
Glyphosate Biodegradation by Airborne Plant Growth-Promoting Bacteria: Influence on Soil Microbiome Dynamics

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Site and Soil Characteristics
2.2. PGPB Strains
2.3. Airborne PGPB Cultivation
2.4. Experimental Set-Up
2.5. DNA Extraction and Metagenomic Analysis
2.6. Library Preparation and Sequencing
2.7. Bioinformatics with Spaghetti Pipeline
2.8. Glyphosate and AMPA Extraction
2.9. Glyphosate and AMPA Quantification by UPLC-MS/MS
2.10. Statistical and Biostatistical Analysis
2.11. Network Analysis
- Vertex (nodes): represent OTUs (operational taxonomic units) in the network.
- Edge (edges): represent correlations between OTUs, indicating potential interactions or associations.
- Average Degree: The average number of edges (connections) per vertex (OTU). This metric indicates the network’s general connectivity.
- Average Path Length: the average shortest path between all pairs of vertices in the network, reflecting how closely connected the network is.
- Network Diameter: the longest shortest path, representing the maximum distance between two vertices.
- Clustering Coefficient: A measure of the degree to which vertices in the network tend to cluster together. It reflects the presence of tightly knit groups within the network.
- Density: the ratio of the number of edges present to the number of possible edges in the network, indicating how densely the network is connected.
- Heterogeneity: It is the variability in vertex connectivity. A higher heterogeneity suggests a more varied distribution of connections among the vertices.
- Centralization: The extent to which the network is organized around central nodes, with a higher centralization indicating a more star-like structure.
- Modularity: The degree to which the network is divided into modules or subcommunities. High modularity suggests a structure with distinct clusters of highly interconnected nodes.
2.12. Redundancy Analysis (RDA)
- Variance filtering: the “zv” (zero variance) and “nzv” (near-zero variance) methods were used to remove variables with little or no variation, ensuring that only informative variables were retained.
- Normalization and transformation: the Yeo–Johnson transformation was applied to stabilize variance and approximate normality.
- Handling missing data: missing values were removed using “na.omit” to ensure that only complete cases were analyzed.
3. Results
3.1. Glyphosate Degradation
3.2. Microbial Composition and Structure
3.2.1. Clusters and Temporal Dynamics in Phylogenetic Composition
3.2.2. Temporal Dynamics and Responses of Microbial Communities to Glyphosate Treatments
3.2.3. Shifts in Soil Bacterial Structure
3.2.4. Network Configuration and Interaction Dynamics
3.2.5. Analysis of Predicted Functionality of Profile Gene
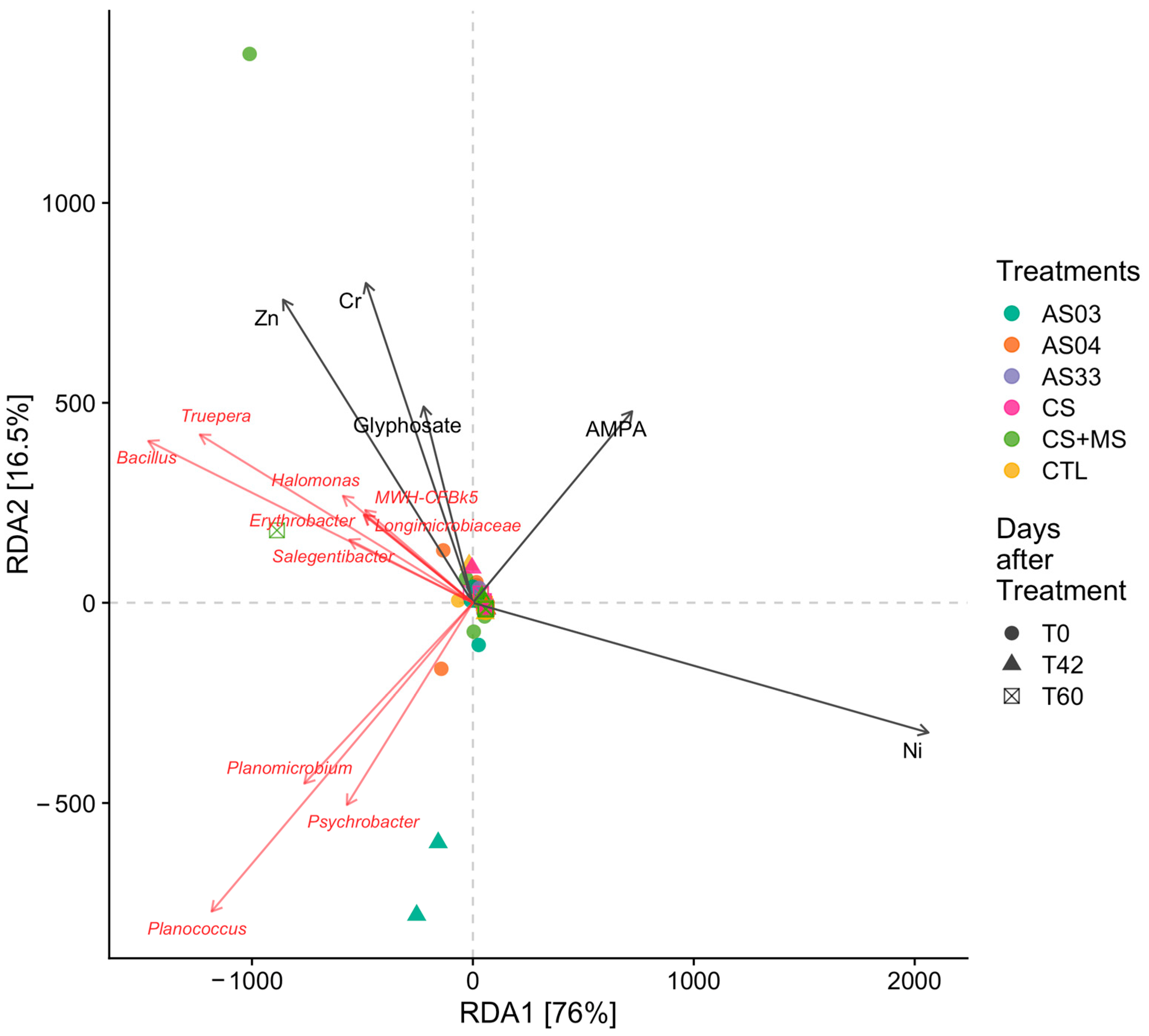
3.2.6. Correlation Between Environmental Variables and Soil Community
4. Discussion
4.1. Overview of Glyphosate Degradation
4.2. Soil Microbial Composition and Structure Response to PGPB Isolates on Glyphosate-Contaminated Soil
4.3. Temporal Dynamics of the Main Soil Bacterial Genera After Glyphosate Application
4.4. Impact of Bacterial Isolates and Consortia on Network Complexity
4.5. Positive and Negative Interactions: Ecological Implications
4.6. Network Modularity and Environmental Resilience
4.7. Predicted Functionality of Profile Gene
5. Conclusions
- −
- Microbial Community Dynamics: The introduction of PGPB significantly altered the soil microbial structure, remarkably increasing the abundance of Proteobacteria and Firmicutes, essential for glyphosate biodegradation. These changes enhance the functional capacity of soil microbiota, potentially leading to sustained soil health and fertility.
- −
- Bioaugmentation Efficacy: Although the consortium of PGPB strains showed a rapid initial decrease in glyphosate levels, Exiguobacterium indicum AS03 maintained superior degradation efficiency over time. These results highlight the effectiveness of specific strains over the consortia for long-term bioremediation.
- −
- Ecological Impacts: Significant shifts in microbial community composition and structure in response to glyphosate exposure and PGPB treatment illustrate the adaptability of the soil microbiomes. These shifts indicate soil resilience and are crucial for developing effective bioremediation strategies that support agricultural sustainability and environmental health.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO; UNEP. Global Assessment of Soil Pollution–Summary for Policy Makers; FAO and UNEP: Rome, Italy, 2021. [Google Scholar]
- FAO. Pesticides Use and Trade, 1990–2021; FAO: Rome, Italy, 2023. [Google Scholar]
- FAOSTAT Statistical Database Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/faostat/en/ (accessed on 25 September 2024).
- Beaumelle, L.; Thouvenot, L.; Hines, J.; Jochum, M.; Eisenhauer, N.; Phillips, H.R.P. Soil Fauna Diversity and Chemical Stressors: A Review of Knowledge Gaps and Roadmap for Future Research. Ecography 2021, 44, 845–859. [Google Scholar] [CrossRef]
- Meftaul, I.M.; Venkateswarlu, K.; Annamalai, P.; Parven, A.; Megharaj, M. Glyphosate Use in Urban Landscape Soils: Fate, Distribution, and Potential Human and Environmental Health Risks. J. Environ. Manag. 2021, 292, 112786. [Google Scholar] [CrossRef]
- Domínguez, A.; Brown, G.G.; Sautter, K.D.; Ribas de Oliveira, C.M.; de Vasconcelos, E.C.; Niva, C.C.; Bartz, M.L.C.; Bedano, J.C. Toxicity of AMPA to the Earthworm Eisenia Andrei Bouché, 1972 in Tropical Artificial Soil. Sci. Rep. 2016, 6, 19731. [Google Scholar] [CrossRef] [PubMed]
- FAO. Pesticides Use and Trade, 1990–2022; FAO: Rome, Italy, 2024. [Google Scholar] [CrossRef]
- Raimondo, E.E.; Saez, J.M.; Aparicio, J.D.; Fuentes, M.S.; Benimeli, C.S. Coupling of Bioaugmentation and Biostimulation to Improve Lindane Removal from Different Soil Types. Chemosphere 2020, 238, 124512. [Google Scholar] [CrossRef]
- Panwar, R.; Mathur, J. Remediation of Polycyclic Aromatic Hydrocarbon-Contaminated Soils Using Microbes and Nanoparticles: A Review. Pedosphere 2023, 33, 93–104. [Google Scholar] [CrossRef]
- Rodríguez-Vázquez, R.; Mesa-Marín, J. Plant Responses to Plant Growth Promoting Bacteria: Insights from Proteomics. J. Plant Physiol. 2023, 287, 154031. [Google Scholar] [CrossRef]
- Guardado-Fierros, B.G.; Tuesta-Popolizio, D.A.; Lorenzo-Santiago, M.A.; Rubio-Cortés, R.; Camacho-Ruíz, R.M.; Castañeda-Nava, J.J.; Gutiérrez-Mora, A.; Contreras-Ramos, S.M. PGPB Consortium Formulation to Increase Fermentable Sugar in Agave Tequilana Weber Var. Blue: A Study in the Field. Plants 2024, 13, 1371. [Google Scholar] [CrossRef]
- Martínez-Rabelo, F.; Gómez-Guzmán, L.A.; García-Segura, D.R.; Villegas-García, E.; Rodriguez-Campos, J.; Velázquez-Fernández, J.B.; Hernández-Castellanos, B.; Barois, I.; Contreras-Ramos, S.M. Hydrocarbon Bioremediation in a Pilot-Scale: A Combination of Bioaugmentation, Phytoremediation, and Vermiremediation. Environ. Technol. Innov. 2023, 31, 103210. [Google Scholar] [CrossRef]
- Rani, R.; Kumar, V.; Gupta, P.; Chandra, A. Potential Use of Solanum Lycopersicum and Plant Growth Promoting Rhizobacterial (PGPR) Strains for the Phytoremediation of Endosulfan Stressed Soil. Chemosphere 2021, 279, 130589. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, W.J.; Chen, S.F.; Liu, M.; Ghorab, M.A.; Mishra, S.; Bhatt, P.; Chen, S. Complete Biodegradation of Glyphosate with Microbial Consortium YS622: Structural Analysis, Biochemical Pathways, and Environmental Bioremediation. J. Environ. Chem. Eng. 2024, 12, 114344. [Google Scholar] [CrossRef]
- Ezaka, E.; Akintokun, A.K.; Akintokun, P.O.; Taiwo, L.B.; Uthman, A.C.O.; Oyedele, O.A.; Aluko, O.I. Glyphosate Degradation by Two Plant Growth Promoting Bacteria (PGPB) Isolated from Rhizosphere of Maize. Microbiol. Res. J. Int. 2018, 26, 1–11. [Google Scholar] [CrossRef]
- Masotti, F.; Garavaglia, B.S.; Gottig, N.; Ottado, J. Bioremediation of the Herbicide Glyphosate in Polluted Soils by Plant-Associated Microbes. Curr. Opin. Microbiol. 2023, 73, 102290. [Google Scholar] [CrossRef] [PubMed]
- Villegas, L.B.; Martínez, M.A.; Rodríguez, A.; Amoroso, M.J. Microbial Consortia, a Viable Alternative for Cleanup of Contaminated Soils. In Bioremediation in Latin America; Springer International Publishing: Cham, Switzerland, 2014; pp. 135–148. [Google Scholar]
- Singh, S.; Kumar, V.; Gill, J.P.K.; Datta, S.; Singh, S.; Dhaka, V.; Kapoor, D.; Wani, A.B.; Dhanjal, D.S.; Kumar, M.; et al. Herbicide Glyphosate: Toxicity and Microbial Degradation. Int. J. Environ. Res. Public Health 2020, 17, 7519. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, W.J.; Chen, S.F.; Lei, Q.; Li, J.; Bhatt, P.; Mishra, S.; Chen, S. Cellular Response and Molecular Mechanism of Glyphosate Degradation by Chryseobacterium sp. Y16C. J. Agric. Food Chem. 2023, 71, 6650–6661. [Google Scholar] [CrossRef]
- Sanmartin Negrete, P.; Ghilardi, C.; Rodriguez Pineda, L.; Perez, E.; Herrera, M.L.; Borroni, V. Biosurfactant Production by Rhodococcus ALDO1 Isolated from Olive Mill Wastes. Biocatal. Agric. Biotechnol. 2024, 57, 103106. [Google Scholar] [CrossRef]
- Mohy-Ud-Din, W.; Akhtar, M.J.; Bashir, S.; Asghar, H.N.; Nawaz, M.F.; Chen, F. Isolation of Glyphosate-Resistant Bacterial Strains to Improve the Growth of Maize and Degrade Glyphosate under Axenic Condition. Agriculture 2023, 13, 886. [Google Scholar] [CrossRef]
- Guardado-Fierros, B.G.; Tuesta-Popolizio, D.A.; Lorenzo-Santiago, M.A.; Rodriguez-Campos, J.; Contreras-Ramos, S.M. Comparative Study between Salkowski Reagent and Chromatographic Method for Auxins Quantification from Bacterial Production. Front. Plant Sci. 2024, 15, 1378079. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, M.; Paramasivan, M. Plant Growth-Promoting Bacterial (PGPB) Mediated Degradation of Hazardous Pesticides: A Review. Int. Biodeterior. Biodegrad. 2024, 190, 105769. [Google Scholar] [CrossRef]
- Sajjad, B.; Hussain, S.; Rasool, K.; Hassan, M.; Almomani, F. Comprehensive Insights into Advances in Ambient Bioaerosols Sampling, Analysis and Factors Influencing Bioaerosols Composition. Environ. Pollut. 2023, 336, 122473. [Google Scholar] [CrossRef]
- Jahne, M.A.; Rogers, S.W.; Holsen, T.M.; Grimberg, S.J.; Ramler, I.P.; Kim, S. Bioaerosol Deposition to Food Crops near Manure Application: Quantitative Microbial Risk Assessment. J. Environ. Qual. 2016, 45, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Ronan, E.; Yeung, C.W.; Hausner, M.; Wolfaardt, G.M. Interspecies Interaction Extends Bacterial Survival at Solid–Air Interfaces. Biofouling 2013, 29, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Gee, G.W.; Bauder, J.W. Particle Size Analysis. In Methods of Soil Analysis. Vol. I Physical and Mineralogical Methods; Klute, A., Ed.; American Society of Agronomy: Madison, WI, USA, 1986; Volume I, pp. 383–411. [Google Scholar]
- Thomas, G.W. Soil PH and Soil Acidity. In Methods of Soil Analysis, Part. 3: Chemical Methods; Wiley: Hoboken, NJ, USA, 2018; pp. 475–490. [Google Scholar] [CrossRef]
- Amato, M. Determination of Carbon 12C and 14C in Plant and Soil. Soil Biol. Biochem. 1983, 15, 611–612. [Google Scholar] [CrossRef]
- Bremner, J.M.; Maktar, H.P.S. Nitrogen Total. In Methods of Soil Analysis: Chemical Methods Part 3. Soil Science Society of America Inc; Sparks, D.L., Ed.; American Society of Agronomy: Madison, WI, USA, 1996; p. 1085. [Google Scholar]
- NOM-021-SEMARNAT-2000; Que Establece las Especificaciones de Fertilidad, Salinidad y Clasificación de Suelos. Estudios, Muestreo y Análisis. Norma Oficial Mexicana: Naulcalpan, México, 2002. Available online: https://platiica.economia.gob.mx/normalizacion/nom-021-semarnat-2000/ (accessed on 25 September 2024).
- Sparks, D.L. Methods of Soil Analysis. Part 3. Chemical Methods; Soil Science Society of America Book Series; CRC Press: Boca Raton, FL, USA, 1996; Volume 3, p. 1264. [Google Scholar]
- Hazaimeh, M.; Kanaan, B.M.; AlFaleh, F.A.; Elhaig, M.M.; Khamaiseh, E.I.; Zia, Q.; Alaidarous, M.; Seth, C.S.; Alsowayeh, N.; Ahmad, F. Biodegradation of Petroleum Hydrocarbons Using a Novel Bacterial Strain Isolated from Hydrocarbons Contaminated Soil of Saudi Arabia. Biocatal. Agric. Biotechnol. 2024, 57, 103074. [Google Scholar] [CrossRef]
- Goswami, D.; Dhandhukia, P.; Patel, P.; Thakker, J.N. Screening of PGPR from Saline Desert of Kutch: Growth Promotion in Arachis Hypogea by Bacillus licheniformis A2. Microbiol. Res. 2014, 169, 66–75. [Google Scholar] [CrossRef] [PubMed]
- APHA/AWWA/WEF. Standard Methods for the Examination of Water and Wastewater; Standard Methods; APHA/AWWA/WEF: Washington, DC, USA, 2012; 541p, ISBN 9780875532356. Available online: https://www.standardmethods.org/doi/book/10.2105/SMWW.2882 (accessed on 21 September 2024).
- Rajawat, M.V.S.; Singh, S.; Tyagi, S.P.; Saxena, A.K. A Modified Plate Assay for Rapid Screening of Potassium-Solubilizing Bacteria. Pedosphere 2016, 26, 768–773. [Google Scholar] [CrossRef]
- Alexander, D.B.; Zuberer, D.A. Use of Chrome Azurol S Reagents to Evaluate Siderophore Production by Rhizosphere Bacteria. Biol. Fertil. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Afanador-Barajas, L.N.; Navarro-Noya, Y.E.; Luna-Guido, M.L.; Dendooven, L. Impact of a Bacterial Consortium on the Soil Bacterial Community Structure and Maize (Zea mays L.) Cultivation. Sci. Rep. 2021, 11, 13092. [Google Scholar] [CrossRef]
- Latorre-Pérez, A.; Gimeno-Valero, H.; Tanner, K.; Pascual, J.; Vilanova, C.; Porcar, M. A Round Trip to the Desert: In Situ Nanopore Sequencing Informs Targeted Bioprospecting. Front. Microbiol. 2021, 12, 768240. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Completing Bacterial Genome Assemblies with Multiplex MinION Sequencing. Microb. Genom. 2017, 3, e000132. [Google Scholar] [CrossRef] [PubMed]
- De Coster, W.; D’Hert, S.; Schultz, D.T.; Cruts, M.; Van Broeckhoven, C. NanoPack: Visualizing and Processing Long-Read Sequencing Data. Bioinformatics 2018, 34, 2666–2669. [Google Scholar] [CrossRef]
- Marijon, P.; Chikhi, R.; Varré, J.S. Yacrd and Fpa: Upstream Tools for Long-Read Genome Assembly. Bioinformatics 2020, 36, 3894–3896. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Minimap2: Pairwise Alignment for Nucleotide Sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “All-Species Living Tree Project (LTP)” Taxonomic Frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [PubMed]
- Gamaarachchi, H.; Parameswaran, S.; Smith, M.A. Featherweight Long Read Alignment Using Partitioned Reference Indexes. Sci. Rep. 2019, 9, 4318. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- De Gerónimo, E.; Aparicio, V.C.; Costa, J.L. Glyphosate Sorption to Soils of Argentina. Estimation of Affinity Coefficient by Pedotransfer Function. Geoderma 2018, 322, 140–148. [Google Scholar] [CrossRef]
- Callahan, B.J.; Sankaran, K.; Fukuyama, J.A.; McMurdie, P.J.; Holmes, S.P. Bioconductor Workflow for Microbiome Data Analysis: From Raw Reads to Community Analyses. F1000Research 2016, 5, 1492. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package. 2022. Available online: https://cran.r-project.org/package=vegan (accessed on 21 September 2024).
- Liu, C.; Cui, Y.; Li, X.; Yao, M. Microeco: An R Package for Data Mining in Microbial Community Ecology. FEMS Microbiol. Ecol. 2021, 97, 255. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2; Springer: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. Circlize Implements and Enhances Circular Visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef]
- Auguie, B. GridExtra: Miscellaneous Functions for “Grid” Graphics 2017. Available online: https://cran.r-project.org/web/packages/gridExtra/gridExtra.pdf (accessed on 22 September 2024).
- Liang, W.; Wu, Y.; Ma, X. Robust Sparse Precision Matrix Estimation for High-Dimensional Compositional Data. Stat. Probab. Lett. 2022, 184, 109379. [Google Scholar] [CrossRef]
- Csárdi, G.; Nepusz, T.; Traag, V.; Horvát, S.; Zanini, F.; Noom, D.; Müller, K. Igraph: Network Analysis and Visualization in R. 2024. Available online: https://igraph.org/r/pdf/latest/igraph.pdf (accessed on 28 October 2024).
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. Proc. Int. AAAI Conf. Web Soc. Media 2009, 3, 361–362. [Google Scholar] [CrossRef]
- Manogaran, M.; Ahmad, S.A.; Yasid, N.A.; Yakasai, H.M.; Shukor, M.Y. Characterisation of the Simultaneous Molybdenum Reduction and Glyphosate Degradation by Burkholderia Vietnamiensis AQ5-12 and Burkholderia Sp. AQ5-13. 3 Biotech 2018, 8, 1–8. [Google Scholar] [CrossRef]
- Massot, F.; Gkorezis, P.; Van Hamme, J.; Marino, D.; Trifunovic, B.S.; Vukovic, G.; d’Haen, J.; Pintelon, I.; Giulietti, A.M.; Merini, L.; et al. Isolation, Biochemical and Genomic Characterization of Glyphosate Tolerant Bacteria to Perform Microbe-Assisted Phytoremediation. Front. Microbiol. 2021, 11, 598507. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, W.J.; Huang, Y.; Li, J.; Zhong, J.; Zhang, W.; Zou, Y.; Mishra, S.; Bhatt, P.; Chen, S. Insights into the Microbial Degradation and Resistance Mechanisms of Glyphosate. Environ. Res. 2022, 215, 114153. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.N.; Sachan, S.G.; Verma, P.; Kaushik, R.; Saxena, A.K. Cold Active Hydrolytic Enzymes Production by Psychrotrophic Bacilli Isolated from Three Sub-Glacial Lakes of NW Indian Himalayas. J. Basic Microbiol. 2016, 56, 294–307. [Google Scholar] [CrossRef]
- Singh, V.K.; Mishra, A.; Jha, B. 3-Benzyl-Hexahydro-Pyrrolo[1,2-a]Pyrazine-1,4-Dione Extracted from Exiguobacterium Indicum Showed Anti-Biofilm Activity against Pseudomonas Aeruginosa by Attenuating Quorum Sensing. Front. Microbiol. 2019, 10, 435900. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Vo, V.T.; Nguyen, T.H.P.; Kiefer, R. Isolation and Optimization of a Glyphosate-Degrading Rhodococcus Soli G41 for Bioremediation. Arch. Microbiol. 2022, 204, 252. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, A.Y.Z. Degradation of Diesel-Oil by a Newly Isolated Kocuria sediminis DDK6. Afr. J. Microbiol. Res. 2017, 11, 400–407. [Google Scholar] [CrossRef]
- Lalević, B.T.; Jović, J.B.; Raičević, V.B.; Kljujev, I.S.; Kiković, D.D.; Hamidović, S.R. Biodegradation of Methyl-Tert-Butyl Ether by Kocuria sp. Hem. Ind. 2012, 66, 717–722. [Google Scholar] [CrossRef]
- Castro, D.B.A.; Pereira, L.B.; e Silva, M.V.M.; da Silva, B.P.; Palermo, B.R.Z.; Carlos, C.; Belgini, D.R.B.; Limache, E.E.G.; Lacerda, G.V.J.; Nery, M.B.P.; et al. High-Quality Draft Genome Sequence of Kocuria marina SO9-6, an Actinobacterium Isolated from a Copper Mine. Genom. Data 2015, 5, 34–35. [Google Scholar] [CrossRef] [PubMed]
- Harshvardhan, K.; Jha, B. Biodegradation of Low-Density Polyethylene by Marine Bacteria from Pelagic Waters, Arabian Sea, India. Mar. Pollut. Bull. 2013, 77, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Soric, A.; Boutin, O. Treatment Technologies and Degradation Pathways of Glyphosate: A Critical Review. Sci. Total Environ. 2020, 742, 140559. [Google Scholar] [CrossRef] [PubMed]
- Battaglin, W.A.; Meyer, M.T.; Kuivila, K.M.; Dietze, J.E. Glyphosate and Its Degradation Product AMPA Occur Frequently and Widely in U.S. Soils, Surface Water, Groundwater, and Precipitation. JAWRA J. Am. Water Resour. Assoc. 2014, 50, 275–290. [Google Scholar] [CrossRef]
- Aslam, S.; Arslan, M.; Nowak, K.M. Microbial Activity, Community Composition and Degraders in the Glyphosate-Spiked Soil Are Driven by Glycine Formation. Sci. Total Environ. 2024, 907, 168206. [Google Scholar] [CrossRef]
- Wilms, W.; Parus, A.; Homa, J.; Batycka, M.; Niemczak, M.; Woźniak-Karczewska, M.; Trzebny, A.; Zembrzuska; Dabert, M.; Táncsics, A.; et al. Glyphosate versus Glyphosate Based Ionic Liquids: Effect of Cation on Glyphosate Biodegradation, SoxA and PhnJ Genes Abundance and Microbial Populations Changes during Soil Bioaugmentation. Chemosphere 2023, 316, 137717. [Google Scholar] [CrossRef] [PubMed]
- Pérez, G.L.; Torremorell, A.; Mugni, H.; Rodríguez, P.; Solange Vera, M.; Do Nascimento, M.; Allende, L.; Bustingorry, J.; Escaray, R.; Ferraro, M.; et al. Effects of the herbicide Roundup on freshwater microbial communities: A mesocosm study. Ecol. Appl. 2007, 17, 2310–2322. [Google Scholar] [CrossRef]
- Guo, B.; Guo, Y.; Hong, H.; Jin, L.; Zhang, L.; Chang, R.Z.; Lu, W.; Lin, M.; Qiu, L.J. Co-Expression of G2-EPSPS and Glyphosate Acetyltransferase GAT Genes Conferring High Tolerance to Glyphosate in Soybean. Front. Plant Sci. 2015, 6, 138289. [Google Scholar] [CrossRef]
- Castrejón-Godínez, M.L.; Tovar-Sánchez, E.; Valencia-Cuevas, L.; Rosas-Ramírez, M.E.; Rodríguez, A.; Mussali-Galante, P. Glyphosate Pollution Treatment and Microbial Degradation Alternatives, a Review. Microorganisms 2021, 9, 2322. [Google Scholar] [CrossRef]
- Elarabi, N.I.; Abdelhadi, A.A.; Ahmed, R.H.; Saleh, I.; Arif, I.A.; Osman, G.; Ahmed, D.S. Bacillus Aryabhattai FACU: A Promising Bacterial Strain Capable of Manipulate the Glyphosate Herbicide Residues. Saudi J. Biol. Sci. 2020, 27, 2207–2214. [Google Scholar] [CrossRef] [PubMed]
- Langarica-Fuentes, A.; Straub, D.; Wimmer, B.; Thompson, K.; Nahnsen, S.; Huhn, C.; Kleindienst, S. Subtle Microbial Community Changes despite Rapid Glyphosate Degradation in Microcosms with Four German Agricultural Soils. Appl. Soil Ecol. 2024, 198, 105381. [Google Scholar] [CrossRef]
- Kanissery, R.; Gairhe, B.; Kadyampakeni, D.; Batuman, O.; Alferez, F. Glyphosate: Its Environmental Persistence and Impact on Crop Health and Nutrition. Plants 2019, 8, 499. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Khan, M.H.; Yuan, Z.; Hussain, S.; Cao, H.; Liu, Y. Response of Soil Microbiome Structure and Its Network Profiles to Four Soil Amendments in Monocropping Strawberry Greenhouse. PLoS ONE 2021, 16, e0245180. [Google Scholar] [CrossRef] [PubMed]
- de Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S.; Hallin, S.; Kaisermann, A.; Keith, A.M.; Kretzschmar, M.; et al. Soil Bacterial Networks Are Less Stable under Drought than Fungal Networks. Nat. Commun. 2018, 9, 3033. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.; Ma, G.; Chen, S.; Lin, C.; Zhao, X. Microbial Network and Soil Properties Are Changed in Bacterial Wilt-Susceptible Soil. Appl. Environ. Microbiol. 2019, 85, 00162-19. [Google Scholar] [CrossRef] [PubMed]
- Wagg, C.; Dudenhöffer, J.H.; Widmer, F.; van der Heijden, M.G.A. Linking Diversity, Synchrony and Stability in Soil Microbial Communities. Funct. Ecol. 2018, 32, 1280–1292. [Google Scholar] [CrossRef]
- Kavya, Y.; Trimurtulu, N.; Gopal, A.V.; Vani, P.M.; Prasad, N.V.V.S.D. Effect of Inoculation of Microbial Consortia on Soil Physicochemical and Nutrient Status. Curr. J. Appl. Sci. Technol. 2020, 39, 1–8. [Google Scholar] [CrossRef]
- Moliterni, E.; Jiménez-Tusset, R.G.; Villar Rayo, M.; Rodriguez, L.; Fernández, F.J.; Villaseñor, J. Kinetics of Biodegradation of Diesel Fuel by Enriched Microbial Consortia from Polluted Soils. Int. J. Environ. Sci. Technol. 2012, 9, 749–758. [Google Scholar] [CrossRef]
- Corstanje, R.; Deeks, L.R.; Whitmore, A.P.; Gregory, A.S.; Ritz, K. Probing the Basis of Soil Resilience. Soil Use Manag. 2015, 31, 72–81. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Caruso, T. Soil Microbial Community Responses to Climate Extremes: Resistance, Resilience and Transitions to Alternative States. Philos. Trans. R. Soc. B 2020, 375, 20190112. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Deng, Y.; Luo, F.; He, Z.; Tu, Q.; Zhi, X. Functional Molecular Ecological Networks. mBio 2010, 1, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Haney, R.; Senseman, S.; Hons, F.; Zuberer, D. Effect of Glyphosate on Soil Microbial Activity and Biomass on JSTOR. Available online: https://www.jstor.org/stable/4046161 (accessed on 15 May 2024).
- Das, S.; Kim, G.W.; Hwang, H.Y.; Verma, P.P.; Kim, P.J. Cropping with Slag to Address Soil, Environment, and Food Security. Front. Microbiol. 2019, 10, 440580. [Google Scholar] [CrossRef]
- Siles, J.A.; García-Sánchez, M.; Gómez-Brandón, M. Studying Microbial Communities through Co-Occurrence Network Analyses during Processes of Waste Treatment and in Organically Amended Soils: A Review. Microorganisms 2021, 9, 1165. [Google Scholar] [CrossRef] [PubMed]
- Ohtonen, R.; Fritze, H.; Pennanen, T.; Jumpponen, A.; Trappe, J. Ecosystem Properties and Microbial Community Changes in Primary Succession on a Glacier Forefront. Oecologia 1999, 119, 239–246. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, M.; Zhao, Y.; Hu, N.; Qin, L.; Ren, Z.; Wang, G.; Jiang, M. Soil Microbial Abundance Was More Affected by Soil Depth than the Altitude in Peatlands. Front. Microbiol. 2022, 13, 1068540. [Google Scholar] [CrossRef]
- Tan, L.; Gu, S.; Li, S.; Ren, Z.; Deng, Y.; Liu, Z.; Gong, Z.; Xiao, W.; Hu, Q. Responses of Microbial Communities and Interaction Networks to Different Management Practices in Tea Plantation Soils. Sustainability 2019, 11, 4428. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, L.; Liu, Z.; Chen, J.; Song, H.; Wang, J.; Cui, H.; Yang, Z.; Xiao, S.; Liu, K.; et al. Microbial Interactions Play an Important Role in Regulating the Effects of Plant Species on Soil Bacterial Diversity. Front. Microbiol. 2022, 13, 984200. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Wang, J.; Huang, X.; Tang, Z.; Liu, S.; Sun, O.J. Relating Microbial Community Structure to Functioning in Forest Soil Organic Carbon Transformation and Turnover. Ecol. Evol. 2014, 4, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Huang, W.; Guo, C.; Wang, R.; Xiao, C. Soil Microbial Properties and Plant Growth Responses to Carbon and Water Addition in a Temperate Steppe: The Importance of Nutrient Availability. PLoS ONE 2012, 7, e35165. [Google Scholar] [CrossRef] [PubMed]
- Song, H.S.; Renslow, R.S.; Fredrickson, J.K.; Lindemann, S.R. Integrating Ecological and Engineering Concepts of Resilience in Microbial Communities. Front. Microbiol. 2015, 6, 164175. [Google Scholar] [CrossRef]
- Zhong, W.; Gu, T.; Wang, W.; Zhang, B.; Lin, X.; Huang, Q.; Shen, W. The Effects of Mineral Fertilizer and Organic Manure on Soil Microbial Community and Diversity. Plant Soil 2010, 326, 511–522. [Google Scholar] [CrossRef]
- Agyekum, D.V.A.; Kobayashi, T.; Dastogeer, K.M.G.; Yasuda, M.; Sarkodee-Addo, E.; Ratu, S.T.N.; Xu, Q.; Miki, T.; Matsuura, E.; Okazaki, S. Diversity and Function of Soybean Rhizosphere Microbiome under Nature Farming. Front. Microbiol. 2023, 14, 1130969. [Google Scholar] [CrossRef] [PubMed]
- Ziesack, M.; Gibson, T.; Oliver, J.K.W.; Shumaker, A.M.; Hsu, B.B.; Riglar, D.T.; Giessen, T.W.; DiBenedetto, N.V.; Bry, L.; Way, J.C.; et al. Engineered Interspecies Amino Acid Cross-Feeding Increases Population Evenness in a Synthetic Bacterial Consortium. mSystems 2019, 4, e00352-19. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.; Wang, J.T.; Deng, Y.; He, J.Z.; Feng, K.; Zhang, L.M. Microbial Community and Functional Structure Significantly Varied among Distinct Types of Paddy Soils but Responded Differently along Gradients of Soil Depth Layers. Front. Microbiol. 2017, 8, 260931. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Liu, J.; Wang, J.; Han, W.; Shen, Z.; Muraina, T.O.; Chen, J.; Sun, D. Comparison of Soil Microbial Community between Reseeding Grassland and Natural Grassland in Songnen Meadow. Sci. Rep. 2020, 10, 16884. [Google Scholar] [CrossRef]
- Cuartero, J.; Özbolat, O.; Sánchez-Navarro, V.; Weiss, J.; Zornoza, R.; Pascual, J.A.; Vivo, J.M.; Ros, M. Long-Term Compost Amendment Changes Interactions and Specialization in the Soil Bacterial Community, Increasing the Presence of Beneficial N-Cycling Genes in the Soil. Agronomy 2022, 12, 316. [Google Scholar] [CrossRef]
- Kulikova, N.A.; Zhelezova, A.D.; Filippova, O.I.; Plyushchenko, I.V.; Rodin, I.A. The Degradation of Glyphosate and Its Effect on the Microbial Community of Agro-Sod–Podzolic Soil under Short-Term Model Experiment Conditions. Mosc. Univ. Soil Sci. Bull. 2020, 75, 138–145. [Google Scholar] [CrossRef]
- Liao, X.; Zhao, J.; Yi, Q.; Li, J.; Li, Z.; Wu, S.; Zhang, W.; Wang, K. Metagenomic Insights into the Effects of Organic and Inorganic Agricultural Managements on Soil Phosphorus Cycling. Agric. Ecosyst. Environ. 2023, 343, 108281. [Google Scholar] [CrossRef]
- Ferdush, J.; Paul, V. A Review on the Possible Factors Influencing Soil Inorganic Carbon under Elevated CO2. Catena 2021, 204, 105434. [Google Scholar] [CrossRef]
- Zhang, B.; Gao, R.; Dong, X. The Seasonal Impact of Thinning Intensities on Soil Carbon Cycling in the Lesser Xing’an Range, Northeast China. Forests 2024, 15, 449. [Google Scholar] [CrossRef]
- Johnson, E.T.; Bowman, M.J.; Gomes, R.P.; Carneiro, L.C.; Dunlap, C.A. Identification of 2,4-Diacetylphloroglucinol Production in the Genus Chromobacterium. Sci. Rep. 2023, 13, 14292. [Google Scholar] [CrossRef] [PubMed]
- Castro Tapia, M.P.; Madariaga Burrows, R.P.; Ruiz Sepúlveda, B.; Vargas Concha, M.; Vera Palma, C.; Moya-Elizondo, E.A. Antagonistic Activity of Chilean Strains of Pseudomonas Protegens Against Fungi Causing Crown and Root Rot of Wheat (Triticum aestivum L.). Front. Plant Sci. 2020, 11, 540430. [Google Scholar] [CrossRef]
- Devi, U.A.; Srinivas, T.; Parvathi, D.; Venkateshwarlu, M.; Ugandhar, T. The Impact of Innovative Research Methods for Enhancing Agricultural Plants for Sustainable Development in The Future. Int. Res. J. Adv. Eng. Manag. (IRJAEM) 2024, 2, 65–73. [Google Scholar] [CrossRef]
- Aguilar-Paredes, A.; Valdés, G.; Araneda, N.; Valdebenito, E.; Hansen, F.; Nuti, M. Microbial Community in the Composting Process and Its Positive Impact on the Soil Biota in Sustainable Agriculture. Agronomy 2023, 13, 542. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Soil a |
|---|---|
| Total Phosphorous (TP) (mg kg−1) | 1253.3 ± 35.9 |
| Total Nitrogen (TN) (mg kg−1) | 2443.4 ± 148.0 |
| Total Organic Carbon (TOC) (%) | 7.8 ± 0.1 |
| MO % | 13.5 ± 0.2 |
| pH | 7.4 ± 0.03 |
| Available Phosphorous (AP) (mg kg−1) | 828.0 ± 84.8 |
| CE mS cm−1 | 19.2 ± 4.4 |
| NH4+ (mg kg−1) | 20.8 ± 1.4 |
| NO2− (mg kg−1) | 0.5 ± 0.03 |
| NO3− (mg kg−1) | 48.3 ± 1.9 |
| Inorganic Nitrogen (mg kg−1) | 69.6 ± 1.0 |
| Sand (%) | 27.7 ± 1.2 |
| Silt (%) | 28.7 ± 3.1 |
| Clay (%) | 43.6 ± 4.0 |
| Texture | Clay |
| Cd (mg kg−1) | <0.25 ± 0.0 |
| Cr (mg kg−1) | 11.4 ± 0.1 |
| Zn (mg kg−1) | 43.9 ± 1.1 |
| Hg (mg kg−1) | <0.1 ± 0.0 |
| Ni (mg kg−1) | 4.1 ± 0.1 |
| Pb (mg kg−1) | 5.7 ± 3.3 |
| Code Strain | Bacteria Identification * | Glyphosate Concentration ** | PGPB Traits | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 40 | 60 | 100 | Phosphorous Solubilization a | Ammonium Production b | Potassium Solubilization c | Siderophores d | IAA Auxin e | |||
| With | Without | |||||||||
| Tryptophane | ||||||||||
| −−−−−−−−−−−−−−−−−−mg L−1−−−−−−−−−−−−−−−−− | −−−−−−−−−−−−S.I **−−−−−−−−−− | −−−−mg L−1−−−− | ||||||||
| As03 | Exiguobacterium indicum (1.75) | •• | •• | •• | 15.1 ± 1.02 a | 28.8 ± 9.1 a | 0 c | 0.5 ± 0.2 bc | 0.08 ± 0.01 b | 0.008 ±0.002 a |
| As04 | Kocuria sediminis (1.8) | •• | •• | •• | 13 ± 1.22 ab | 23.06 ± 0.15 ab | 8.4 ± 0.20 a | 0.63 ± 0.15 ab | 0.41 ± 0.30 b | 0.02 ± 0.003 a |
| As33 | Rhodococcus rhodochrous (2.17) | •• | •• | •• | 10.2 ± 2.1 ab | 31.1 ± 2.65 a | 6.6 ± 0.3 ab | 0.86 ± 0.3 ab | 86.61 ± 8.09 a | 0.002 ± 0.001 a |
| Code for Treatments | Soil Condition | Inoculation | Treatment Description (Responsible) |
|---|---|---|---|
| CTL | Not sterilized | None | Control soil (autochthonous microorganisms) |
| AS03 | Sterilized | Exiguobacterium indicum As03 | As03 strain (Exiguobacterium indicum) |
| AS04 | Sterilized | Kocuria sediminis As04 | As04 strain (Kocuria sediminis) |
| AS33 | Sterilized | Rhodococcus rhodochrous As33 | As33 strain (Rhodococcus rhodochrous) |
| CS | Sterilized | Consortium As03-AS04-AS33 strains | Consortium (As03-AS04-AS33 strains) |
| CS + MS | Not sterilized | Consortium As03-AS04-AS33 strains | Consortium plus soil microorganisms (consortium As03-AS04-AS33 + autochthonous microorganisms) |
| Variables | Pearson’s Correlation Coefficient | p-Value | Significance |
|---|---|---|---|
| Glyphosate | 0.06563304 | 0.025 | * |
| AMPA | 0.10441577 | 0.005 | * |
| Cr | −0.01319377 | 0.625 | |
| Zn | −0.03910445 | 0.845 | |
| Ni | −0.03034851 | 0.789 | |
| Total_P | −0.06525634 | 0.964 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guardado-Fierros, B.G.; Lorenzo-Santiago, M.A.; Gumiere, T.; Aid, L.; Rodriguez-Campos, J.; Contreras-Ramos, S.M. Glyphosate Biodegradation by Airborne Plant Growth-Promoting Bacteria: Influence on Soil Microbiome Dynamics. Agriculture 2025, 15, 362. https://doi.org/10.3390/agriculture15040362
Guardado-Fierros BG, Lorenzo-Santiago MA, Gumiere T, Aid L, Rodriguez-Campos J, Contreras-Ramos SM. Glyphosate Biodegradation by Airborne Plant Growth-Promoting Bacteria: Influence on Soil Microbiome Dynamics. Agriculture. 2025; 15(4):362. https://doi.org/10.3390/agriculture15040362
Chicago/Turabian StyleGuardado-Fierros, Beatriz Genoveva, Miguel Angel Lorenzo-Santiago, Thiago Gumiere, Lydia Aid, Jacobo Rodriguez-Campos, and Silvia Maribel Contreras-Ramos. 2025. "Glyphosate Biodegradation by Airborne Plant Growth-Promoting Bacteria: Influence on Soil Microbiome Dynamics" Agriculture 15, no. 4: 362. https://doi.org/10.3390/agriculture15040362
APA StyleGuardado-Fierros, B. G., Lorenzo-Santiago, M. A., Gumiere, T., Aid, L., Rodriguez-Campos, J., & Contreras-Ramos, S. M. (2025). Glyphosate Biodegradation by Airborne Plant Growth-Promoting Bacteria: Influence on Soil Microbiome Dynamics. Agriculture, 15(4), 362. https://doi.org/10.3390/agriculture15040362