Phenolic Compounds and Antioxidant Activity of Sprouts from Seeds of Citrus Species



 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Sprouting
2.2. Chemicals
2.3. Samples Extraction
2.4. Total Phenolic Concentration (TPC)
2.5. HPLC Analysis: Chromatographic Conditions
2.6. Antioxidant Activity
2.7. Statistical Analysis
3. Results
3.1. Germination and Sprout Growth
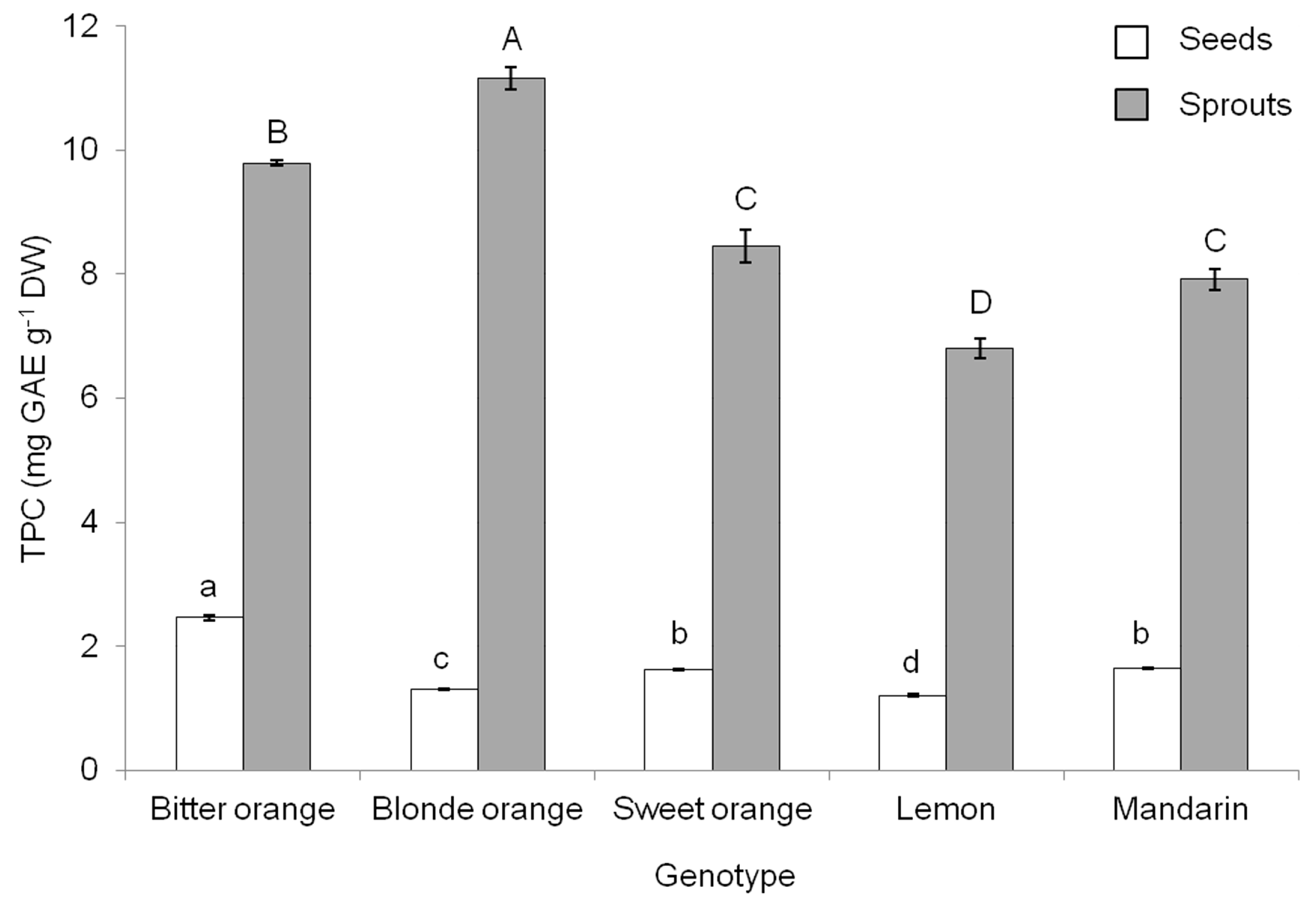
3.2. Total Phenolic Concentration
3.3. Phenolic Acids
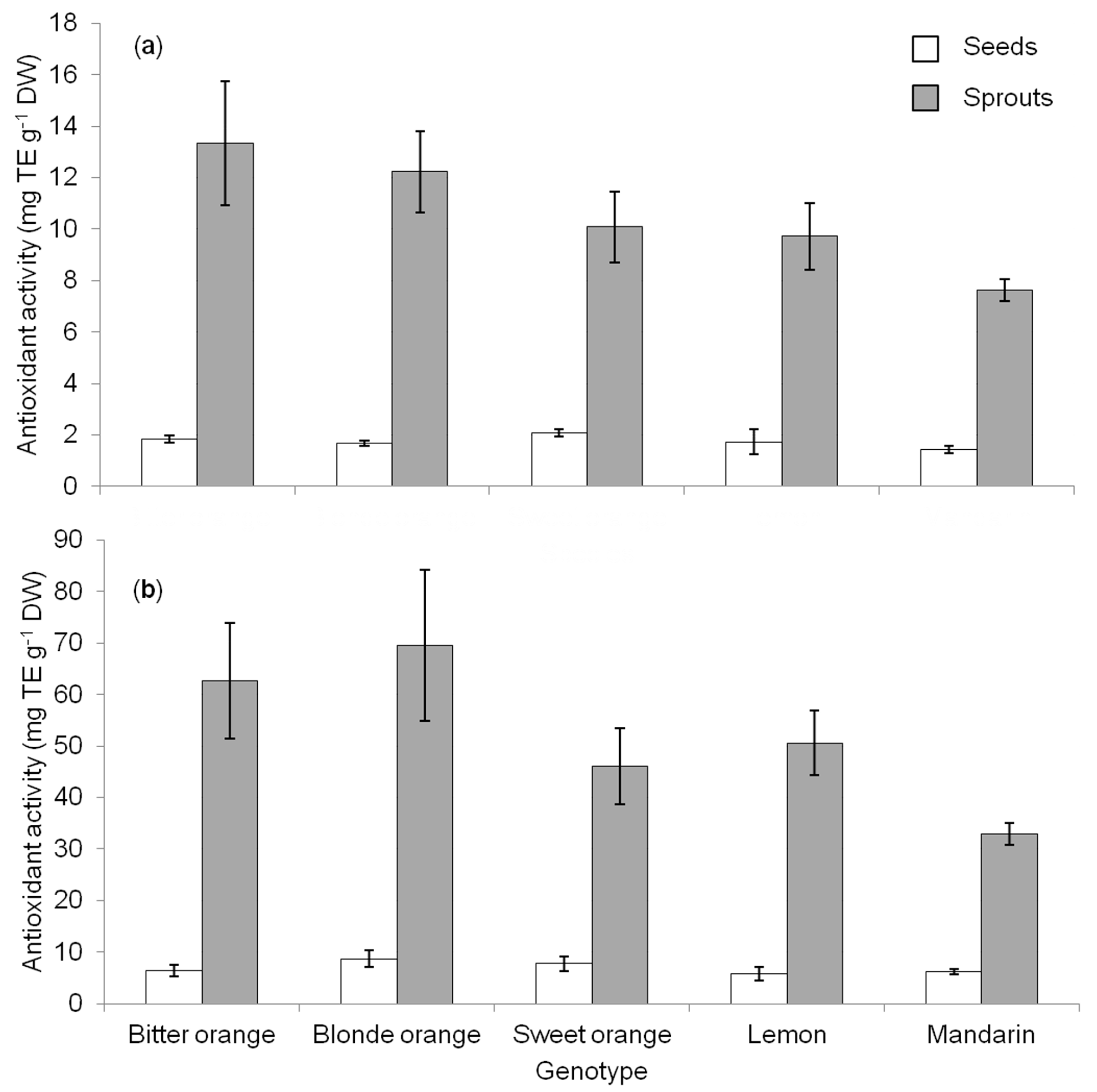
3.4. Antioxidant Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moulehi, I.; Bourgou, S.; Ourghemmi, I.; Tounsi, M.S. Variety and ripening impact on phenolic composition and antioxidant activity of mandarin (Citrus reticulate Blanco) and bitter orange (Citrus aurantium L.) seeds extracts. Ind. Crops Prod. 2012, 39, 74–80. [Google Scholar] [CrossRef]
- Bocco, A.; Cuvelier, M.E.; Richard, H.; Berset, C. Antioxidant activity and phenolic composition of citrus peel and seed extracts. J. Agric. Food Chem. 1998, 46, 2123–2129. [Google Scholar] [CrossRef]
- Malacrida, C.R.; Kimura, M.; Jorge, N. Phytochemicals and antioxidant activity of citrus seed oils. Food Sci. Technol. Res. 2012, 18, 399–404. [Google Scholar] [CrossRef]
- Falcinelli, B.; Marconi, O.; Maranghi, S.; Lutts, S.; Rosati, A.; Famiani, F.; Benincasa, P. Effect of genotype on the sprouting of pomegranate (Punicagranatum L.) seeds as a source of phenolic compounds from juice industry by-products. Plant. Food Hum. Nutr. 2017, 72, 432–438. [Google Scholar] [CrossRef]
- Falcinelli, B.; Maranghi, S.; Paoletti, A.; Marconi, O.; Rosati, A.; Famiani, F.; Benincasa, P. Sprouting olive (Oleaeuropaea, L.) seeds as a source of antioxidants from residual whole stones. Sci. Hortic. 2018, 240, 558–560. [Google Scholar] [CrossRef]
- Benincasa, P.; Falcinelli, B.; Lutts, S.; Stagnari, F.; Galieni, A. Sprouted grains: A comprehensive review. Nutrients 2019, 11, 421. [Google Scholar] [CrossRef]
- Hassanein, A.M.; Azooz, M.M. Propagation of Citrus reticulata via in vitro seed germination and shoot cuttings. Biol. Plant. 2003, 47, 173–177. [Google Scholar] [CrossRef]
- Prajapati, D.G.; Satodiya, B.N.; Desai, A.B.; Nagar, P.K. Influence of storage period and growing media on seed germination and growth of acid lime seedlings (Citrus aurantifolia Swingle) Cv. Kagzi. J. Pharmacogn. Phytochem. 2017, 6, 1641–1645. [Google Scholar]
- Prajapati, D.G.; Satodiya, B.N.; Nagar, P.K. Effect of storage period and growing media on seed germination and physiological attributes of acid lime seedlings (Citrus aurantifolia Swingle) cv. Kagzi. Int. J. Conserv. Sci. 2017, 5, 1945–1948. [Google Scholar]
- Zaher-Ara, T.; Boroomand, N.; Sadat-Hosseini, M. Physiological and morphological response to drought stress in seedlings of ten citrus. Trees 2016, 30, 985–993. [Google Scholar] [CrossRef]
- Xiong, B.; Ye, S.; Qiu, X.; Liao, L.; Sun, G.; Luo, J.; Dai, L.; Rong, Y.; Wang, Z. Transcriptome analyses of two citrus cultivars (Shiranuhi and Huangguogan) in seedling etiolation. Sci. Rep. 2017, 7, 46245. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.A.; Roberts, E.H. The quantification of ageing and survival in orthodox seeds. Seed Sci. Technol. 1981, 9, 373–409. [Google Scholar]
- Benincasa, P.; Galieni, A.; Manetta, A.C.; Pace, R.; Guiducci, M.; Pisante, M.; Stagnari, F. Phenolic compounds in grains, sprouts and wheatgrass of hulled and non-hulled wheat species. J. Sci. Food. Agric. 2015, 95, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Viticult. 1965, 16, 144–158. [Google Scholar]
- Masaldan, S.; Iyer, V.V. Antioxidant and antiproliferative activities of methanolic extract of Aloe vera leaves in human cancer cell lines. J. Pharm. Res. 2011, 4, 2791–2796. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- İnan, Ö.; Özcan, M.M.; Aljuhaimi, F. Effect of location and Citrus species on total phenolic, antioxidant, and radical scavenging activities of some Citrus seed and oils. J. Food Process. Pres. 2018, 42, e13555. [Google Scholar] [CrossRef]
- Hung, V.P.; Hatcher, D.W.; Barker, W. Phenolic acid composition of sprouted wheats by ultra-performanceliquid chromatography (UPLC) and their antioxidant activities. Food Chem. 2011, 126, 1896–1901. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Falcinelli, B.; Sileoni, V.; Marconi, O.; Perretti, G.; Quinet, M.; Lutts, S.; Benincasa, P. Germination under moderate salinity increases phenolic content and antioxidant activity in rapeseed (Brassica napusvaroleifera Del.) sprouts. Molecules 2017, 22, 1377. [Google Scholar] [CrossRef]
- Ti, H.; Zhang, R.; Zhang, M.; Li, Q.; Wei, Z.; Zhang, Y.; Tang, X.; Deng, Y.; Liu, L.; Ma, Y. Dynamic changes in the free and bound phenolic compounds and antioxidant activity of brown rice at different germination stages. Food Chem. 2014, 161, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Heleno, S.A.; Martins, A.; Queiroz, M.J.R.; Ferreira, I.C. Bioactivity of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Genotype | Seed Germination Indexes | Sprout Growth Indexes | |||||
|---|---|---|---|---|---|---|---|
| G (%) | MGT (DAS) | TG50 (DAS) | Fresh Weight (mg) | Dry Matter Concentration (%) | |||
| Bitter orange | 90 (2) | 42 (1) | 43 (1) | 28.3 (1.9) | 20.1 (0.7) | ||
| Blonde orange | 72 (9) | 51 (1) | 62 (5) | 22.4 (2.5) | 20.6 (0.5) | ||
| Sweet orange | 58 (6) | 50 (3) | 68 (4) | 23.0 (2.6) | 21.3 (0.5) | ||
| Lemon | 75 (5) | 35 (1) | 37 (1) | 33.5 (4.5) | 17.5 (1.2) | ||
| Mandarin | 19 (5) | 33 (0) | N.R. | 18.2 (2.2) | 20.9 (1.4) | ||
| Genotype | Phenolic Acids (μg g−1 DW) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Hydroxybenzoic Acids | Hydroxycinnamic Acids | Total | ||||||||
| VA | CaA | FA | p-CA | |||||||
| Seeds | ||||||||||
| Bitter orange | 3.3 (0.2) | 4.8 (0.2) | 3.1 (0.2) c | 15.0 (0.2) b | 26.2 (0.2) b | |||||
| Blonde orange | NQ | ND | 3.4 (0.2) c | 3.1 (0.2) c | 6.5 (0.2) d | |||||
| Sweet orange | NQ | ND | 8.2 (0.5) b | 6.5 (0.3) c | 14.7 (0.8) c | |||||
| Lemon | 13.8 (1.6) | ND | 12.7 (0.9) a | 30.8 (2.4) a | 57.3 (4.7) a | |||||
| Mandarin | 4.4 (0.3) | ND | 7.6 (0.3) b | 6.0 (0.2) c | 18.0 (0.8) c | |||||
| Sprouts | ||||||||||
| Bitter orange | NQ | 6.9 (0.2) B | 22.3 (0.2) B | 11.6 (0.3) B | 40.8 (0.3) C | |||||
| Blonde orange | NQ | 2.8 (0.3) C | 12.7 (0.3) D | 7.1(0.1) D | 22.6 (0.4) E | |||||
| Sweet orange | NQ | 12.9 (0.3) A | 33.9 (0.2) A | 10.2 (0.3) C | 57.1 (0.3) A | |||||
| Lemon | NQ | ND | 15.1 (0.1) C | 32.4 (0.5) A | 47.5 (0.6) B | |||||
| Mandarin | NQ | 13.5 (0.3) A | 9.4 (0.2) E | 4.7 (0.3) E | 27.6 (0.4) D | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falcinelli, B.; Famiani, F.; Paoletti, A.; D’Egidio, S.; Stagnari, F.; Galieni, A.; Benincasa, P. Phenolic Compounds and Antioxidant Activity of Sprouts from Seeds of Citrus Species. Agriculture 2020, 10, 33. https://doi.org/10.3390/agriculture10020033
Falcinelli B, Famiani F, Paoletti A, D’Egidio S, Stagnari F, Galieni A, Benincasa P. Phenolic Compounds and Antioxidant Activity of Sprouts from Seeds of Citrus Species. Agriculture. 2020; 10(2):33. https://doi.org/10.3390/agriculture10020033
Chicago/Turabian StyleFalcinelli, Beatrice, Franco Famiani, Andrea Paoletti, Sara D’Egidio, Fabio Stagnari, Angelica Galieni, and Paolo Benincasa. 2020. "Phenolic Compounds and Antioxidant Activity of Sprouts from Seeds of Citrus Species" Agriculture 10, no. 2: 33. https://doi.org/10.3390/agriculture10020033
APA StyleFalcinelli, B., Famiani, F., Paoletti, A., D’Egidio, S., Stagnari, F., Galieni, A., & Benincasa, P. (2020). Phenolic Compounds and Antioxidant Activity of Sprouts from Seeds of Citrus Species. Agriculture, 10(2), 33. https://doi.org/10.3390/agriculture10020033