Microbial Growth and Biogenic Amine Production in a Balkan-Style Fresh Sausage during Refrigerated Storage under a CO2-Containing Anaerobic Atmosphere: Effect of the Addition of Zataria multiflora Essential Oil and Hops Extract

 ,
,  , ,
, ,  , , ,
, , ,
Abstract
1. Introduction
2. Results and Discussion
2.1. Water Activity, pH, Microbial Contents, and Biogenic Amine Production
2.2. Identification of Lactic Acid Bacteria
3. Materials and Methods
3.1. Experimental Plan
3.2. Lamb Meat, Hops Extract, and Essential Oil
3.3. Sausage Manufacture
3.4. Analysis of Water Activity, pH, Microbial Content, and Biogenic Amine Production
3.5. Identification of Lactic Acid Bacteria
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cocolin, L.; Rantsiou, K.; Iacumin, L.; Urso, R.; Cantoni, C.; Comi, G. Study of the ecology of fresh sausages and characterization of populations of lactic acid bacteria by molecular methods. Appl. Environ. Microbiol. 2004, 70, 1883–1894. [Google Scholar] [CrossRef] [PubMed]
- González-Tenorio, R.; Fonseca, B.; Caro, I.; Fernández-Diez, A.; Kuri, V.; Soto, S.; Mateo, J. Changes in biogenic amine levels during storage of Mexican-style soft and Spanish-style dry-ripened sausages with different aw values under modified atmosphere. Meat Sci. 2013, 94, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Capillas, C.; Jiménez-Colmenero, F. Effect of an argon-containing packaging atmosphere on the quality of fresh pork sausages during refrigerated storage. Food Control 2010, 21, 1331–1337. [Google Scholar]
- Raimondi, S.; Nappi, M.R.; Sirangelo, T.M.; Leonardi, A.; Amaretti, A.; Ulrici, A.; Magnani, R.; Montanari, C.; Tabanelli, G.; Gardini, F.; et al. Bacterial community of industrial raw sausage packaged in modified atmosphere throughout the shelf life. Int. J. Food Microbiol. 2018, 280, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Martínez, L.; Djenane, D.; Cilla, I.; Beltrán, J.A.; Roncalés, P. Effect of different concentrations of carbon dioxide and low concentration of carbon monoxide on the shelf-life of fresh pork sausages packaged in modified atmosphere. Meat Sci. 2005, 71, 563–570. [Google Scholar] [CrossRef]
- Dias, F.S.; Ramos, C.L.; Schwan, R.F. Characterization of spoilage bacteria in pork sausage by PCR-DGGE analysis. Food Sci. Technol. 2013, 33, 468–474. [Google Scholar] [CrossRef]
- Fougy, L.; Desmonts, M.-H.; Coeuret, G.; Fassel, C.; Hamon, E.; Hézard, B.; Champomier-Vergès, M.-C.; Chaillou, S. Reducing salt in raw pork sausages increases spoilage and correlates with reduced bacterial diversity. Appl. Environ. Microbiol. 2016, 82, 3928–3939. [Google Scholar] [CrossRef]
- Ruiz-Capillas, C.; Pintado, T.; Jiménez-Colmenero, F. Biogenic amine formation in refrigerated fresh sausage “chorizo” keeps in modified atmosphere. J. Food Biochem. 2012, 36, 449–457. [Google Scholar] [CrossRef]
- Hugo, C.J.; Hugo, A. Current trends in natural preservatives for fresh sausage products. Trends Food Sci. Technol. 2015, 45, 12–23. [Google Scholar] [CrossRef]
- Hrnčič, M.K.; Španinger, E.; Košir, I.; Knez, Ž.; Bren, U. Hop compounds: Extraction techniques, chemical analyses, antioxidative, antimicrobial, and anticarcinogenic effects. Nutrients 2019, 11, 257. [Google Scholar] [CrossRef]
- Kramer, B.; Thielmann, J.; Hickisch, A.; Muranyi, P.; Wunderlich, J.; Hauser, C. Antimicrobial activity of hop extracts against foodborne pathogens for meat applications. J. Appl. Microbiol. 2015, 118, 648–657. [Google Scholar] [CrossRef]
- Bogdanova, K.; Kolar, M.; Langova, K.; Dusek, M.; Mikyska, A.; Bostikova, V.; Bostik, P.; Olsovska, J. Inhibitory effect of hop fractions against Gram-positive multi-resistant bacteria. A pilot study. Biomed. Pap. 2018, 162, 276–283. [Google Scholar] [CrossRef]
- Singh, M.; Smith, J.; Bailey, M. Using natural antimicrobials to enhance the safety and quality of poultry. In Handbook of Natural Antimicrobials for Food Safety and Quality; Taylor, T.M., Ed.; Woodhead Publishing: Cambridge, UK, 2014; pp. 375–401. [Google Scholar]
- Tiwari, B.K.; Valdramidis, V.P.; O’ Donnell, C.P.; Muthukumarappan, K.; Bourke, P.; Cullen, P.J. Application of natural antimicrobials for food preservation. J. Agric. Food Chem. 2009, 57, 5987–6000. [Google Scholar] [CrossRef] [PubMed]
- Calo, J.R.; Crandall, P.G.; O’Bryan, C.A.; Ricke, S.C. Essential oils as antimicrobials in food systems–A review. Food Control 2015, 54, 111–119. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Jayasena, D.D.; Jo, C. Essential oils as potential antimicrobial agents in meat and meat products: A review. Trends Food Sci. Technol. 2013, 34, 96–108. [Google Scholar] [CrossRef]
- Villalobos-Delgado, L.H.; Nevárez-Moorillon, G.V.; Caro, I.; Quinto, E.J.; Mateo, J. Natural antimicrobial agents to improve foods shelf life. In Food Quality and Shelf Life; Galanakis, C.M., Ed.; Elsevier: Amsterdam, The Nederland, 2019; pp. 125–157. [Google Scholar]
- Saei-Dehkordi, S.S.; Tajik, H.; Moradi, M.; Khalighi-Sigaroodi, F. Chemical composition of essential oils in Zataria multiflora Boiss. from different parts of Iran and their radical scavenging and antimicrobial activity. Food Chem. Toxicol. 2010, 48, 1562–1567. [Google Scholar] [CrossRef]
- Shakeri, M.-S.; Shahidi, F.; Beiraghi-Toosi, S.; Bahrami, A. Antimicrobial activity of Zataria multiflora Boiss. essential oil incorporated with whey protein based films on pathogenic and probiotic bacteria. Int. J. Food Sci. Technol. 2011, 46, 549–554. [Google Scholar] [CrossRef]
- Rezaeigolestani, M.; Misaghi, A.; Khanjari, A.; Basti, A.A.; Abdulkhani, A.; Fayazfar, S. Antimicrobial evaluation of novel poly-lactic acid based nanocomposites incorporated with bioactive compounds in-vitro and in refrigerated vacuum-packed cooked sausages. Int. J. Food Microbiol. 2017, 260, 1–10. [Google Scholar] [CrossRef]
- Basti, A.A.; Gandomi, H.; Noori, N.; Khanjari, A. Shirazi thyme (Zataria multiflora Boiss) Oils. In Essential Oils in Food Preservation, Flavor and Safety; Preedy, V.R., Ed.; Elsevier: Amsterdam, The Nederland, 2016; pp. 731–736. [Google Scholar]
- Tajik, H.; Aminzare, M.; Mounesi Raad, T.; Hashemi, M.; Hassanzad Azar, H.; Raeisi, M.; Naghili, H. Effect of Zataria multiflora Boiss essential oil and grape seed extract on the shelf life of raw buffalo patty and fate of onoculated Listeria monocytogenes. J. Food Process. Preserv. 2015, 39, 3005–3013. [Google Scholar] [CrossRef]
- Torab, M.; Basti, A.A.; Khanjari, A. Effect of free and nanoencapsulated forms of Zataria multiflora boiss. Essential oil on some microbial and chemical properties of beef burger. Carpathian J. Food Sci. Technol. 2017, 9, 93–102. [Google Scholar]
- Moradi, M.; Tajik, H.; Razavi Rohani, S.M.; Oromiehie, A.R. Effectiveness of Zataria multiflora Boiss essential oil and grape seed extract impregnated chitosan film on ready-to-eat mortadella-type sausages during refrigerated storage. J. Sci. Food Agric. 2011, 91, 2850–2857. [Google Scholar] [CrossRef]
- da Silveira, S.M.; Luciano, F.B.; Fronza, N.; Cunha, A.; Scheuermann, G.N.; Vieira, C.R.W. Chemical composition and antibacterial activity of Laurus nobilis essential oil towards foodborne pathogens and its application in fresh Tuscan sausage stored at 7 °C. LWT Food Sci. Technol. 2014, 59, 86–93. [Google Scholar] [CrossRef]
- Mastromatteo, M.; Incoronato, A.L.; Conte, A.; Del Nobile, M.A. Shelf life of reduced pork back-fat content sausages as affected by antimicrobial compounds and modified atmosphere packaging. Int. J. Food Microbiol. 2011, 150, 1–7. [Google Scholar] [CrossRef]
- Sharma, H.; Mendiratta, S.K.; Agrawal, R.K.; Gurunathan, K.; Kumar, S.; Singh, T.P. Use of various essential oils as bio preservatives and their effect on the quality of vacuum packaged fresh chicken sausages under frozen conditions. LWT Food Sci. Technol. 2017, 81, 118–127. [Google Scholar] [CrossRef]
- Šojić, B.; Pavlić, B.; Zeković, Z.; Tomović, V.; Ikonić, P.; Kocić-Tanackov, S.; Džinić, N. The effect of essential oil and extract from sage (Salvia officinalis L.) herbal dust (food industry by-product) on the oxidative and microbiological stability of fresh pork sausages. LWT 2018, 89, 749–755. [Google Scholar] [CrossRef]
- Lerasle, M.; Federighi, M.; Simonin, H.; Anthoine, V.; Rezé, S.; Chéret, R.; Guillou, S. Combined use of modified atmosphere packaging and high pressure to extend the shelf-life of raw poultry sausage. Innov. Food Sci. Emerg. Technol. 2014, 23, 54–60. [Google Scholar] [CrossRef]
- Pothakos, V.; Devlieghere, F.; Villani, F.; Björkroth, J.; Ercolini, D. Lactic acid bacteria and their controversial role in fresh meat spoilage. Meat Sci. 2015, 109, 66–74. [Google Scholar] [CrossRef]
- Benson, A.K.; David, J.R.D.; Gilbreth, S.E.; Smith, G.; Nietfeldt, J.; Legge, R.; Kim, J.; Sinha, R.; Duncan, C.E.; Ma, J.; et al. Microbial successions are associated with changes in chemical profiles of a model refrigerated fresh pork sausage during an 80-day shelf life study. Appl. Environ. Microbiol. 2014, 80, 5178–5194. [Google Scholar] [CrossRef]
- Narasimha Rao, D.; Sachindra, N.M. Modified atmosphere and vacuum packaging of meat and poultry products. Food Rev. Int. 2002, 18, 263–293. [Google Scholar] [CrossRef]
- Stanborough, T.; Fegan, N.; Powell, S.M.; Tamplin, M.; Chandry, P.S. Insight into the genome of Brochothrix thermosphacta, a problematic meat spoilage bacterium. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [PubMed]
- Samelis, J. Food Spoilage Microorganisms; Blackburn, C.D.W., Ed.; Woodhead Publishing: Cambridge, UK, 2006. [Google Scholar]
- Gribble, A.; Brightwell, G. Spoilage characteristics of Brochothrix thermosphacta and campestris in chilled vacuum packaged lamb, and their detection and identification by real time PCR. Meat Sci. 2013, 94, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Tremonte, P.; Sorrentino, E.; Succi, M.; Reale, A.; Maiorano, G.; Coppola, R. Shelf life of fresh sausages stored under modified atmospheres. J. Food Prot. 2005, 68, 2686–2692. [Google Scholar] [CrossRef] [PubMed]
- Chiavaro, E.; Zanardi, E.; Bottari, B.; Ianieri, A. Efficacy of different storage practices in maintaining the physicochemical and microbiological properties of fresh pork sausage. J. Muscle Foods 2008, 19, 157–174. [Google Scholar] [CrossRef]
- Vignolo, G.; Castellano, P.; Fontana, C.; Cocconcelli, P.S.; Fadda, S. Lactic acid bacteria in meat fermentations: Role of autochthonous starter. In Lactic Acid Bacteria: Microbiological and Functional Aspects; Vinderola, G., Ouwehand, A., Salminen, S., von Wright, A., Eds.; CRC Press: Boca Ratón, FL, USA, 2019; p. 764. [Google Scholar]
- Durlu-Özkaya, F.; Ayhan, K.; Vural, N. Biogenic amines produced by Enterobacteriaceae isolated from meat products. Meat Sci. 2001, 58, 163–166. [Google Scholar] [CrossRef]
- Suzzi, G.; Gardini, F. Biogenic amines in dry fermented sausages: A review. Int. J. Food Microbiol. 2003, 88, 41–54. [Google Scholar] [CrossRef]
- Nowak, A.; Czyzowska, A. In vitro synthesis of biogenic amines by Brochothrix thermosphacta isolates from meat and meat products and the influence of other microorganisms. Meat Sci. 2011, 88, 571–574. [Google Scholar] [CrossRef]
- Lu, S.; Ji, H.; Wang, Q.; Li, B.; Li, K.; Xu, C.; Jiang, C. The effects of starter cultures and plant extracts on the biogenic amine accumulation in traditional Chinese smoked horsemeat sausages. Food Control 2015, 50, 869–875. [Google Scholar] [CrossRef]
- Santos, M.H.S. Biogenic amines: Their importance in foods. Int. J. Food Microbiol. 1996, 29, 213–231. [Google Scholar] [CrossRef]
- Ruiz-Capillas, C.; Jiménez-Colmenero, F. Biogenic amines in meat and meat products. Crit. Rev. Food Sci. Nutr. 2005, 44, 489–599. [Google Scholar] [CrossRef]
- Stratton, J.E.; Hutkins, R.W.; Taylor, S.L. Biogenic amines in cheese and other fermented foods: A Review. J. Food Prot. 1991, 54, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Halász, A.; Baráth, Á.; Simon-Sarkadi, L.; Holzapfel, W. Biogenic amines and their production by microorganisms in food. Trends Food Sci. Technol. 1994, 5, 42–49. [Google Scholar] [CrossRef]
- Sattler, J.; Häfner, D.; Klotter, H.J.; Lorenz, W.; Wagner, P.K. Food-induced histaminosis as an epidemiological problem: Plasma histamine elevation and haemodynamic alterations after oral histamine administration and blockade of diamine oxidase (DAO). Agents Actions 1988, 23, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Del Blanco, A.; Caro, I.; Quinto, E.J.; Mateo, J. Quality changes in refrigerated stored minced pork wrapped with plastic cling film and the effect of glucose supplementation. Meat Sci. 2017, 126, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Fontana, C.A.; Fadda, S.; Cocconcelli, P.S.; Vignolo, G. Lactic acid bacteria in meat fermentations. In Lactic acid Bacteria: Microbiological and Functional Aspects; Lahtinen, S., Ouwehand, A.C., Salminen, S., von Wright, A., Eds.; CRC Press: Boca Ratón, FL, USA, 2011; pp. 247–259. [Google Scholar]
- Meat and Meat Products: Measurment of pH; ISO: 2917; International Organization for Standardization: Geneva, Switzerland, 1999.
- Eerola, S.; Hinkkanen, R.; Lindfors, E.; Hirvi, T. Liquid chromatographic determination of biogenic amines in dry sausages. J. AOAC Int. 1993, 76, 575–577. [Google Scholar]
- Clark, A.E.; Kaleta, E.J.; Arora, A.; Wolk, D.M. Matrix-assisted laser desorption ionization-time of flight mass spectrometry: A fundamental shift in the routine practice of clinical microbiology. Clin. Microbiol. Rev. 2013, 26, 547–603. [Google Scholar] [CrossRef]
- Patel, R. Matrix-assisted laser desorption ionization-time of flight mass spectrometry in clinical microbiology. Clin. Infect. Dis. 2013, 57, 564–572. [Google Scholar] [CrossRef]

 ), Lactobacillus spp. (
), Lactobacillus spp. ( ), Leuconostoc mesenteroides (
), Leuconostoc mesenteroides ( ), Enterococcus faecalis (
), Enterococcus faecalis ( ), Carnobacterium maltromaticum (
), Carnobacterium maltromaticum ( ), Lactobacillus curvatus (
), Lactobacillus curvatus ( ), Lactococcus lactis (
), Lactococcus lactis ( ).
), Lactobacillus spp. (), Leuconostoc mesenteroides (), Enterococcus faecalis (), Carnobacterium maltromaticum (), Lactobacillus curvatus (), Lactococcus lactis ().
).
), Lactobacillus spp. (), Leuconostoc mesenteroides (), Enterococcus faecalis (), Carnobacterium maltromaticum (), Lactobacillus curvatus (), Lactococcus lactis ().

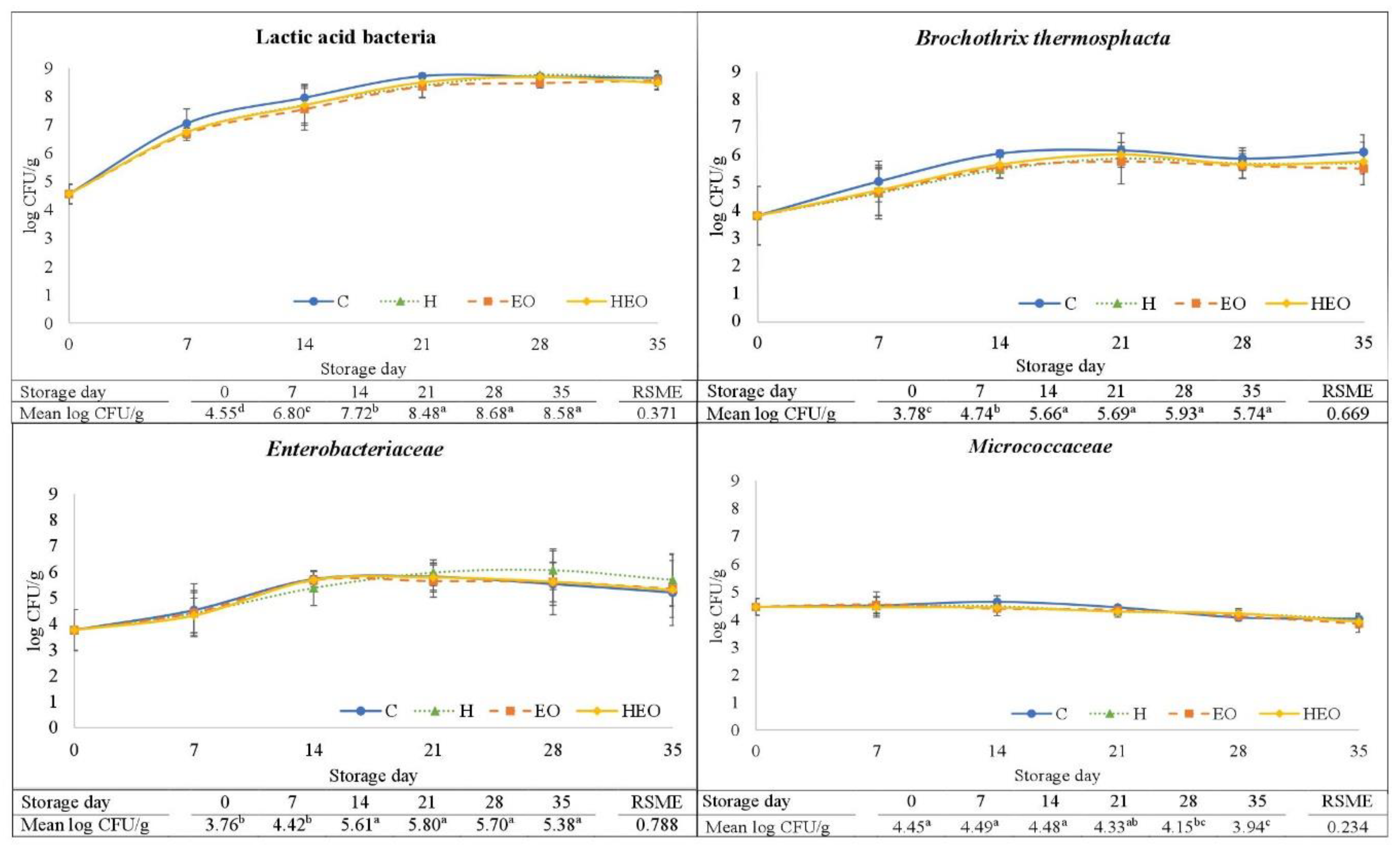{kind=link}
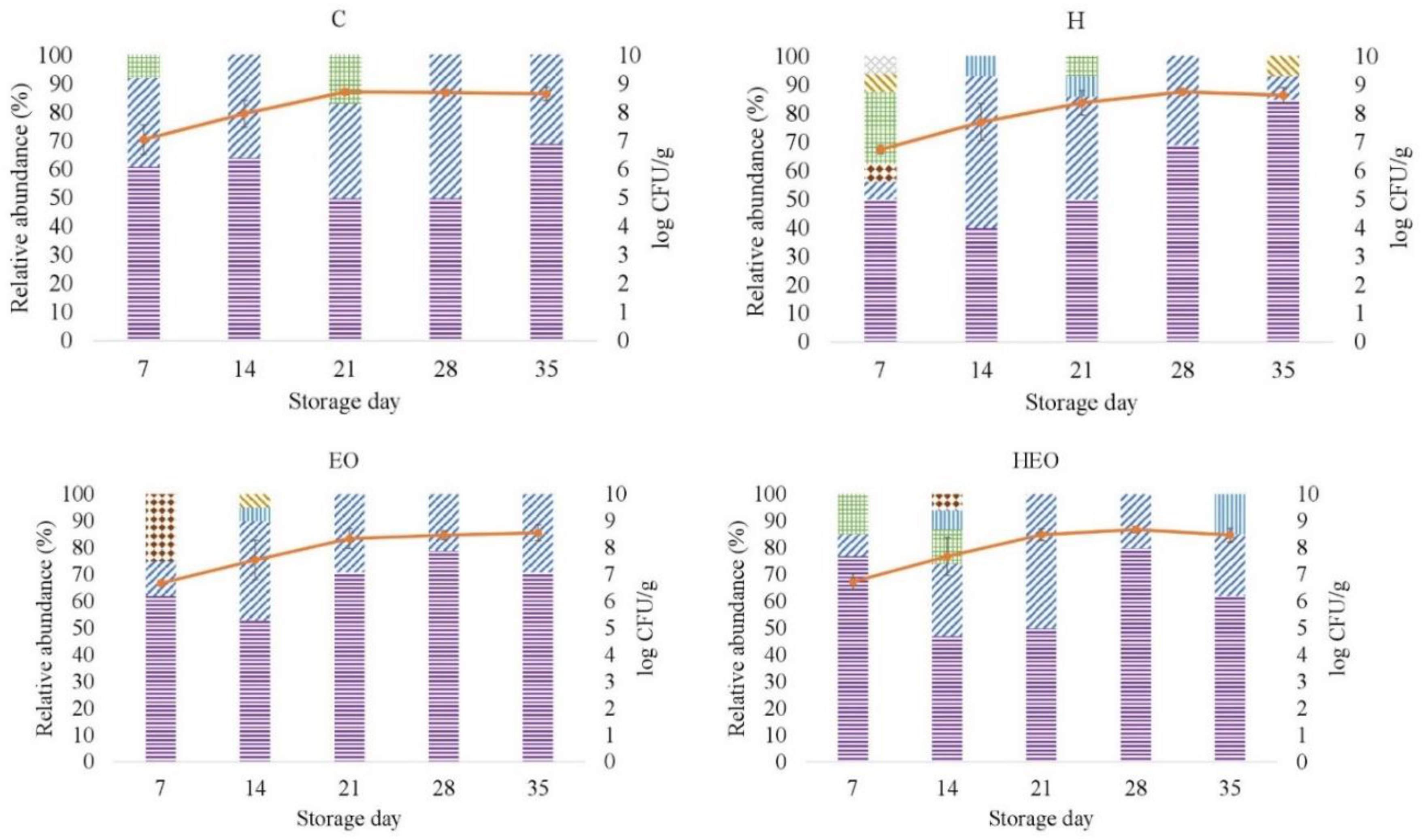{kind=link}
| Biogenic Amine | Treat | Time | RMSE | P-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | H | EO | HEO | 0 | 7 | 14 | 21 | 28 | 35 | Treat | Time | Treat × Time | ||
| Monoamines | ||||||||||||||
| Tryptamine | 17.66 | 20.48 | 17.52 | 19.08 | 5.58 b | 6.28b | 16.85 ab | 18.80 ab | 29.61 a | 34.99 a | 16.33 | 0.942 | 0.000 | 1.000 |
| Histamine | 6.90 | 7.58 | 4.53 | 8.24 | 0.53 b | 0.37 b | 2.40 b | 4.31 b | 13.96 ab | 19.29 a | 17.48 | 0.926 | 0.046 | 1.000 |
| Tyramine | 143.04 | 152.28 | 150.89 | 150.40 | 19.88 e | 99.04 d | 138.37 cd | 176.22 bc | 206.08 ab | 255.31 a | 44.25 | 0.924 | 0.000 | 1.000 |
| Diamines | ||||||||||||||
| Putrescine | 8.36 | 6.97 | 6.04 | 6.99 | 1.36 b | 1.86 b | 3.06 ab | 7.03 ab | 12.23 ab | 17.00 a | 12.35 | 0.955 | 0.015 | 1.000 |
| Cadaverine | 98.03 | 121.74 | 126.37 | 114.29 | 2.14 c | 14.33 c | 66.69 c | 148.73 b | 203.61 ab | 255.14 a | 54.56 | 0.432 | 0.000 | 0.996 |
| Poliamines | ||||||||||||||
| Spermine | 25.52 b | 28.88 ab | 29.22 ab | 29.89 a | 30.12 | 30.17 | 26.57 | 26.88 | 27.16 | 29.34 | 4.72 | 0.037 | 0.200 | 0.506 |
| Spermidine | 5.83 | 5.80 | 6.06 | 6.22 | 5.56 | 6.02 | 5.97 | 6.06 | 5.87 | 6.37 | 1.24 | 0.715 | 0.738 | 0.587 |
| Species | 0 (n = 33) | 7 (n = 50) | 14 (n = 63) | 21 (n = 52) | 28 (n = 56) | 35 (n = 53) |
|---|---|---|---|---|---|---|
| Aerococcus viridans | 12 | - | - | - | - | - |
| Carnobacterium maltraromaticum | - | - | 5 | 2 | - | 4 |
| Lactobacillus casei | 3 | - | - | - | - | - |
| Lactobacillus curvatus | - | 2 | 2 | - | - | 2 |
| Lactobacillus sakei | 33 | 62 | 51 | 56 | 70 | 72 |
| Lactobacillus spp. | 12 | 14 | 38 | 37 | 30 | 23 |
| Lactococcus lactis | 24 | 2 | - | - | - | - |
| Leuconostoc mesenteroides | 12 | 14 | 3 | 6 | - | - |
| Enterococcus faecalis | - | 6 | 2 | - | - | - |
| Streptococcus salivarius | 3 | - | - | - | - | - |
| Ingredients | Treatments | |||
|---|---|---|---|---|
| C | H | EO | HEO | |
| Lamb meat | 980 | 980 | 980 | 980 |
| Salt | 20 | 20 | 20 | 20 |
| Sodium bicarbonate | 3 | 3 | 3 | 3 |
| Spice infusion a | 20 | 20 | 20 | 20 |
| Water | 30 | - | 30 | 15 |
| Hops extract | - | 30 | - | 15 |
| Essential oil | - | - | 1 | 0.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carballo, D.E.; Mateo, J.; Andrés, S.; Giráldez, F.J.; Quinto, E.J.; Khanjari, A.; Operta, S.; Caro, I. Microbial Growth and Biogenic Amine Production in a Balkan-Style Fresh Sausage during Refrigerated Storage under a CO2-Containing Anaerobic Atmosphere: Effect of the Addition of Zataria multiflora Essential Oil and Hops Extract. Antibiotics 2019, 8, 227. https://doi.org/10.3390/antibiotics8040227
Carballo DE, Mateo J, Andrés S, Giráldez FJ, Quinto EJ, Khanjari A, Operta S, Caro I. Microbial Growth and Biogenic Amine Production in a Balkan-Style Fresh Sausage during Refrigerated Storage under a CO2-Containing Anaerobic Atmosphere: Effect of the Addition of Zataria multiflora Essential Oil and Hops Extract. Antibiotics. 2019; 8(4):227. https://doi.org/10.3390/antibiotics8040227
Chicago/Turabian StyleCarballo, Diego E., Javier Mateo, Sonia Andrés, Francisco Javier Giráldez, Emiliano J. Quinto, Ali Khanjari, Sabina Operta, and Irma Caro. 2019. "Microbial Growth and Biogenic Amine Production in a Balkan-Style Fresh Sausage during Refrigerated Storage under a CO2-Containing Anaerobic Atmosphere: Effect of the Addition of Zataria multiflora Essential Oil and Hops Extract" Antibiotics 8, no. 4: 227. https://doi.org/10.3390/antibiotics8040227
APA StyleCarballo, D. E., Mateo, J., Andrés, S., Giráldez, F. J., Quinto, E. J., Khanjari, A., Operta, S., & Caro, I. (2019). Microbial Growth and Biogenic Amine Production in a Balkan-Style Fresh Sausage during Refrigerated Storage under a CO2-Containing Anaerobic Atmosphere: Effect of the Addition of Zataria multiflora Essential Oil and Hops Extract. Antibiotics, 8(4), 227. https://doi.org/10.3390/antibiotics8040227