||||||
| Cacyreus marshalli | + | + | Invasive | ||||||
| Mythimna unipuncta | + | + | Invasive | ||||||
| Pararge aegeria | + | Native | |||||||
| Blatta orientalis | + | + | Native | ||||||
| Nezara viridula | + | Native | |||||||
| Panorpa communis | + | + | Native | ||||||
| Pseudomallada clathratus | + | + | + | Native | |||||
| Pseudomallada flavifrons | + | + | + | Native | |||||
| Pseudomallada marianus | + | + | + | Native |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez, O.; Arias, A. All That Glitters Is Not Gold: The Other Insects That Fall into the Asian Yellow-Legged Hornet Vespa velutina ‘Specific’ Traps. Biology 2021, 10, 448. https://doi.org/10.3390/biology10050448
Sánchez O, Arias A. All That Glitters Is Not Gold: The Other Insects That Fall into the Asian Yellow-Legged Hornet Vespa velutina ‘Specific’ Traps. Biology. 2021; 10(5):448. https://doi.org/10.3390/biology10050448
Chicago/Turabian StyleSánchez, Omar, and Andrés Arias. 2021. "All That Glitters Is Not Gold: The Other Insects That Fall into the Asian Yellow-Legged Hornet Vespa velutina ‘Specific’ Traps" Biology 10, no. 5: 448. https://doi.org/10.3390/biology10050448
APA StyleSánchez, O., & Arias, A. (2021). All That Glitters Is Not Gold: The Other Insects That Fall into the Asian Yellow-Legged Hornet Vespa velutina ‘Specific’ Traps. Biology, 10(5), 448. https://doi.org/10.3390/biology10050448