Abstract
Specialized pro-resolving mediators (SPMs), including lipoxins, resolvins, protectins, and maresins, actively terminate inflammation and restore tissue homeostasis. This review addresses how specialized pro-resolving mediators (SPMs) and their omega-3/omega-6 PUFA precursors influence inflammatory pathways, disease mechanisms, and therapeutic potential across major inflammatory skin disorders. MEDLINE/PubMed was searched on 4 October 2025. Eligible studies included experimental, animal, mechanistic human, and interventional research examining SPMs or omega-3/omega-6 fatty acids. Non-English articles, reviews, conference abstracts, and dietary questionnaire–only studies were excluded. Two reviewers independently screened and extracted data. Due to heterogeneity, a narrative synthesis was performed. No formal risk-of-bias assessment was undertaken Of 359 records, 57 studies were included (26 psoriasis, 24 atopic dermatitis, 7 acne; scarce hidradenitis suppurativa data). Preclinical data consistently demonstrated that SPMs modulate key inflammatory pathways, support epithelial repair, and help restore immune balance. Human studies revealed altered cutaneous and systemic lipid mediator profiles—characterized by reduced omega-3–derived SPMs and predominance of omega-6-driven inflammatory mediators—suggesting impaired resolution mechanisms across these disorders. Interventional studies showed that omega-3 supplementation may reduce inflammatory markers, improve barrier function, and alleviate clinical symptoms. Early evidence on SPMs analogues and receptor agonists indicates promising therapeutic potential, but clinical data remain sparse. The body of evidence is limited by scarce human data, small sample sizes, heterogeneous interventions and variable methods. Many studies rely on subjective or non-standardized clinical outcomes, and the predominance of experimental models further limits the translational relevance of current findings. In summary, disturbances in PUFA-derived lipid mediator pathways and insufficient activation of pro-resolving mechanisms may contribute to the persistence of cutaneous inflammation. Omega-3 supplementation and SPMs-based novel therapies therefore represent plausible adjunctive approaches; however, their therapeutic relevance requires confirmation in future mechanistic and clinical studies.
1. Introduction
Historically, the resolution of inflammation was considered a passive process that occurred naturally once pro-inflammatory signals diminished. This long-standing concept was overturned by discoveries at the end of the 20th century, which revealed that inflammation resolution is an active and tightly regulated process initiated within the earliest hours of the inflammatory response [1]. Resolution begins with a granulocyte-initiated biochemical switch that redirects arachidonic acid (AA) metabolism from the production of pro-inflammatory prostaglandins and leukotrienes toward the generation of lipoxins. Lipoxins, together with resolvins, protectins, and maresins, constitute a class of specialized lipid mediators known as specialized pro-resolving mediators (SPMs) [2]. These bioactive lipids are enzymatically synthesized from dietary omega-3 and omega-6 PUFAs, including eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA), and AA [2].
SPMs play a pivotal role in concluding inflammatory responses by restricting neutrophil migration [3], promoting macrophage-mediated clearance of apoptotic cells and debris (efferocytosis) [4], and supporting tissue repair and regeneration—all without impairing host immune defense [5]. SPMs limit neutrophil recruitment by reducing chemotaxis, impairing adhesion to endothelial cells, and restricting migration across vascular and epithelial barriers [6]. Moreover, SPMs consistently boost macrophage efferocytic capacity, as mediators such as resolvin D1 (RvD1) and its aspirin-triggered epimer were found to enhance clearance of apoptotic neutrophils [7], while maresin 1 (MaR1) markedly strengthens both efferocytosis and antibacterial phagocytosis [8]. This improved clearance further amplifies local SPMs generation, reinforcing pro-resolving pathways [9]. SPMs redirect macrophages from pro-inflammatory toward pro-resolving states by promoting reparative gene programs and suppressing nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB)–driven cytokine production [10]. Growing evidence shows that SPMs modulate macrophage metabolism by promoting oxidative phosphorylation and fatty-acid oxidation, as well as enhancing mitochondrial function, thereby favoring reparative, pro-resolving functions over pro-inflammatory activity [11]. Beyond their established effects on innate immunity, SPMs also influence adaptive immune responses by limiting memory B-cell proliferation and supporting regulatory T-cell activity [12,13]. They can promote Treg differentiation from naive T cells, enhance Treg functional properties, and indirectly favor Treg-mediated suppression of pro-inflammatory T-cell subsets, thereby contributing to the resolution of inflammation and maintenance of immune homeostasis [12,14]. Inadequate production or dysregulation of these mediators can result in incomplete resolution, contributing to the persistence and progression of various chronic inflammatory and autoimmune conditions [5].
Chronic inflammatory skin disorders, including psoriasis, atopic dermatitis (AD), acne vulgaris, and hidradenitis suppurativa (HS), share overlapping pathogenic mechanisms. These include dysregulation of both innate and adaptive immune responses [15,16,17,18], cytokine-driven inflammation [15,18,19,20], excessive keratinocyte activation [21,22,23], and disruption of skin barrier integrity [18,24]. Despite advances in treatment—such as biologic therapies targeting specific cytokines—current strategies largely focus on suppressing inflammation rather than restoring immune balance and physiological resolution. Consequently, there remains a need for approaches that actively promote the natural termination of inflammation while minimizing long-term immunosuppression.
SPMs offer a novel avenue in this regard, as they restore tissue homeostasis without directly inhibiting immune activation. Their biosynthesis from dietary omega-3 and omega-6 fatty acids (FAs) further suggests that nutritional interventions could serve as practical adjuncts for modulating chronic inflammation. Additionally, SPMs levels and their metabolic pathways hold promise as biomarkers for disease activity and therapeutic response, paving the way for more personalized treatment strategies in dermatology.
The present systematic review aims to provide a comprehensive overview of the evidence regarding SPMs and their omega-3/omega-6 FAs precursors in selected inflammatory skin diseases, including psoriasis, AD, acne vulgaris, and HS. Specifically, this work aims to (i) summarize current knowledge on the roles of SPMs in cutaneous inflammation and resolution, (ii) evaluate interventional studies investigating omega-3/omega-6 FAs supplementation, and (iii) explore the translational potential for therapeutic development and biomarker discovery.
2. Materials and Methods
This systematic review was conducted in accordance with the PRISMA 2020 guidelines. To promote transparency, the full study protocol and search strategy have been archived and are publicly available at https://doi.org/10.17605/OSF.IO/FV9PR. Following peer-review suggestions, the protocol was amended to include a narrative synthesis of the data and an assessment of risk of bias for studies included in the synthesis. We systematically searched MEDLINE (via PubMed) on 4 October 2025, using Medical Subject Headings (MeSH) without date restrictions. The objective of this review was to evaluate the available evidence on SPMs and their dietary precursors—omega-3 and omega-6 FAs—with a focus on their role in the pathogenesis and therapeutic potential of inflammatory skin diseases, including psoriasis, AD, acne vulgaris, and HS.
The PubMed search strategy combined terms for inflammatory skin diseases (“Psoriasis”, “Dermatitis, Atopic”, “Acne Vulgaris”, “Hidradenitis Suppurativa”) with terms related to lipid mediators and fatty acids (“Lipoxins”, “Docosahexaenoic Acids”, “Eicosapentaenoic Acid”, “Fatty Acids, Omega-3”, “Fatty Acids, Omega-6”) using the Boolean operator AND. The complete PubMed search string is provided in the Supplementary Material.
All retrieved records were merged, and duplicates were removed. Eligible studies included original experimental, animal, and human studies addressing the role of SPMs, or omega-3/omega-6 FAs in the context of inflammatory skin diseases. Non-English publications, conference abstracts without full text, narrative reviews, editorials, and commentaries were excluded from the analysis. Interventional studies involving omega-fatty acid supplementation as well as observational studies based on objective biochemical measurements of fatty acid profiles were included, while studies relying solely on self-reported dietary intake were excluded.
To achieve a comprehensive understanding of the SPMs system in inflammatory skin diseases, this review integrates evidence from two main streams: (i) studies measuring or directly reporting SPMs (lipoxins, resolvins, protectins, maresins) in humans, animal models, or experimental systems, and (ii) interventional studies involving the administration of long-chain omega-3 FAs (EPA, DHA) on the course of the disease. The collected evidence was synthesized and compared across these two streams using narrative approach to highlight consistent patterns and mechanistic insights.
Titles and abstracts of all retrieved records were independently screened by two reviewers (A.B.-B. and O.J.-Z.), with disagreements resolved by a third reviewer (H.M.). Full-text articles of potentially relevant studies were then assessed for eligibility. Data extraction was performed independently by two reviewers (A.B.-B. and O.J.-Z.) using a standardized form. Extracted information included study design, population, intervention/exposure characteristics, analytical methods, lipid mediator measurements, and clinical outcomes. We extracted all reported outcomes related to SPMs concentrations, omega-3/omega-6 fatty acid profiles, clinical disease severity, and mechanistic inflammatory markers, irrespective of measurement methods or time points. Additional variables included participant characteristics, disease subtype, intervention dose and duration, model system, and analytical methods; missing or unclear information was extracted as reported without assumptions. Due to heterogeneity in study designs, analytical methods, and outcome measures, the results were summarized using a structured narrative synthesis framework, and therefore no effect measures were predefined, nor were reporting bias or certainty assessments performed.
3. Results
The search resulted in the retrieval of 359 records of which 354 were screened for relevance. Seventy-seven full-text articles were assessed for eligibility, and 20 were excluded. The reasons for exclusion were as follows: (1) assessment of omega-3 exposure based exclusively on food frequency questionnaires (n = 6) [25,26,27,28,29,30], nutritional counseling (n = 2) [31,32], or multi-ingredient supplements not specific to omega-3 fatty acids (n = 1) [33]; (2) outcomes unrelated to inflammatory skin diseases (n = 6) [34,35,36,37,38,39]; (4) ineligible study design, including observational studies focused solely on metabolic biomarkers (n = 2) [40,41]; and case report series (n = 1) [42]; and (5) insufficient or missing data on pro-resolving or anti-inflammatory markers (n = 2) [43,44]. Fifty-seven studies were ultimately included in qualitative synthesis.
The flowchart presenting full search strategy is demonstrated in Figure 1.
Figure 1.
PRISMA 2020 flow diagram illustrating the study identification, screening, eligibility assessment, and inclusion process.
The basic characteristics of the original papers on the researched topic are summarized in Table 1, Table 2, Table 3 and Table 4. Each table includes study design, population characteristics, intervention or exposure details, biochemical endpoints related to omega-3 fatty acids or SPMs, and clinical outcomes relevant to inflammatory skin diseases. In total, 21 experimental, 28 human interventional, and 8 human observational studies were identified, including two that combined in vitro and human investigations. Of the 57 studies, the majority focused on psoriasis (n = 26) and atopic dermatitis (n = 24), with a smaller number addressing acne (n = 7). Given the heterogeneity of included studies, which range from in vitro and animal models to mechanistic human investigations and clinical interventions and the focus on mechanistic insights into SPMs and their precursors, a formal risk of bias assessment was not conducted. Results of individual studies are presented narratively in the disease-specific sections and corresponding tables; quantitative effect estimates were inconsistently reported and could not be standardized. No meta-analysis was conducted, and consequently, heterogeneity analyses, sensitivity analyses, reporting bias assessments, and certainty-of-evidence evaluations were not undertaken.

Table 1.
Effects of Specialized Pro-Resolving Lipid Mediators on Psoriasis Pathophysiology.
Table 1.
Effects of Specialized Pro-Resolving Lipid Mediators on Psoriasis Pathophysiology.
| Author | Year | Population | Design/Intervention | Comparator | Key Observation |
|---|---|---|---|---|---|
| Experimental and Preclinical Studies | |||||
| Chen et al. [45] | 2011 | NHDF and NHEK | Treatment with lipoxin A4 | Untreated control cells | LXA4 suppressed the expression of IL-6 and IL-8 in keratinocytes and fibroblasts. |
| Liu et al. [46] | 2017 | IMQ-induced psoriasiform dermatitis mice model LPS-induced keratinocytes | Treatment with BML111 (LXA4 receptor agonist) | IMQ-induced mice without treatment (vehicle control) | BML111, a lipoxin A4 receptor agonist, treatment reduced IMQ-induced psoriasiform dermatitis. The effect of BML111 and lipoxin A4 is mediated by through HMGB and subsequent downregulation of inflammatory pathways. |
| Xu et al. [47] | 2018 | IMQ-induced psoriasiform dermatitis mice model | Pretreatment with RvD1 | IMQ-induced mice without treatment (vehicle control) | Pretreatment with RvD1 mitigates IMQ-induced psoriasiform dermatitis by targeting ALX/FPR2, and consequent inhibition of the IL-23/IL-17 axis. |
| Sawada et al. [48] | 2018 | IMQ-induced psoriasisiform dermatitis mice model | Treatment with RvE1 | Untreated or IMQ-induced control mice | RvE1 markedly reduced inflammatory infiltration, epidermal hyperplasia, and IL-23 expression in psoriatic skin. RvE1 inhibited IL-23 production and migration of dendritic cells and IL-17–producing γδ T cells through BLT1 antagonism. |
| Park et al. [49] | 2021 | IMQ-induced psoriasiform dermatitis mice model NHEK | Treatment with PD1 | Untreated or IMQ-induced control | PD1 mitigated psoriatic symptoms, epidermal thickening, erythema, and scaling. PD1 reduced pro-inflammatory cytokines and suppressed STAT1 and NF-κB activation in skin lesions. PD1 alleviated systemic inflammation and cytokine expression in keratinocytes. |
| Sorokin et al. [50] | 2023 | Mouse model of psoriasis-like inflammation | Dietary supplementation with DHA or EPA | Normal diet control | DHA supplementation increases skin levels of RvD5, protectin DX, and maresin 2.EPA supplementation reduces skin PGE2 and TXB2 levels. |
| Observational and Mechanistic Human Studies | |||||
| Sorokin et al. [51] | 2018 | Psoriatic patients | Psoriatic patients compared with healthy controls: blood samples, lesional and nonlesional skin punch biopsies. | Healthy controls (matched blood and skin samples) | 14-HDHA, 17-HDHA, precursors of D-series resolvins and protectins) are elevated in psoriatic skin lesions RvD2 is detected in ~50% of psoriasis and healthy individuals (no significant quantitative difference reported) 14,15-EpETE; 17,18-EpETE are mostly undetectable in both psoriasis and control groupsA shift toward ω-6 pro-inflammatory dominance and relative deficiency of ω-3-derived pro-resolving mediators in psoriasis |
| Sorokin et al. [52] | 2018 | Psoriatic patients (blood samples, lesional and nonlesional skin punch biopsies.) NHEK | NHEK cultivated in complete keratinocyte growth media-2 and subsequently treated with vehicle or TNFα or RvD1/RvD5 or TNFα, RvD1 and RvD5. | Vehicle-treated or TNFα-only treated NHEK | RvD5, protectin DX, and lipoxin are found only in lesional PSO skin, while protectin D1 is present in nonlesional PSO skin. RvD1 and RvD5 decrease IL-24 and S100A12 expression in human keratinocytes. |
Abbreviations: ALX/FPR2, lipoxin A4/formyl peptide receptor 2; BLT1, leukotriene B4 receptor 1; DHA, docosahexaenoic acid; EPA, eicosapentaenoic acid; HMGB, high-mobility group box protein; IL, interleukin; IMQ, imiquimod; LPS, lipopolysaccharide; LXA4, lipoxin A4; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; NHDF, normal human dermal fibroblasts; NHEK, normal human epidermal keratinocytes; PD1, protectin D1; PGE2, prostaglandin E2; PSO, psoriasis; RvD1, resolvin D1; RvD2, resolvin D2; RvD5, resolvin D5; RvE1, resolvin E1; SPM, specialized pro-resolving mediator; STAT1, signal transducer and activator of transcription 1; TNFα, tumor necrosis factor alpha; TXB2, thromboxane B2.
Table 2.
Clinical and Preclinical Evidence of ω-3 Polyunsaturated Fatty Acids in Psoriasis.
Table 2.
Clinical and Preclinical Evidence of ω-3 Polyunsaturated Fatty Acids in Psoriasis.
| Author | Year | Population | Design/Intervention | Comparator | Key Observation |
|---|---|---|---|---|---|
| Experimental and Preclinical Studies | |||||
| Karrys et al. [53] | 2018 | Human keratinocytes (HEKn) homozygous for LCE3C_LCE3B deletion | In vitro DHA and curcumin treatment (VDR ligands) | Untreated/TNFα-stimulated cells | DHA and curcumin increased LCE3A/D/E expression, reduced MAPK activation, suggesting anti-inflammatory and skin barrier–repair effects relevant to psoriasis |
| Wannick et al. [54] | 2018 | Mouse model of Aldara-induced psoriasis-like dermatitis | Preclinical study; treatment with Compound A, a potent synthetic FFA4/GPR120 agonist | Vehicle-treated mice | Compound A did not improve psoriasis-like dermatitis, suggesting that sole FFA4 activation is insufficient to mediate the anti-inflammatory effects of ω3-PUFAs |
| Morin et al. [55] | 2021 | In vitro psoriatic and healthy skin substitutes | 10 μM DHA supplementation in culture medium | Unsupplemented control media | DHA reduced abnormal keratinocyte differentiation, decreased PGE2 and 12-HETE, rebalanced PPAR expression, and lowered TNF-α, attenuating psoriatic features. |
| Simard et al. [56] | 2021 | reconstructed 3D skin substitutes: psoriatic and healthy | In vitro 3D skin models derived from healthy and psoriatic donor cells, with keratinocytes cultured in 10 μM α-linolenic acid (ALA, ω-3 PUFA) medium | Skin substitutes cultured in standard medium (ALA) | ALA reduced keratinocyte proliferation (↓ Ki67), improved differentiation (↑ filaggrin, loricrin), incorporated into epidermal phospholipids, metabolized to EPA and ω-3 DPA, increased 15-HEPE and 18-HEPE, decreased ω-6 lipid mediators, and activated ERK1/2 signaling, normalizing psoriatic phenotype |
| Son et al. [57] | 2022 | FFA4 WT and KO BALB/c mice with imiquimod-induced psoriasis-like skin lesions | In vivo animal study; treatment with Compound A (30 mg/kg), a selective FFA4 agonist | Vehicle-treated mice (imiquimod only); FFA4 KO mice | Compound A reduced PASI, epidermal thickness, keratinocyte proliferation, TH17/TH1 cytokines, and CD4+IL-17A+ T cells in FFA4 WT mice; no effect in KO mice, indicating FFA4-mediated anti-psoriatic ω-3 PUFA effects. |
| Morin et al. [58] | 2023 | In vitro healthy and psoriatic skin substitutes with polarized T cells | Culture media supplemented with 10 μM EPA | Unsupplemented psoriatic skin model | EPA increased epidermal EPA, DPA, and DHA, elevated anti-inflammatory lipid mediators (PGE3, 12-HEPE, EPEA), and reduced ω-6–derived metabolites, restoring lipid homeostasis. |
| ω-3 Fatty Acids Intervention Studies on the Course of Psoriasis | |||||
| Gupta et al. [59] | 1989 | 18 patients with stable plaque psoriasis treated with UVB | 15-week RCT; diet enriched in fish oil (3.6 g of EPA 2.4 g of DCHA) | diet enriched in olive oil (76% olecic acid, 9% palmitic acid, 7% linoleic acid, 2%, stearic acid). | Fish oil supplementation led to greater improvement in BSA, redness, scaling, erythema, epidermal thickness, and overall global response compared to olive oil diet. |
| Dewsbury et al. [60] | 1989 | 11 psoriatic patients treated simultaneously with topical EPA and vehicle creams on different lesions | 7-week single-blind within-patient comparison; topical EPA | placebo (vehicle) cream | 8/11 patients showed subjective and objective improvement with EPA-treated plaques vs. placebo |
| Kojima et al. [61] | 1991 | 9 patients with chronic stable psoriasis | EPA supplementation 3.6 g/day for up to 12 months | Baseline/pre-treatment status | Modest but sustained clinical improvement; increased plasma EPA and LTB5, reduced LTB4 |
| Grimminger et al. [62] | 1993 | 21 patients with guttate psoriasis | Double-blind RCT;IV ω-3 emulsion (4.2 g EPA+DHA daily for 10 days) | IV ω-6 lipid emulsion | ω-3 group showed rapid, significant clinical improvement and increased EPA-derived mediators; ω-6 group showed only slight changes |
| Søyland et al. [63] | 1993 | 124 adults with stable plaque psoriasis | 4-month, double-blind RCT; six capsules daily of highly concentrated fish-oil ethyl esters (51% EPA, 32% DHA) | Six capsules daily of corn oil (oleic acid 18:1 n-9, linoleic acid 18:2 ω-6) | Fish-oil raised serum ω-3 PUFA but showed no PASI or symptom improvement vs. corn oil; minor reductions in scaling and infiltration seen in both groups. |
| Henneicke-von Zepelin et al. [64] | 1993 | 52 patients with moderate to severe psoriasis | Multicenter, double-blind RCT; topical ω-3 PUFA cream (1% or 10%) applied for 8 weeks | Placebo cream applied to matched control lesion on the same patient | Both treated and placebo lesions improved vs. baseline, but no significant difference between groups. Topical omega-3 PUFA was well tolerated; one case of mild perilesional eczema. |
| Mayser et al. [65] | 1998 | 83 patients with chronic plaque psoriasis | Double-blind RCT; IV ω-3 lipid emulsion (8.4 g EPA + DHA daily for 14 days) | IV ω-6 lipid emulsion | Greater PASI reduction in ω-3 group (p = 0.048); 37% achieved ≥50% PASI improvement vs. 23% controls; increased plasma EPA and anti-inflammatory eicosanoids |
| Danno et al. [66] | 1998 | 40 patients with moderate plaque psoriasis | 12-week randomized open study; combination therapy etretinate + 1800 mg/day EPA | etretinat | The combination therapy was more effective than monotherapy, especially with regard to improvement >75% |
| Guida et al. [67] | 2014 | 44 obese patients with mild-to-severe plaque psoriasis on stable immunosuppressive therapy | 6-month RCT; an energy-restricted diet enriched in ω-3 PUFAs (2.6 g/day) and low in ω-6 PUFAs | usual diet without nutritional intervention | ω-3 enriched diet significantly improved PASI, itch, and DLQI scores at 3 and 6 months (p < 0.05), and reduced body weight, waist circumference, triglycerides, total cholesterol, and ω-6/ω-3 ratio compared to controls. |
| Kristensen et al. [68] | 2017 | 145 patients with psoriatic arthritis | Randomized, double-blind trial; 3 g/day marine ω-3 PUFA for 24 weeks | 3 g/day olive oil | Improved disease activity measures, including PASI and reduced NSAID/paracetamol use; decreased leukotriene B4 and increased leukotriene B5 formation. |
| Petrovic et al. [69] | 2023 | 64 patients with mild psoriasis | Double blind RCT; Daily oral herring roe oil capsules providing 2.6 g ω-3 PUFA (DHA:EPA 3:1 ratio) | Matching placebo capsules with coconut oil (medium-chain triglycerides) | Significant PASI reduction after 26 weeks vs. placebo. ↓ CD38+ CD4+/CD8+ T cells and CD56^bright NK cells, maintained monocytes, ↓ CCL2, ↑ IFN-γR1 and CXCL10. Cytokine changes correlated with clinical improvement, indicating anti-inflammatory ω-3 PUFA effects. |
| Huang et al. [70] | 2024 | Genome-wide association study datasets | Two-sample Mendelian randomization assessing the causal relationship between fatty acid levels (ω-3, ω-6, and others) and psoriasis risk. | Genetic variants associated with lower or higher fatty acid levels. | Elevated genetically predicted ω-3 fatty acids were associated with lower psoriasis risk; no causal effect observed for other FA types. |
Abbreviations: ALA, α-linolenic acid; BALB/c, Bagg albino laboratory-bred mouse strain c; BSA, body surface area; CD, cluster of differentiation; CCL2, C-C motif chemokine ligand 2; CXCL10, C-X-C motif chemokine ligand 10; DLQI, Dermatology Life Quality Index; DHA, docosahexaenoic acid; DPA, docosapentaenoic acid; DCHA, docosahexaenoic acid (older abbreviation); EPA, eicosapentaenoic acid; EPEA, eicosapentaenoyl ethanolamide; ERK1/2, extracellular signal-regulated kinase 1/2; FA, fatty acid; FFA4, free fatty acid receptor 4 (GPR120); GPR120, G-protein-coupled receptor 120; HEKn, human epidermal keratinocytes (neonatal); HETE, hydroxyeicosatetraenoic acid; HEPE, hydroxyeicosapentaenoic acid; IFN-γR1, interferon gamma receptor 1; IL, interleukin; KO, knockout; LCE, late cornified envelope; LTB, leukotriene B; MAPK, mitogen-activated protein kinase; NK, natural killer; NSAID, non-steroidal anti-inflammatory drug; PASI, Psoriasis Area and Severity Index; PGE2, prostaglandin E2; PGE3, prostaglandin E3; PPAR, peroxisome proliferator-activated receptor; PUFA, polyunsaturated fatty acid; RCT, randomized controlled trial; TH1, T-helper 1 cell; TH17, T-helper 17 cell; TNFα, tumor necrosis factor alpha; UVB, ultraviolet B; VDR, vitamin D receptor; WT, wild-type; ↓, decrease; ↑, increase.
Table 3.
Summary of Clinical, Experimental, and Observational Evidence on Omega-3 Fatty Acids and Specialized Pro-Resolving Mediators in Acne Vulgaris.
Table 3.
Summary of Clinical, Experimental, and Observational Evidence on Omega-3 Fatty Acids and Specialized Pro-Resolving Mediators in Acne Vulgaris.
| Author | Year | Population | Design/Intervention | Comparator | Key Observation |
|---|---|---|---|---|---|
| Khayef et al. [71] | 2012 | 13 males with inflammatory acne | Supplementation of fish oil capsules daily for 12 weeks that contained a total of 930 mg EPA, 720 mg DHA, and 174 mg DPA per 3 capsules. | No control group. | 8 patients reduced acne symptoms, but no significant changes in total lesion count or redness was observed. |
| Jung et al. [72] | 2014 | 45 patients with mild to moderate acne | A 10-week double-blind RCT, parallel study including 45 patients divided in 2 groups: supplementing omega-3 (500 mg/day EPA and 500 mg/day DHA) or suplementing GLA 200 mg/day | A control group on regular diet. | Reduced mean-count inflammatory and non-inflammatory acne lesions in treatment groups compared to placebo group. |
| Aslan et al. [73] | 2016 | 31 female patients with moderate or severe acne | Cross-sectional study measuring serum FA and inflammatory mediators | 21 healthy controls | Significantly decreased EPA levels, increased AA/EPA and GLA/EPA ratios, increased serum LPL, increased sPLA2 in acne vulgaris patients |
| Zainab et al. [74] | 2021 | 60 patients treating acne with oral isotretinoin | Double-blind RCT including 34 patients on isotretinoin (0.5 mg/kg) plus omega-3 (1 mg/kg) | 26 patients on oral isotretinoin (0.5 mg/kg) and placebo | Omega-3 group had significantly fewer cases of cheilitis, lip dryness, and xerosis compared to placebo (p < 0.05). |
| Guertler et al. [75] | 2024 | 60 patients with acne | 16-week interventional study;Mediterranean diet + algal DHA/EPA (600–800 mg DHA, 300–400 mg EPA/day). | No control group; within-subject comparison across visits | Acne patients exhibited an EPA/DHA deficiency; supplementation significantly improved acne severity and reduced DLQI scores |
| Huang et al. [76] | 2024 | 46 acne vulgaris patients taking isotretinoin; three- and six-week-old male Sprague–Dawley rats | RCT: isotretinoin ± ω-3 fatty acids (2.4 g/day for 12 weeks); evaluation by clinical assessment, biochemical markers, gut microbiota. Parallel animal study using acne-induced rats and fecal microbiota transplantation (FMT). | Isotretinoin alone vs. isotretinoin + ω-3 fatty acids; 20 healthy controls. | Combined ω-3 + isotretinoin produced greater clinical improvement, reduced TG, increased HDL, and restored gut microbiota diversity. In rats, ω-3 supplementation reduced inflammation and comedones; FMT from ω-3-treated donors replicated the benefit, confirming a gut–skin axis mechanism. |
| Zhang et al. [77] | 2025 | Data from randomized controlled trials on acne patients and genetic datasets (European ancestry) for MR analysis. | Mendelian randomization evaluating causal links between serum UFA metabolites and acne. | genetic non-exposure variants | MR analysis identified EPA and AA as protective factors and DGLA as a risk factor (Enzymes FADS1/FADS2 implicated in acne regulation). |
Abbreviations: AA—Arachidonic Acid; DGLA—Dihomo-γ-Linolenic Acid; DLQI—Dermatology Life Quality Index; DPA—Docosapentaenoic Acid; DHA—Docosahexaenoic Acid; EPA—Eicosapentaenoic Acid; FA—fatty acids; FADS1/FADS2—fatty acid desaturase 1/2; FMT—Fecal Microbiota Transplantation; GLA—γ-Linolenic Acid; HDL—High-Density Lipoprotein; LPL—Lipoprotein Lipase; MR—Mendelian Randomization; RCT—Randomized Controlled Trial; sPLA2—Secretory Phospholipase A2; TG—Triglycerides; UFA—Unsaturated Fatty Acid; ω-3/ω-6—Omega-3/Omega-6 Fatty Acids.
Table 4.
Summary of Clinical, Experimental, and Observational Evidence on Omega-3 Fatty Acids and Specialized Pro-Resolving Mediators in Atopic Dermatitis.
Table 4.
Summary of Clinical, Experimental, and Observational Evidence on Omega-3 Fatty Acids and Specialized Pro-Resolving Mediators in Atopic Dermatitis.
| Author | Year | Population | Design/Intervention | Comparator | Key Observation |
|---|---|---|---|---|---|
| Experimental and Preclinical Studies | |||||
| Weise et al. [78] | 2011 | BALB/c mouse model of allergen-induced dermatitis | Dietary DHA + AA (24 mg/kg DHA + 48 mg/kg AA daily for 64 days) | Control diet, DHA alone, or AA alone | Oral DHA + AA reduced dermatitis severity and Ki67 expression, increased Foxp3+ T cells and IL-10, and suppressed keratinocyte TSLP; DHA or AA alone ineffective. |
| Kim et al. [79] | 2012 | DNFB-induced mouse AD model | Experimental study. Intraperitoneal RvE1 (100 or 200 ng/mouse/day) on days 7–13 after AD induction. | DNFB-treated (induction-only) and vehicle-treated controls | RvE1 in dose-dependent manner ameliorated DNFB-induced AD via downregulation of IgE and Th1/Th2 cytokine responses, and limiting immune cell infiltration (CD4+ T, CD8+ T, eosinophils, and mast cells) |
| Han et al. [80] | 2015 | DNCB-induced experimental ADmouse model | Experimental study. Oral DHA (100 mg/kg/day in drinking water) for 30 days; DHA-M2 macrophages transfused intravenously at days 12 and 19. | DNCB-induced AD without DHA or DHA-M2 transfusion; hydrocortisone group for comparison | DHA reduced IgE, histamine, ear thickness, epidermal thickness, and inflammatory cell infiltration. DHA increased CD4+Foxp3+ Tregs in LN and skin; suppressed Th1/Th2/Th17 cytokines; increased TGF-β, CTLA-4 DHA-M2 macrophages transferred protective effects, reduced inflammation, increased Tregs at inflammation sites. |
| Yoshida et al. [81] | 2016 | NC/Nga mice with AD-like dermatitis | DHA/EPA administered with FK506 | FK506 alone | Combined DHA/EPA + FK506 reduced dermatitis severity, LTB4 levels, immune cell infiltration, serum IgE, and IL-13/IL-17A secretion; effects reversed by LTB4 injection, indicating LTB4-dependent mechanism. |
| Fujii et al. [82] | 2018 | HR-AD mice with diet-induced AD–like symptoms | Oral administration of EPA ethyl ester (EPA-E) at 600 or 3000 mg/10 mL/kg once daily for 15 days | Vehicle (oleic acid) and HR-AD diet alone | High-dose EPA-E improved AD-like skin symptoms EPA-E lowered TSLP and IL-4 but not IL-5 expression EPA-E administration restored the depleted covalently bound ceramides, which resulted in improved barrier function |
| Lee at al. [83] | 2019 | mouse models of AD and psoriasis | Dietary supplementation with 5% EPA for 4 weeks prior to disease induction; AD induced with Dermatophagoides farinae body extract ointment, psoriasis induced with 5% imiquimod cream | Control diet | EPA altered skin lipid profiles by reducing AA-derived mediators (PGE2, TXB2, LTB4) and increasing EPA metabolites (e.g., RvE1), but failed to reduce scratching or dermatitis severity in AD and psoriasis models. |
| Sato et al. [84] | 2025 | Rat hind paw edema (PAF-induced) and mouse model of AD | Topical squid phospholipids (1% or 5% ointment) extracted from Todarodes pacificus, rich in DHA, EPA, and AA | Vehicle (Vaseline), soybean PC (1%), DHA (1.2%) | ω-3–rich squid phospholipids (DHA/EPA) reduced PAF-induced inflammation and AD-like skin lesions in mice DHA or soybean PC alone ineffective |
| Observational and Mechanistic Human Studies | |||||
| Montes et al. [85] | 2013 | 211 non-atopic mothers and their infants followed-up to 14 months | Prospective cohort assessing maternal and cord plasma FA composition during pregnancy and risk of infant atopic eczema | Non-eczema vs. eczema infants | Higher maternal and cord ω-3 LC-PUFAs (particularly DHA) associated with lower eczema risk |
| Mihály et al. [86] | 2014 | 20 adult AD patients and 20 healthy controls | Cross-sectional study assessing plasma and PBMC fatty acid composition, FADS2 and SCD1 expression | Healthy control group | AD patients showed elevated FADS2 expression in PBMCs and increased FADS2-derived ω-6 PUFAs, with reduced ω-3 PUFA levels; suggesting altered desaturase activity may drive ω-6 dominance and impaired ω-3-mediated resolution pathways. |
| Gardner et al. [87] | 2020 | 1131 mother–child pairs from a USA prenatal cohort (CANDLE study) | Observational prospective cohort. Maternal plasma phospholipid PUFAs measured during 2nd trimester; child AD assessed by ISAAC-based questionnaire at 4–6 years. | Comparison by maternal PUFA profiles (ω-3 PUFA, ω-6 PUFA, ω-6:ω-3 ratio, and combined EPA+DHA quartiles). | Maternal ω-6 PUFA exposure during pregnancy was positively associated with childhood AD, particularly among atopic mothers. ω-3 PUFAs showed no protective association overall, though moderate levels of EPA + DHA might reduce risk. No consistent link was observed for the ω-6:ω-3 ratio. |
| Mao et al. [88] | 2025 | 268,589 UK Biobank participants (after exclusion of prevalent AD cases | Prospective cohort study examining associations between FA profiles and risk of AD over a median 14.4-year follow-up | Comparison across FA quintiles and genetic subgroups | Higher plasma ω -3 and non-DHA ω-3 reduced AD risk, while elevated ω -6/ω -3 ratio increased risk; SFA, MUFA, ω -6, and LA showed no association. ω -3 attenuated genetic risk linked to rs1692120 and rs174448, with ~9% of AD cases potentially preventable through ω -3 supplementation. |
| Interventional Human Studies | |||||
| Bjørneboe et al. [89] | 1987 | 31 adults with moderate to severe AD and personal or family history of atopy | 12-week double-blind RCT. Intervention: capsules containing fish oil (~1.8 g EPA and 1.2 g DHA). | Placebo (olive oil) capsules. Mild topical steroids allowed. | Significant patient-reported improvement in itch, scaling, and total score; no significant physician-rated change or steroid use difference. |
| Berth-Jones et al. [90] | 1993 | 123 patients with AD | 16-week double-blindRCT Active arms: (1) Evening primrose oil, (2) Evening primrose + fish oil,; allowed topical steroids/emollients as required. | Placebo capsules topical steroids/emollients as required. | No significant improvement in disease severity, percentage of skin affected, topical steroid use, or patient symptom |
| Watanabe et al. [91] | 1999 | 64 patients with AD | Topical EPA/DHA ointment (1.2% DHA + 0.6% EPA, applied 2–3×/day for 4 weeks) | Placebo: hydrophilic ointment base applied to contralateral limb (n = 12 subset) | Significant improvement in erythema, papules, scaling, itching, and thickening; >80% improvement vs. 0–40% with placebo |
| Mayser et al. [92] | 2002 | 22 hospitalized patients with moderate-to-severe AD | Double-blind RCTl daily IV infusion of ω-3 fatty acid-based lipid emulsion for 10 days | ω-6 lipid emulsion | IV ω-3 lipid infusion improved AD severity and elevated plasma/membrane EPA and EPA/AA ratio, inducing EPA-derived mediators without affecting lymphocyte function. |
| Dunstan et al.l. [93] | 2003 | 98 atopic pregnant women; follow-up of their infants at high risk of allergic disease to 12 months | Double-blind RCT. Maternal supplementation with fish oil: 4 × 1 g/day capsules (total 3.7 g ω-3 PUFAs: 56% DHA, 27.7% EPA) from mid-pregnancy until delivery. | Olive oil capsules (placebo) | No significant difference in overall incidence of atopic dermatitis between groups. Infants in the fish oil group exhibited lower allergen sensitization rates and significantly reduced severity of AD. |
| Koch et al. [94] | 2008 | 53 adults with atopic eczema; 44 completed (DHA n = 21, Control n = 23) | 8-week double-blind RCT DHA 5.35 g + EPA 0.37 g/day | Isoenergetic control capsules containing MCT. | DHA supplementation significantly improved SCORAD score, reduced affected skin area, decreased plasma TG, increased HDL. Marked rise in ω-3 PUFA levels and reduced ω-6/ω-3 ratio. Ex vivo PBMC showed significant reduction in induced IgE production. |
| Furuhjelm et al. [95] | 2009 | 145 pregnant women with family history of allergic disease; follow-up of their infants to 24 months | Double-blind, RCT. Maternal supplementation ω-3 fatty acids (1.6 g/day EPA + 1.1 g/day DHA) from gestational week 25 until delivery | Placebo: soybean oil (2.5 g/day linoleic acid + 0.28 g/day α-linolenic acid) | ω-3 supplementation in pregnancy reduced IgE-associated eczema in infants during the first 2 years. Overall eczema frequency similar, but fewer IgE-mediated cases in the ω-3 group. Eczema severity (SCORAD) did not differ between groups. Trend toward lower IgE-associated allergic disease overall, especially in children of non-allergic mothers. Higher DHA and lower AA/EPA ratios in mothers and infants were linked to reduced allergic risk. |
| Noakes et al. [96] | 2012 | 123 pregnant women with low fish intake and atopic family history | RCT: 2 portions of salmon/week from 20 wks gestation to delivery (~3.45 g EPA + DHA; 28 µg vit D3) | Habitual low-fish diet | Increased cord plasma EPA + DHA and decreased AA; reduced CBMC IL-2, IL-4, IL-5, IL-10, TNF-α, and PGE2 responses; no difference in IgE, AD incidence, or severity at 6 months |
| Palmer et al. [97] | 2012 | Pregnant women with a personal or family history of allergic disease and their infants followed up to 1 year | Double-blind RCT. From 21 weeks’ gestation until delivery, mothers received fish oil capsules daily (800 mg DHA, 100 mg EPA). | Control group: vegetable oil capsules | Increased cord plasma DHA + EPA; decreased AA; reduced incidence of atopic eczema at 1 year; no difference in overall IgE-associated allergy or food allergy. |
| Wu et al. [98] | 2013 | 60 infants (aged 1–12 months, mean 4.2 months) with acute/subacute facial atopic eczema diagnosed by AAD criteria. | Double-blind RCT. Topical 0.1% 15(R/S)-methyl-lipoxin A4 (LXA4) cream applied twice daily for 10 days. | Placebo cream and 0.1% mometasone furoate (Eloson® Schering-Plough, Shanghai, China) cream. | LXA4 improved erythema and pruritus by day 3, papules/vesicles/scaling by day 5, and lichenification by day 10; effects persisted 1 week post-treatment. Efficacy comparable to mometasone (EASI, SSS), both improved IDQOL. No adverse events; relapse at day 40: LXA4 63%, mometasone 53% |
| Komulainen et al. [99] | 2023 | 439 pregnant women (BMI ≥ 25 kg/m2, <18 weeks gestation, no chronic disease) and 287 infants followed to 24 months | Double-blind RCT. Maternal supplementation: (1) Fish oil (2.4 g ω-3/day; DHA 1.9 g + EPA 0.22 g), (2) Probiotics, (3) Fish oil + probiotics, (4) Double placebo. | Placebo capsules (medium-chain triglycerides) and/or probiotics (microcrystalline cellulose). | No reduction in risk of physician-diagnosed atopic eczema at 12 or 24 months in any intervention group (all p > 0.05). |
| Figueroa-Garduño et al. [100] | 2023 | 193 Mexican mother–child pairs from the POSGRAD cohort; follow-up of infants to 5 years | Double-blind RCT. Prenatal DHA 400 mg/day from 18 to 22 weeks gestation to delivery. Assessment of maternal urinary arsenic at mid-pregnancy. | Soy/corn-oil placebo group without DHA supplementation. | DHA supplementation decreased early-childhood AD and counteracted the effect of prenatal arsenic exposure. |
| Niseteo et al. [101] | 2024 | 52 children (1–8 years) with moderate-to-severe AD | 4-month—triple-blind RCT. Fish oil syrup (600 mg EPA, 400 mg DHA, 10 mg GLA, 5 µg vit D3 daily) + standard AD care | Placebo syrup (MCT oil) | After 4 months, SCORAD significantly decreased in the intervention group vs. no change in placebo. PO-SCORAD, itch, sleep disturbance, and FDLQI improved significantly in the intervention group. No difference in TCS use. |
Abbreviations: AA, arachidonic acid; AA/EPA ratio, arachidonic acid to eicosapentaenoic acid ratio; AAD, American Academy of Dermatology; AD, atopic dermatitis; AD-like, atopic dermatitis–like; CBMC, cord blood mononuclear cell; CTLA-4, cytotoxic T-lymphocyte-associated protein 4; DHA, docosahexaenoic acid; DHA-M2, docosahexaenoic acid–induced M2 macrophages; DNCB, 2,4-dinitrochlorobenzene; DNFB, 2,4-dinitrofluorobenzene; EASI, Eczema Area and Severity Index; EPA, eicosapentaenoic acid; EPA-E, eicosapentaenoic acid ethyl ester; FA, fatty acid; FADS2, fatty acid desaturase 2; FDLQI, Family Dermatology Life Quality Index; FK506, tacrolimus; GLA, γ-linolenic acid; HDL, high-density lipoprotein; HR-AD, hairless atopic dermatitis; IgE, immunoglobulin E; IL, interleukin; ISAAC, International Study of Asthma and Allergies in Childhood; LC-PUFA, long-chain polyunsaturated fatty acid; LN, lymph node; LXA4, lipoxin A4; LTB4, leukotriene B4; MCT, medium-chain triglyceride; MUFA, monounsaturated fatty acid; PAF, platelet-activating factor; PBMC, peripheral blood mononuclear cell; PGE2, prostaglandin E2; PO-SCORAD, Patient Oriented SCORing Atopic Dermatitis; PUFA, polyunsaturated fatty acid; RvE1, resolvin E1; SCORAD, SCORing Atopic Dermatitis; SFA, saturated fatty acid; SSS, Symptom Severity Score; TCS, topical corticosteroid; TG, triglyceride; Th, T helper; TGF-β, transforming growth factor-β; TSLP, thymic stromal lymphopoietin; Treg, regulatory T cell; TXB2, thromboxane B2; ω-3/ω-6, omega-3/omega-6 fatty acid.
3.1. Psoriasis
In total, 26 studies evaluating lipid mediators and omega-3 fatty acids were included in the analysis. Within the first evidence stream, eight studies directly examined SPMs in murine models (n = 6) or human samples (n = 2) (Table 1).
One animal study demonstrated that dietary DHA increased cutaneous levels of several D-series resolvins, protectins, and maresins while simultaneously reducing prostaglandin E2 (PGE2) and thromboxane B2 (TXB2) [50]. The remaining preclinical studies investigated exogenous administration of SPMs or their receptor agonists, consistently showing attenuation of imiquimod-induced psoriasiform dermatitis, decreased epidermal hyperplasia, and suppression of pro-inflammatory cytokines such as IL-6, IL-8, and IL-23, accompanied by inhibition of the IL-23/IL-17 axis, along with downregulation of parallel inflammatory pathways involving signal transducer and activator of transcription 1 (STAT1) and NF-κB [45,46,47,48,49]. Human mechanistic and observational investigations further revealed altered profiles of omega-3– and omega-6–derived lipid mediators in psoriatic lesions compared with nonlesional skin and healthy controls, with lesional–control differences exceeding those observed in blood [51,52].
The second evidence stream predominantly concerned the effects of omega-3 fatty acids on psoriasis (Table 2).
Experimental and preclinical studies in cellular and reconstructed skin models showed that DHA, EPA, α-linolenic acid (ALA), and selective free fatty acid receptor 4 (FFA4) agonists improved keratinocyte differentiation, restored lipid-mediator balance, and reduced TH17/TH1-related cytokines [53,55,56,58]. Two studies examining lipoxin A4 receptor agonists in imiquimod-induced murine models reported divergent clinical outcomes, with one study showing no clinical benefit and the other demonstrating significant improvement in psoriasiform skin inflammation [54,57]. Interventional studies in humans demonstrated heterogeneous but overall beneficial outcomes across topical, oral, and intravenous omega-3 PUFA interventions, including reductions in Psoriasis Area and Severity Index (PASI), improvements in quality-of-life measures, and modulation of inflammatory lipid pathways in selected trials. Topical omega-3 PUFA application showed improvement in treated lesions, though differences versus placebo were generally modest [60,64]. Intravenous omega-3 lipid emulsions were associated with rapid clinical response and increased anti-inflammatory lipid mediators [62,65]. Oral omega-3 supplementation yielded variable outcomes, with most trials reporting improvements in clinical outcomes, including PASI reduction, decreased erythema and scaling, and enhanced quality-of-life scores [59,66,67]. Several studies additionally demonstrated modulation of inflammatory lipid mediators and cytokine profiles, supporting anti-inflammatory effects of omega-3 PUFAs [59,68,69]. However, a large randomized controlled trial of high-dose fish-oil ethyl esters showed no clinical benefit over corn oil despite increased serum omega-3 levels [63]. Finally, Mendelian randomization analysis supported a protective association between genetically predicted omega-3 fatty acid levels and psoriasis risk [70].
The included studies on psoriasis exhibited several sources of bias. Preclinical investigations showed high indirectness due to reliance on imiquimod models or in-vitro keratinocyte systems, which do not fully reflect psoriatic pathogenesis. Human mechanistic studies were constrained by small sample sizes, potential confounding, and low detectability of several SPMs, contributing to measurement bias. Clinical interventions were highly heterogeneous, often lacking detailed randomization, blinding, or preregistered protocols, and showed variable dosing and outcome measures, resulting in high risk of performance, detection, and reporting bias.
3.2. Acne Vulgaris
In total, seven studies evaluated the role of omega-3 and omega-6 fatty acids as well as PUFA-derived lipid mediators in acne vulgaris; however, none of the included studies directly quantified SPM levels (Table 3).
Evidence from preclinical studies demonstrated that omega-3 supplementation reduced inflammation and comedone formation in acne-induced rats, while fecal microbiota transplantation from omega-3-treated donors confirmed a gut–skin axis mechanism [76]. Observational and genetic evidence indicates that acne patients often present with reduced EPA levels and elevated AA/EPA ratios, while Mendelian randomization analysis supports a protective role of EPA and AA, and a risk-enhancing effect of DGLA, implicating fatty acid desaturase 1 (FADS1) and fatty acid desaturase 2 (FADS2) enzymes in acne pathophysiology [73,77]. Within the interventional evidence stream, five studies evaluating omega-3 fatty acid supplementation showed reductions in acne lesion counts, improvements in patient-reported outcomes, or attenuation of isotretinoin-induced adverse effects [71,72,74,75,76]. Mediterranean diet supplemented with algal DHA/EPA improved acne severity and quality-of-life scores in a 16-week study [75]. In general, omega-3 fatty acids appear to exert protective effects in acne, reducing lesion severity and alleviating treatment-related adverse effects during isotretinoin therapy, although the magnitude of effects varies across studies. Additionally, omega-3 supplementation may beneficially modulate the gut microbiome, highlighting potential systemic mechanisms underlying their therapeutic action.
The included acne studies were limited by small sample sizes, indirect preclinical models, and confounding in human studies. Importantly, none of the studies directly quantified SPMs. Interventional trials were heterogeneous in design, dosing, and outcome measures, resulting in moderate-to-high risk of bias across performance, detection, and reporting domains.
3.3. Atopic Dermatitis
As summarized in Table 4, a total of 24 studies addressing AD met the inclusion criteria of this systematic review.
Among these, only two studies directly investigated specialized pro-resolving mediators [79,98]. In preclinical model of AD, intraperitoneal administration of RvE1 attenuated disease severity by suppressing serum IgE, Th1/Th2 cytokine responses, and inflammatory cell infiltration in dose-dependent manner [79]. In interventional human studies, topical application of lipoxin A4 in infants with atopic dermatitis resulted in rapid and clinically significant improvements in erythema, pruritus, and inflammatory skin lesions, with efficacy comparable to that of a topical corticosteroid [98].
Regarding omega—fatty acids, a total of 22 eligible studies were identified, including 6 preclinical investigations in animal models, 4 observational studies in humans, and 12 interventional trials. The latter comprised 1 topical intervention [91], 1 intravenous administration [92], 4 oral omega-3 supplementation studies in patients with atopic dermatitis [89,90,94,101], and 6 prenatal supplementation trials assessing the development of allergic diseases, including atopic dermatitis, in infants [93,95,96,97,99,100].
Across preclinical models of atopic dermatitis, dietary or topical administration of omega-3 fatty acids, including DHA and EPA, generally reduced skin inflammation and disease severity, serum IgE levels, inflammatory cell infiltration, and Th2/Th17-related cytokines, while enhancing regulatory immune responses [78,80,81,82,84]. Several studies also demonstrated suppression of keratinocyte-derived thymic stromal lymphopoietin (TSLP) and partial restoration of epidermal barrier function [78,82], whereas selected models showed changes in lipid mediator profiles without clear clinical improvement [83].
Observational and mechanistic human studies indicate that higher maternal or circulating omega-3 long-chain PUFAs, particularly DHA, are generally associated with a lower risk of atopic dermatitis in infants [85,88], whereas elevated omega-6 levels or a higher omega-6/omega-3 ratio increase disease risk [86,87]. In one study, moderate EPA and DHA were linked to lower risk, while omega-3 PUFAs overall and the omega-6/omega-3 ratio showed no consistent association [87]. In patients with atopic dermatitis, FADS2 activity was elevated, resulting in increased omega-6 PUFA production and decreased omega-3 levels [86].
In interventional studies, topical application of EPA/DHA ointment significantly improved clinical outcomes in patients with AD [91], while intravenous infusion of an omega-3–based lipid emulsion reduced disease severity and increased plasma and membrane EPA levels and the EPA/AA ratio [92]. Among studies investigating oral omega-3 supplementation in patients with AD, results were mixed. Some trials reported significant improvements in patient-reported outcomes [89,101], SCORAD scores [94,101], affected skin area, and immunologic markers [94], while other studies found no significant effect on disease severity, topical steroid use, or clinical symptoms [89,90]. In maternal supplementation studies, omega-3 fatty acid intake during pregnancy generally increased DHA and EPA levels in maternal and cord plasma [96,97]. Effects on infant atopic outcomes varied: some trials reported reduced severity of atopic dermatitis [93], lower overall incidence of AD [97,100], reductions limited to IgE-associated eczema [95], or decreased allergen sensitization [93,95], while others found no significant differences in overall incidence or severity of AD during the first 1–2 years of life [93,96,99].
The included AD studies exhibited multiple sources of bias. Preclinical studies were generally indirect, relying on murine or rat models that may not fully reflect human disease. Sample sizes were small, and outcomes were often surrogate or mechanistic, contributing to indirectness and potential measurement bias. Observational human studies were subject to confounding, and variability in fatty acid assessment methods. Interventional trials varied widely in design, dosing, route of administration, and outcome measures. Only two studies directly assessed SPMs, limiting direct mechanistic evidence in humans. Prenatal supplementation trials were generally well-conducted but heterogeneous in timing, dose, and follow-up duration, leading to variable effects on AD incidence or severity.
3.4. Factors Contributing to Outcome Differences
Heterogeneity across studies was observed for all three disease entities—psoriasis, acne vulgaris, and atopic dermatitis—arising from multiple sources. Preclinical studies varied in model type (murine, rat, or in vitro keratinocyte/skin substitutes), induction method, and outcome assessment, limiting direct translational comparability. Clinical studies differed in population characteristics, intervention type (oral, intravenous, topical), dose, duration, and timing of supplementation. Outcome measures were diverse, including disease severity scores, lesion counts, cytokine profiles, lipid mediator levels, and quality-of-life indices. Methodological differences, such as randomization, blinding, sample size, and follow-up length, further contributed to variability. Collectively, these factors likely explain the observed heterogeneity in intervention effects and highlight the need for standardized protocols in future trials.
4. Discussion
4.1. Biosynthesis of Lipid Mediators
Lipid mediators are a collective term referring to signaling molecules derived from the metabolism of PUFAs, which play a key role in regulating the inflammatory response. This broad category includes both pro-inflammatory and pro-resolving molecules, with the overall control of inflammation depending on the balance between two main enzymatic pathways: the cyclooxygenase (COX) and lipoxygenase (LOX) pathways [102]. In the initial step of the inflammatory response, phospholipase A2 (PLA2) releases PUFAs from membrane phospholipids. These FAs are subsequently metabolized by COX, LOX, and cytochrome P450 enzymes (CYP450) into various classes of lipid mediators. COX metabolizes the omega-6 AA into pro-inflammatory eicosanoids such as prostaglandins and thromboxanes. However, one of the intermediate metabolites can serve as a substrate for 5-lipoxygenase, leading to the production of lipoxins [103].
The primary precursors of most SPMs are the omega-3 FAs EPA and DHA. Through enzymatic processing by the 5-LOX and 12-LOX pathways, EPA is converted into the E-series resolvins (RvE1–RvE4). In contrast, DHA gives rise to the D-series resolvins (RvD1-RvD6), protectins, and maresins—key lipid mediators that collectively promote the resolution phase of inflammation [104]. Figure 2 illustrates the interconnected biosynthetic pathways that generate both pro-inflammatory and pro-resolving mediators PUFAs.
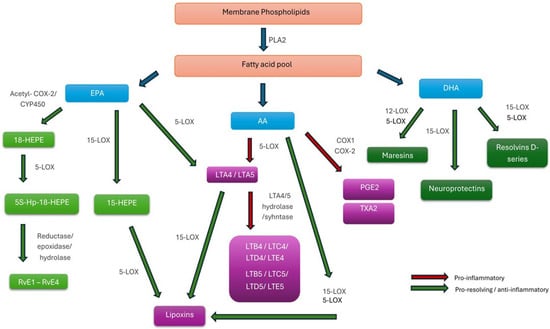
Figure 2.
Biosynthetic Pathways of Pro-Inflammatory and Specialized Pro-Resolving Lipid Mediators Derived from Polyunsaturated Fatty Acids. Abbreviations: AA, arachidonic acid; EPA, eicosapentaenoic acid; DHA, docosahexaenoic acid; PLA2, phospholipase A2; COX-1/COX-2, cyclooxygenase-1/2; acetyl-COX-2, aspirin-acetylated cyclooxygenase-2; CYP450, cytochrome P450 monooxygenase; LOX, lipoxygenase; 5-LOX/12-LOX/15-LOX, 5-, 12-, or 15-lipoxygenase; PG, prostaglandin; PGE2, prostaglandin E2; TXA2, thromboxane A2; LT, leukotriene; LTA4–LTE4, leukotrienes A4–E4; HEPE, hydroxy-eicosapentaenoic acid; HpEPE, hydroperoxy-eicosapentaenoic acid; RvE1–4, E-series resolvins; RvD1–6, D-series resolvins.
Dietary omega-3 FAs, primarily EPA and DHA, can be incorporated into membrane phospholipids, thereby increasing the pool of substrates available for the synthesis of SPMs. Consequently, a diet enriched in these FAs enhances the body’s capacity to generate such mediators. The beneficial and protective roles of omega-3 FAs in mitigating chronic inflammatory diseases across various organ systems are well established [105]. Nonetheless, despite decades of research, the therapeutic efficacy of omega-3 supplementation remains a matter of debate, as findings from controlled clinical trials have been inconsistent [106,107,108,109]. The identification of SPMs and their specific receptors has created new opportunities for the development of targeted pharmacological interventions that modulate these pathways. This is of particular interest because pro-resolving mechanisms are anticipated to achieve inflammation control with fewer adverse effects compared to conventional anti-inflammatory agents [104].
4.2. Overview of SPMs Classes
SPMs comprise four major families—lipoxins, resolvins, protectins, and maresins [104]. The individual classes of SPMs differ in their functional profiles and timing of action. Each class reaches peak concentrations and displays maximal biological activity at distinct stages of the inflammatory process [104]. Moreover, several SPMs exhibit tissue-specific patterns of distribution, achieving their highest levels in organs characterized by high cellular metabolic activity [110].
The biological effects of SPMs are mediated through binding to specific receptors, enabling precise modulation of cellular responses [111]. Their actions are complementary and occur in a defined chronological sequence, ensuring a smooth transition from the pro-inflammatory phase to the gradual reduction in inflammatory infiltrates and pro-inflammatory mediators, activation of regenerative processes, tissue repair, and ultimately the restoration of homeostasis [104]. These processes are schematically illustrated in Figure 3, which summarizes the key pro-resolving, immunomodulatory, and other actions of SPMs leading to restoration of homeostasis.
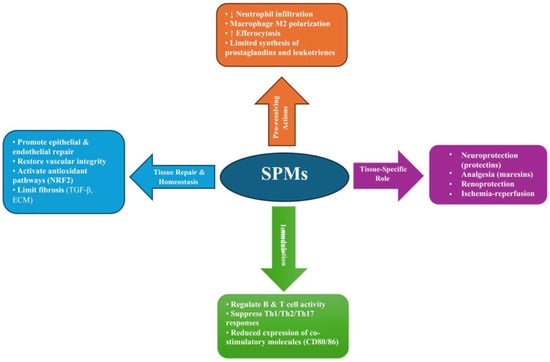
Figure 3.
Overview of the pro-resolving and immunomodulatory actions of SPMs, highlighting their roles in resolving inflammation, regulating immune responses, and promoting tissue repair and homeostasis. Abbreviations: SPMs, Specialized Pro-Resolving Mediators; Th1/Th2/Th17, T helper cell subsets type 1, 2, and 17; CD80/CD86, Co-stimulatory molecules expressed on antigen-presenting cells; NRF2, Nuclear Factor Erythroid 2–Related Factor 2; TGF-β, Transforming Growth Factor beta; ECM, Extracellular Matrix; M2, Alternatively activated (anti-inflammatory) macrophage phenotype; ↓, decrease; ↑, increase.
4.2.1. Lipoxins
Lipoxins were the first identified members of the SPMs family [103]. They are produced in multiple tissues via transcellular pathways involving cooperative interactions between migrating polymorphonuclears and epithelial or endothelial cells at the site of inflammation [112]. Lipoxins function predominantly during the early phase of resolution, exerting multiple actions that together promote the termination of inflammation and support tissue homeostasis. Acting primarily through the Lipoxin A4 Receptor/Formyl Peptide Receptor 2 (ALX/FPR2), they limit the production of pro-inflammatory cytokines such as tumor necrosis factor α (TNF-α) and IL-1β, and promote epithelial repair together with the restoration of vascular barrier integrity [113]. Lipoxins modulate both innate and adaptive immune responses to facilitate the resolution of inflammation. They reprogram macrophages toward a proresolving, M2-like phenotype, enhancing efferocytosis and activating transcriptional programs that support tissue repair, while simultaneously reducing neutrophil infiltration [114]. Beyond their effects on innate immunity, lipoxins influence adaptive immune cells by regulating B and T cell responses during the post-resolution phase and limiting the proliferation of memory B cells [12,13]. In addition to their immunomodulatory functions, lipoxins limit fibrosis by modulating transforming growth factor-β (TGF-β) signaling, inhibiting fibroblast proliferation, and reducing extracellular matrix deposition, with evidence that they can reverse established fibrotic lesions in experimental models [115]. In addition, they activate cytoprotective antioxidant pathways, including nuclear factor erythroid 2–related factor 2 (NRF2) signaling, thereby protecting tissues from oxidative stress [116]. Taken together, these actions highlight the central role of lipoxins in orchestrating the transition from inflammation to resolution while preserving tissue integrity and promoting repair.
The metabolism of eicosanoids can be modulated by certain pharmacological agents in ways that favor the formation of lipoxins. For instance, aspirin-induced acetylation or statin-mediated nitrosylation of COX-2 can shift the enzyme’s catalytic function from its typical endoperoxidase activity toward a lipoxygenase-like pathway [117,118]. This functional reprogramming promotes the biosynthesis of epimeric molecules, commonly referred to as aspirin-triggered lipoxins (ATLs). The epimers retain the pro-resolving and anti-inflammatory properties characteristic of native lipoxins but exhibits enhanced metabolic stability and resistance to enzymatic degradation [119]. Other strategies for modulating the lipoxin pathway include the use of stable synthetic analogs, receptor-specific agonists targeting ALX/FPR2, and cell-penetrating peptides that influence intracellular G protein–coupled receptor (GPCR) signaling, all of which enhance lipoxin-mediated resolution and provide potential avenues for therapeutic intervention in chronic inflammatory diseases [120,121].
4.2.2. Resolvins
Resolvins constitute a distinct family of SPMs biosynthesized from omega-3 FAs and are categorized into two main series: the E-series (RvE1–RvE4), derived from EPA, and the D-series (RvD1–RvD6), originating from DHA [120,122,123]. Biologically, resolvins differ from lipoxins in several key aspects. Unlike lipoxins, which act primarily by limiting neutrophil infiltration, resolvins exert broader actions across immune, vascular, and neural systems [120]. They not only dampen inflammation but also actively promote tissue repair, angiogenesis, and neuroprotection, functioning at picomolar to nanomolar concentrations. Aspirin-triggered (AT) epimers, such as AT-RvD1 and AT-RvD3, are generated via acetylation of COX-2 by aspirin, which redirects biosynthetic pathways toward pro-resolving mediators [124,125]. Although resolvins generally signal through distinct receptors, not all have fully characterized binding partners. Some can engage receptors shared with lipoxins, interacting with alternative binding sites and functioning as biased agonists, thereby eliciting divergent intracellular signaling pathways [126]. Collectively, these account for their therapeutic potential in chronic inflammatory diseases, infections, asthma, atherosclerosis, and neurodegenerative disorders.
4.2.3. Protectins
Protectin D1 (PD1), also known as neuroprotectin D1 (NPD1) in neural tissues, is a DHA–derived SPM whose actions depend on tissue context and cell type [127]. In the nervous system, NPD1 promotes neuronal survival by counteracting oxidative stress and enhancing apoptosis [127]. Outside the nervous system, PD1 functions as an immunoresolvent, limiting cytokine release, reducing neutrophil infiltration, and promoting macrophage-mediated clearance [127]. It also exhibits antiviral activity and contributes to tissue repair [128]. The dual capacity of PD1 to suppress inflammation while enhancing cell survival highlights its relevance as a potential therapeutic mediator in neurodegenerative, retinal, and ischemic pathologies.
4.2.4. Maresins
Maresins are SPMs derived from DHA, possessing both anti-inflammatory and pro-resolving properties. The group comprises three primary forms: maresin 1 (MaR1), maresin 2 (MaR2), and maresin conjugates in tissue regeneration (MCTR) [129]. These mediators are produced by 12-LOX in macrophages and dendritic cells, where they promote efferocytosis and facilitate the resolution of inflammation. Maresins also suppress the expression of pro-inflammatory cytokines and limit polymorphonuclear leukocyte infiltration [130]. Notably, unlike other classes of pro-resolving mediators, maresins exhibit analgesic effects [131], highlighting their unique role in both inflammation resolution and pain modulation.
4.3. Lipids as a Structural and Bioactive Regulators of Cutaneous Homeostasis
The skin is a lipid-rich organ, and its barrier function critically depends on the composition of PUFAs [132]. Both omega-3 and omega-6 PUFAs are integral components of epidermal phospholipids and ceramides. Omega-6 PUFAs, particularly linoleic acid, facilitate ceramide synthesis, thereby enhancing barrier function and reducing transepidermal water loss (TEWL) [133], whereas omega-3 PUFAs exhibit anti-inflammatory and antioxidant properties, modulating cutaneous immune responses and protecting against environmental stressors such as UV radiation and pollution [134,135]. Beyond structural roles, these FAs serve as precursors for a diverse array of bioactive lipid mediators, including SPMs [2].
Dysregulation of PUFAs metabolism and chronic inflammation are central features in the pathogenesis of persistent dermatological conditions, including AD, psoriasis, and acne [56,136,137]. Altered ratios of omega-6 to omega-3 PUFAs can exacerbate pro-inflammatory signaling, leading to sustained epidermal barrier dysfunction, increased TEWL, and heightened susceptibility to environmental insults [138,139]. Moreover, chronic inflammatory states are often accompanied by disturbances in lipid homeostasis, including impaired ceramide synthesis and altered phospholipid composition, which further compromise barrier integrity and skin hydration [140]. These observations have stimulated increasing interest in dietary interventions as modulators of disease course, with omega-3 PUFAs supplementation demonstrating benefits in reducing inflammatory markers, restoring lipid balance, and improving clinical outcomes in chronic inflammatory skin disorders [58,67]. Enhanced availability of omega-3 PUFAs facilitates the enzymatic conversion of EPA and DHA into their respective SPMs families, thereby augmenting the intrinsic capacity of the host to generate pro-resolving lipid mediators [141,142]. Accumulating evidence shows that omega-3 PUFAs can also directly modulate immune regulation by promoting expansion and functional activity of Tregs, shifting immune balance toward resolution rather than inflammation. For instance, in a murine model of atopic dermatitis, administration of DHA enhanced the generation of CD4+Foxp3+ Tregs and increased anti-inflammatory IL-10/TGF-β signaling, reducing skin inflammation [80]. Another example comes from a 3D psoriatic skin model, where EPA treatment decreased IL-17A–secreting T cells and increased FOXP3+ Treg populations, resulting in normalized keratinocyte proliferation and reduced inflammatory mediator levels [58]. Consequently, an increased dietary or systemic supply of omega-3 FAs serves not only as the substrate pool but also promotes the biochemical milieu favorable for the resolution of inflammation. Targeting PUFAs composition through nutrition represents a promising adjunct strategy for managing chronic skin diseases by addressing both inflammation and lipid metabolic imbalance.
4.4. Psoriasis
4.4.1. Lipid Dysregulation in Psoriasis
Psoriasis is widely recognized as a disease predominantly driven by Th1/Th17 immune responses, with elevated levels of cytokines such as IL-17A, IL-17F, IL-22, IL-23, TNF-α, and IFN-γ playing central roles in its pathogenesis [143]. While traditionally considered an immune-mediated disorder developing in genetically predisposed host, increasing evidence highlights a crucial contribution of lipid metabolism to its pathogenesis. Psoriatic skin exhibits altered lipid composition, including disturbed levels of ceramides, free FAs, and eicosanoids, which can impair barrier integrity and amplify inflammatory cascades through the AA and prostaglandin pathways [51,140]. Genes involved in lipid metabolism, such as PLA2G4E, ELOVL, and FABP family members, have been shown to be dysregulated in psoriatic skin, indicating that altered lipid processing may play a role in disease pathogenesis [140,144,145]. These genetic variations may alter the synthesis or turnover of structural and signaling lipids, thereby facilitating immune activation and perpetuating chronic inflammation. Recent research has provided evidence that higher genetically predicted omega-3 FAs levels are associated with a reduced risk of psoriasis, thereby strengthening the evidence that omega-3 FAs may play a direct, biologically meaningful role in modulating psoriasis susceptibility [70]. Thus, the interplay between genetic predisposition and lipid dysregulation represents a key but still incompletely understood component of psoriasis pathogenesis.
These genetic and metabolic disturbances are reflected in a distinct lipidomic profile observed in individuals with psoriasis, characterized by an imbalance between unsaturated FAs classes [146]. The observed shift toward a higher proportion of monounsaturated species suggests altered desaturase activity and disrupted metabolic regulation, while the concurrent reduction in polyunsaturated fractions and a distorted omega-6 to omega-3 equilibrium may contribute to the pro-inflammatory milieu typical of psoriasis. Current evidence indicates that patients with psoriasis display decreased levels of omega-3 PUFAs and elevated omega-6 FAs in both serum and erythrocyte membranes, correlating with higher inflammatory markers such as C-reactive protein (CRP) and IL-6, as well as greater disease severity [146,147]. Studies also suggest that an increased omega-6/omega-3 ratio and elevated saturated FAs content may promote systemic inflammation, metabolic disturbances, and comorbidities including obesity, hepatic dysfunction, and cardiovascular risk [147,148].
4.4.2. Evidence for Omega-3 Fatty Acids Supplementation
Investigations into the therapeutic potential of omega-3 FAs supplementation, including topical applications, in psoriasis began in the late 1980s [59,60]. Early studies demonstrated encouraging outcomes in small patient cohorts; however, these findings were subsequently scrutinized, partly due to the lack of double-blind study designs. In the following decades, further evidence emerged from randomized, double-blind trials; however, the findings remained heterogeneous, primarily due to variations in dosage, study duration, and selected clinical endpoints. Two small studies evaluating the efficacy of a topical formulation containing omega-3 FAs were conducted in comparison with a placebo cream, applied simultaneously to matched lesions within the same patients [60,64]. The first report, involving a cohort of 11 patients with psoriasis, demonstrated objective improvement and suggested the superiority of the omega-3 cream over the placebo [60]. However, a subsequent larger, multicenter study found that general improvement occurred with both formulations, but neither formulation was significantly more effective than the other [64]. The vast majority of available data come from clinical trials investigating the impact of oral supplementation with omega-3 FAs on the course of psoriasis. Most studies demonstrated a generally positive effect on disease activity and associated symptoms, including pruritus; however, differences were not always statistically significant when assessed using objective measures such as the PASI score [67,68]. Interpretation of these results is challenging due to the heterogeneity of study designs, with dosages ranging from 2.6 to 3.6 g/day of EPA ± DHA and treatment durations from 12 weeks to 12 months [61,67]. According to the findings reported by Petrovic et al. [69], the beneficial effect of omega-3 PUFAs supplementation in plaque psoriasis appears to be mediated by anti-inflammatory mechanisms, as evidenced by alterations in immune cell populations and cytokine profiles that correlated with clinical improvement. Notably, two studies assessed the effects of intravenous monotherapy with omega-3–enriched lipid emulsions in comparison to omega-6–based formulations [62,65]. Both studies reported a significant reduction in PASI scores alongside an increase in omega-3–derived eicosanoids, suggesting a distinct therapeutic effect attributable to high daily doses of EPA and DHA. Furthermore, omega-3–enriched parenteral nutrition is recognized as a key component of postoperative care and the management of critically ill patients, as emphasized in current guidelines on intravenous lipid emulsions [149]. Although the meta-analysis assessing fish oil supplementation in psoriasis indicated no statistically significant improvement in PASI scores compared to controls (MD −0.28; 95% CI −1.74 to 1.19; I2 = 57%), individual studies within the analysis reported trends toward improvement in body surface area, as well as reductions in erythema, scaling, and induration, particularly when fish oil was combined with adjunctive therapies [150]. Morin et al. demonstrated in in vitro models of psoriatic and healthy skin that supplementation with DHA or EPA—exerts notable anti-inflammatory and homeostatic effects. DHA reduced abnormal keratinocyte differentiation, lowered pro-inflammatory mediators (PGE2, 12-HETE, TNF-α), and rebalanced PPAR expression, thereby attenuating psoriatic characteristics [55]. EPA supplementation enriched epidermal omega-3 lipid content, increased anti-inflammatory metabolites (PGE3, 12-HEPE, EPEA), and decreased omega-6–derived products, restoring lipid and inflammatory equilibrium [58]. Moreover, DHA supplementation may also regulate epidermal keratinization processes [53]. In approximately 75% of psoriasis cases, a deletion of two late cornified envelope genes (LCE3B–C) is observed [151]. DHA and other omega-3 FAs have been shown to act as potent modulators of epidermal inflammation and differentiation in psoriasis. An open question remains regarding the precise pathways through which omega-3 FAs exert their beneficial effects in alleviating psoriasis. It has been postulated that these mechanisms involve competition as substrates for COX, increased production of SPMs, and modulation of the FFA4 receptor [54,57]. Two studies investigating a synthetic FFA4 agonist yielded conflicting results. A study led by Wannick et al. [54] reported no improvement in a murine model of psoriasis. In contrast, Son et al. observed both clinical improvements and reductions in pro-inflammatory mediators, confirming that the effects were mediated via the FFA4 receptor [57]. Notably, the two studies differed in the route of administration (oral versus parenteral, respectively), which may account for the observed discrepancies.
4.4.3. Evidence for SPMs in Psoriasis
Although the clinical effects of omega-3 supplementation may result from multiple mechanisms, experimental evidence suggests that enhanced SPMs biosynthesis and modulation of the IL-23/IL-17 axis play a key role. The first direct evidence supporting this theory dates back to 2011, when a research team demonstrated that lipoxin A4, in vitro, inhibits the expression of IL-6 and TNF-α in both keratinocytes and fibroblasts [45]. In line with these findings, treatment with a synthetic LXA4 agonist markedly reduced psoriatic inflammation in a murine model [46]. Similarly, Xu et al. demonstrated that pretreatment with RvD1 attenuated imiquimod (IMQ)-induced psoriasiform dermatitis by activating ALX/FPR2, leading to the suppression of the IL-23/IL-17 inflammatory axis [47]. A key downstream pathway implicated in post-receptor signaling involves high-mobility group box protein 1 (HMGB1), a pro-inflammatory mediator that amplifies the IL-23/IL-17A pathway, promotes keratinocyte activation, and facilitates the recruitment of immune cells to psoriatic lesions [152]. Elevated HMGB1 levels have been correlated with disease severity and the persistence of inflammatory responses in psoriasis [152].
Several other SPMs, including RvE1 and PD1, have also been identified as potent modulators of psoriatic inflammation [48,49]. Their administration reduces epidermal scaling, plaque thickness, and erythema, reflecting restoration of epidermal homeostasis. In addition, these mediators downregulate key cytokines involved in psoriasis pathogenesis. In preclinical models, SPMs have been shown to downregulate IL-17 and TNF-α expression, suppress the Th17 axis, reduce immune cell infiltration, and mitigate psoriasiform skin inflammation, highlighting their potential as pro-resolving therapeutic agents [47,48]. In murine models, omega-3 FA supplementation increased cutaneous levels of pro-resolving mediators, including RvD5, protectin DX, and MaR2, while EPA reduced pro-inflammatory PGE2 and TXB2 [50]. These findings indicate that omega-3 supplementation can simultaneously enhance resolution pathways and suppress pro-inflammatory eicosanoids. RvD1 and RvD5 reduce keratinocyte-driven inflammation by downregulating IL-24 and S100A12, suggesting their potential to attenuate psoriatic plaque formation and limit immune cell recruitment in affected skin [52].
Human data on SPMs in psoriasis remain limited and derive from only two studies [51,52]. These investigations compared lipid mediator profiles in the serum and skin biopsies of patients with psoriasis versus healthy controls, and additionally assessed differences between lesional and non-lesional skin within affected individuals. Within psoriatic lesions, an increase in DHA–derived metabolites (14-HDHA and 17-HDHA) was observed, suggesting partial activation of DHA-associated resolution pathways [52]. Conversely, metabolites originating from EPA, such as 14,15-epoxyeicosatetraenoic acid (14,15-EpETE) and 17,18-epoxyeicosatetraenoic acid (17,18-EpETE), were scarcely detectable or absent [51]. This pattern suggests a disruption or insufficiency in the EPA-derived lipid mediator pathway, leading to an incomplete activation of resolution-phase processes. Several omega-3 and omega-6 metabolites found in the skin were also detected in the bloodstream, although their differences were less marked. Higher serum hydroxyeicosatetraenoic acid (HETE) levels, an oxylipin derived from AA, in healthy individuals and comparable RvD2 detection across groups suggest that lipid mediator dysregulation in psoriasis is largely confined to the skin, making serum not representative for local inflammatory activity [51]. Clinically, localized disruption of lipid mediator homeostasis may sustain chronic skin inflammation despite modest systemic involvement. Thus, circulating biomarkers may have limited diagnostic value, while interventions that restore lipid balance—such as omega-3 supplementation, LOX/COX modulation, or correction of lipid oxidation and bile acid metabolism—could support resolution of psoriatic inflammation.
In summary, current evidence underscores the pivotal yet underexplored role of specialized SPMs and omega-3–derived lipids in psoriasis. Alterations in cutaneous lipid mediator networks—characterized by a relative deficiency of omega-3–derived pro-resolving metabolites and dominance of omega-6–driven inflammation—appear to impair resolution pathways and perpetuate chronic skin inflammation. These findings highlight the therapeutic potential of targeting lipid metabolism and enhancing SPMs biosynthesis as complementary strategies for restoring immune and epidermal homeostasis in psoriasis.
4.5. Hidradenitis Suppurativa
HS is a chronic, relapsing inflammatory skin disease characterized by a pathogenic triad of aberrant immune activation, dysregulated metabolic signaling, and impaired epithelial differentiation [153]. It predominantly affects intertriginous areas, including the axillae, groin, inframammary, and perianal regions, where it manifests as recurrent painful nodules, abscesses, and draining sinus tracts [154]. The disease follows a debilitating course and profoundly impacts multiple aspects of patients’ lives, including occupational functioning, social interactions, and sexual health, often resulting in significant psychosocial distress and social isolation [155]. HS is frequently associated with a range of systemic comorbidities, most notably components of metabolic syndrome. Estimates suggest that up to half of all patients with HS meet the diagnostic criteria for metabolic syndrome, including obesity, hypertension, dyslipidemia, and insulin resistance [156]. The chronic inflammatory state contributes to a markedly increased overall health burden. Consequently, individuals with HS experience a substantially reduced life expectancy, with some studies reporting a decrease of approximately 15 years compared to the general population [157].
This elevated morbidity and mortality underscore the systemic nature of the disease. HS is characterized by a dominant Th1/Th17 immune response, with lesional skin exhibiting elevated levels of pro-inflammatory cytokines including TNF-α, IL-1β, IL-6, IL-17, and IL-23 [154]. Existing evidence indicates that HS is characterized by both systemic lipid dysregulation and a localized imbalance of cutaneous lipid mediators. Lesional skin exhibits impaired regulation of sphingolipid-metabolizing enzymes, with a shift toward catabolic pathways [157]. Recent lipidomic analyses in HS have revealed systemic alterations in lipid metabolism, particularly dysregulation of sphingolipid pathways, and elevated levels of pro-inflammatory oxylipins—such as HETEs and DHETs—correlating with disease activity [158]. Although systemic changes were less pronounced than those observed in lesional skin [159], the study’s methodological design ensured that the detected changes reflected a disease-specific lipid imbalance independent of metabolic comorbidities.
To date, direct evidence concerning SPMs in HS is lacking; therefore, this section aims to summarize the available data on pro-inflammatory lipid mediators and identify existing gaps in the current literature. Lipidomic profiling of lesional HS skin revealed a distinct pro-inflammatory shift, characterized by elevated levels of 5-LOX–derived metabolites from AA—notably 5-HETE, 5-oxo-eicosatetraenoic acid (5-oxo-ETE), and leukotriene B4 (LTB4) —accompanied by reduced concentrations of anti-inflammatory products generated via 12-LOX and 15-LOX pathways from EPA and DHA [133]. Although the study did not directly measure SPMs, the observed reduction in 15-hydroxyeicosapentaenoic acid (15-HEPE)—a key precursor of E-series resolvins—indirectly suggests a diminished capacity for SPMs biosynthesis. This deficiency may contribute to the chronic, non-resolving Th1/Th17-driven inflammatory phenotype characteristic of HS, while highlighting the potential underrepresentation of pro-resolving pathways in the lesional lipidome. Taken together, these findings underscore a clear pro-inflammatory lipid signature in HS but also point to several important gaps in knowledge that require further investigation. First, the detailed composition of FAs in both lesional and circulating compartments of HS patients—particularly the balance between omega-3 and omega-6 species—remains poorly characterized. Second, the specific contributions of pro-resolving lipid mediators to HS pathophysiology have been largely unexplored, with the majority of existing data focusing on pro-inflammatory eicosanoids. Third, it remains unclear whether lipid dysregulation acts as a primary driver of HS pathogenesis or arises secondarily as a consequence of chronic inflammation. Finally, therapeutic strategies targeting lipid pathways—such as dietary modulation of FAs or pharmacologic manipulation of lipid mediators—are urgently needed, given that outcomes achieved with currently available treatments, which primarily focus on the inhibition of pro-inflammatory cytokines, remain largely unsatisfactory.
4.6. Acne Vulgaris
Acne vulgaris is a common chronic dermatological condition involving the pilosebaceous unit. It is one of the most prevalent diseases worldwide, affecting approximately 9.4% of the global population [160], and its incidence appears to be increasing [74]. The condition may manifest with non-inflammatory lesions (open and closed comedones) as well as inflammatory lesions (papules, pustules, and nodules) [161]. Although acne typically begins during adolescence and predominantly affects young males, it may also develop in adults, among whom it is more frequently observed in women. The pathogenesis of acne involves increased follicular keratinization, excess production of sebum, inflammation, and colonization of the hair follicles by Cutibacterium acnes [161]. Genetic and ethnic factors influence acne susceptibility; however, increasing incidence has been associated with the adoption of Western lifestyles, which are characterized by low intake of PUFAs and high consumption of saturated fats and refined carbohydrates [162]. Acne affects daily functioning, causing feelings of embarrassment, anxiety, and discomfort [163].
Initially, inflammation was described as occurring in the later stages of acne, accompanying clinical lesions such as papules and pustules. More recent studies, however, indicate that inflammation is also present in the early stages of acne development [77,164]. Moreover, evidence suggests that inflammation may precede hyperkeratinization of the pilosebaceous unit. This is supported by the observation of elevated levels of CD3+ and CD4+ T cells in the perifollicular and papillary dermis, as well as an increased number of macrophages in clinically unaffected skin in acne patients [165,166].
Scientific studies indicate possible lipid abnormalities in patients with acne. Sobhan et al. demonstrated significantly higher total cholesterol levels, particularly among male patients with acne [167]. Other lipid parameters (triglicerydes, LDL, and HDL) were higher in the acne group than in controls, but the differences were not statistically significant. Jiang et al. reported significantly elevated levels of total cholesterol, LDL, and lipoprotein A in acne patients compared to control groups, with male patients also showing higher triglyceride levels [168]. Research suggests that certain lipid components and their ratios—particularly the omega-6/omega-3 and PUFA/MUFA ratios—may have a causal influence on acne risk [137]. Gokdemir et al. found statistically higher serum triglyceride concentrations in patients with acne [169]. Although the existing data present inconsistent findings, they collectively highlight potential directions for future large-scale investigations.
The enzyme lipoprotein lipase (LPL) catalyzes the hydrolysis of triglycerides in plasma triglyceride-rich lipoproteins at the surface of capillary endothelial cells. Its expression has also been detected in skin appendages, implying that sebaceous glands possess the ability to sequester FAs, which may be associated with the pathogenesis of acne [73]. The angiopoietin-like proteins (ANGPTLs) 3 and 4 are the main regulators of plasma lipid metabolism due to their inhibitory effect on LPL. Secretory PLA2, hydrolyzes phospholipids, generating free FAs and lysophospholipids, which exacerbate the inflammation [73]. Aslan et al. demonstrated increased activity of sPLA2 and elevated AA/EPA and dihomylo-γ-gamma-linoleic acid/eicosapentaenoic acid (DGLA/EPA) ratios in patients with acne. Significantly higher activity of LPL was also observed in the same cohort [73]. The activities of both enzymes in sebaceous glands generate free FAs that can fuel inflammation leading to acne pathogenesis.
Recent studies have begun to elucidate the relationship between lipids and inflammation in acne, with particular emphasis on PUFAs [73,74,75,77,160,168,169]. Among these studies, omega-3 supplementation was associated with an improvement in acne lesions [71,137,165,167], and with a beneficial effect on gut microbial diversity [76]. However, current evidence regarding PUFA-enriched diets remains limited due to small sample sizes, heterogeneous interventions, and inconsistent outcome measures. Moreover, during systemic acne therapy with isotretinoin, omega-3 FAs were shown to reduce some of the drug’s adverse effects [74]. In contrast to omega-3, the role of omega-6 FAs in acne remains unclear. γ -linoleic acid (GLA) appears to improve acne lesions [72], whereas Zhang et al. demonstrated that DGLA may exacerbate acne [77]. Additionally, AA and EPA were identified as protective factors against the development of inflammation in acne [77]. In acne vulgaris patients higher levels of AA/EPA and DGLA/EPA ratios were observed [73]. These findings suggest that a higher dietary intake of omega-3 FAs, combined with reduced consumption of saturated fats, may exert a beneficial therapeutic effect in the management of acne. Furthermore, Zhang et al. identified several SNPs within the fatty acid desaturase 1 (FADS1) and fatty acid desaturase 2 (FADS2) genes. These variants may affect the enzymatic activity of FADS1 and FADS2 in PUFA metabolism, thereby contributing to the worsening of acne symptoms [77]. Personalized approaches based on lipid and genomic profiling could therefore help optimize therapeutic outcomes in acne patients.
Acne vulgaris is associated with a mixed but predominantly Th1/Th17-type immune response, characterized by elevated levels of pro-inflammatory cytokines such as IL-1β, IL-6, IL-8, IL-17 and TNF-α, which contribute to follicular inflammation, neutrophil infiltration and lesion formation [170,171]. Emerging evidence suggests that SPMs can limit pro-inflammatory cytokine production and leukocyte infiltration, thereby promoting resolution of inflammation and offering potential therapeutic benefits in acne. So far, no direct link has been established between SPMs and acne development. However, the involvement of inflammatory processes in the early stages of acne formation and comedogenesis, together with the beneficial effects of omega-3 FAs on the course of the disease, suggests that inadequate resolution of inflammation and disturbances in lipid mediator homeostasis may contribute to the pathogenesis of acne vulgaris. While SPMs represent a promising new direction for acne research, the beneficial effects of omega-3 FAs cannot be explained solely by their conversion to SPMs. Omega-3 FAs exert multifaceted biological actions among which the bactericidal activity of long chain PUFAs against Cutibacterium acnes and Staphylococcus aureus appears particularly relevant to acne pathogenesis [39]. Moreover, omega-3 FAs provide alternative substrates for LPL and sPLA2, potentially reducing the synthesis of pro-inflammatory mediators. Thus, understanding how SPMs and PUFA-derived lipid mediators contribute to the regulation of inflammation and microbial balance in acne may open new therapeutic perspectives.
4.7. Atopic Dermatitis
Similar to psoriasis, AD is characterized by an imbalance between pro- and anti-inflammatory lipids, prompting investigation into the role of omega-3 FAs and SPMs in this condition. AD constitutes a major global dermatological health burden, affecting approximately 10–20% of children and up to 3% of adults [172]. Multifactorial pathogenesis of AD encompasses genetic predisposition, epidermal barrier dysfunction, immune dysregulation, and environmental determinants [18,101]. Of particular note, the prevalence of AD has shown a progressive increase over recent decades, a trend partly attributed to changes in dietary habits, particularly the shift toward higher consumption of omega-6 PUFAs and the resulting imbalance in the omega-6 to omega-3 ratio [173]. Growing evidence indicates that disruption of the omega-6 to omega-3 ratio contributes to dysregulated eicosanoid signaling and persistent inflammation [174].Immunologically, acute AD is primarily mediated by a Th2-dominant response, with heightened expression of IL-4, IL-5, IL-13, IL-31, and occasionally IL-22, while chronic or treatment-resistant lesions frequently involve additional activation of Th1, Th17, and Th22 pathways [175].
At the molecular level, omega-3–derived lipid mediators exert pleiotropic immunomodulatory and barrier-stabilizing effects on the cutaneous microenvironment characteristic of AD [78,80,81,82]. EPA and DHA compete with AA for COX and LOX pathways, thereby reducing the generation of pro-inflammatory eicosanoids such as PGE2 and LTB4 [81,83]. In parallel, the conversion of EPA and DHA into specialized pro-resolving lipid mediators actively terminates inflammation through the downregulation of Th2- and Th17-driven cytokines (IL-4, IL-5, IL-13, IL-17A) and the suppression of IgE synthesis [80,82]. These mediators also enhance regulatory T-cell (Treg) activity, promote macrophage M2 polarization, and restore epidermal barrier integrity by normalizing ceramide synthesis and reducing TEWL [44,80]. Given the dominant Th2 cytokine milieu in acute AD, which expands to include Th17 and Th22 responses in chronic stages, SPM-mediated downregulation of IL-4, IL-13, and other inflammatory mediators offers a promising mechanism to rebalance immune responses, support barrier repair, and potentially reduce pruritus and allergen sensitivity. Indeed, impaired generation of SPMs has been proposed as a contributing factor to the chronic, non-resolving inflammation in AD, and restoration of pro-resolving lipid mediator pathways is increasingly viewed as a potential novel therapeutic strategy [176].
Dietary modulation of PUFA intake and direct SPMs administration have been explored as therapeutic approaches for AD [78,79,81,82,83,89,90,92,93,96,97,99,101]. The following studies, summarized in Table 4, encompass both clinical and preclinical investigations exploring the influence of omega-3/omega-6 FAs and SPMs on the onset and severity of AD. These studies evaluate diverse interventions—including maternal supplementation during pregnancy, direct oral intake of EPA/DHA, topical formulations, and experimental administration of SPMs analogues—in both human cohorts and animal models. Experimental studies consistently demonstrate that EPA- and DHA-derived mediators attenuate Th2/Th17 inflammation, reduce IgE and histamine levels, and restore epidermal lipid balance, thereby improving barrier function [78,79,80,81,82,84]. These effects are further supported by data showing enhanced Treg activity and suppression of pro-inflammatory cytokines following SPMs or omega-3 administration [80,96].
In contrast, clinical findings are more heterogeneous: while several randomized controlled trials and maternal supplementation studies report reduced IgE-associated eczema [95], decreased allergen sensitization, or milder disease severity [93], others find no significant preventive or therapeutic effects [89,90,96,99]. Recent trials further substantiate both the potential and the limitations of omega-3 interventions. Supplementation with DHA/EPA in adults and children with AD has been shown to significantly reduce SCORAD scores and improve patient-reported outcomes [94,101], and maternal omega-3 intake during pregnancy was associated with a lower incidence of IgE-mediated eczema in infants [95]. In the context of SPMs and omega-3/omega-6 FAs in AD, there is emerging evidence that maternal DHA intake may modify the increased risk of AD associated with prenatal arsenic exposure [100]. However, other well-controlled studies demonstrated no reduction in eczema risk despite clear biochemical evidence of increased maternal and infant DHA/EPA levels and dampened cytokine responses [96,99]. Such discrepancies may be attributed to heterogeneity in population risk profiles, baseline dietary patterns, genetic factors, and timing or dosage of supplementation. Recent meta-analytic evidence indicates that although maternal omega-3 PUFAs supplementation during pregnancy does not significantly reduce the overall incidence of eczema in offspring, it may confer a potential subgroup benefit for IgE-associated eczema in children under three years of age, highlighting age-dependent and immunophenotype-specific effects of prenatal omega-3 exposure [177]. Overall, while omega-3 FAs exhibit immunomodulatory and barrier-stabilizing properties, the inconsistency of clinical outcomes precludes universal recommendations and highlights the multifactorial nature of AD pathophysiology.
Consequently, attention has increasingly shifted toward SPMs, which exert potent anti-inflammatory and pro-resolving actions at nanomolar concentrations [124]. Their ability to actively terminate inflammation and promote tissue homeostasis has drawn researchers’ attention to the development of stable analogues or receptor agonists for clinical application. To date, the only human study in AD utilizing an SPMs-based approach evaluated a topical 15(R/S)-methyl-lipoxin A4 cream in infants with acute facial eczema, which significantly improved erythema, pruritus, and lesion severity within days [98]. Moreover, the cream demonstrated comparable efficacy to medium-potency corticosteroids without adverse effects; however, the relapse rate at 40 days was higher than in the control group treated with mometasone [98]. These findings suggest that targeting the SPM pathway may represent a promising, mechanism-driven therapeutic strategy distinct from conventional anti-inflammatory treatments.
4.8. Integrated Summary of Findings and Overall Limitations
Across psoriasis, AD, acne, and HS, the findings of this systematic review demonstrate that disturbances in lipid mediator networks—particularly imbalances between omega-3– and omega-6–derived metabolites—are consistently associated with aggravated inflammation, impaired barrier function, and dysregulated immune responses. Hidradenitis suppurativa, psoriasis, and acne vulgaris are predominantly Th1/Th17-driven, with elevated IL-17A, IL-17F, IL-23, TNF-α, and IFN-γ promoting keratinocyte activation, neutrophil recruitment, and chronic inflammation, whereas atopic dermatitis is primarily Th2-mediated, with IL-4, IL-5, IL-13, and IL-31 driving IgE production, barrier dysfunction, and allergic sensitization. SPMs modulate immune responses in a context-dependent manner, targeting the dominant T-helper cell profile: suppressing IFN-γ and TNF-α in Th1-driven diseases, downregulating IL-4, IL-5, and IL-13 in Th2-mediated disorders, and attenuating IL-17/IL-23 signaling and neutrophil-driven inflammation in Th17-skewed conditions. Across all contexts, SPMs enhance Treg activity and promote macrophage M2 polarization, providing a conserved mechanism for inflammation resolution and tissue repair. Human data remain limited but broadly align with experimental findings, suggesting that insufficient SPMs biosynthesis or precursor availability may contribute to chronic, non-resolving inflammation. Interventional studies involving omega-3 supplementation show variable but generally favorable outcomes in psoriasis, acne, and AD, particularly regarding patient-reported symptoms. Evidence in HS is minimal and indirect. Taken together, the evidence suggests that lipid mediators derived from PUFAs act as central regulators of immune resolution in the skin.
This body of evidence has notable limitations, including the scarcity of human studies, small cohorts, and heterogeneous interventions. Many trials depend on non-standardized or subjective outcomes, and dietary studies may be influenced by confounding factors, including lifestyle and genetic susceptibility. Moreover, mechanistic in vitro and animal data may not adequately reflect human physiology, reducing translational relevance. The reliance on mechanistic and animal studies, together with the probable underrepresentation of studies reporting null or negative outcomes, limits confidence in the estimated therapeutic benefits of omega-3 fatty acids and SPMs in humans.
This review also has methodological limitations. Only one database (MEDLINE/PubMed) was searched, which may have led to missed studies published elsewhere. A major limitation of this review is the lack of a formal risk-of-bias assessment. This was due to the substantial heterogeneity and mechanistic nature of the included evidence, which comprised in vitro studies, animal models, and exploratory human research—study types for which no unified risk-assessment framework exists. Additionally, highly variable methodologies, small sample sizes, and inconsistent outcome reporting across studies prevented meta-analysis, assessment of reporting bias, and formal evaluation of certainty of evidence. Moreover, considerable variability in analytical techniques for quantifying lipid mediators—including differences in sample preparation, detection platforms, and quantification methods—further limits comparability across studies and introduces potential measurement inconsistency. Additionally, publication bias could not be evaluated, and certainty of evidence was not assessed due to the nature of the included data.
Clinically, omega-3 supplementation may offer complementary benefits in inflammatory skin diseases, although current evidence is insufficient for formal recommendations. SPM-based therapies, particularly stable analogues and receptor agonists, represent a promising yet still emerging therapeutic direction. Future research should focus on well-designed clinical trials with standardized clinical and biochemical endpoints, comprehensive lipidomic profiling, and mechanistic work on SPM analogues and agonists, as these agents offer the most realistic route toward therapeutic translation.
5. Conclusions
Dysregulated lipid metabolism—particularly imbalances in omega-3 and omega-6–derived mediators—plays a central role in the pathogenesis of psoriasis, HS, acne vulgaris and AD. SPMs act at the interface of lipid metabolism, immune regulation, and tissue repair, facilitating the active resolution of cutaneous inflammation. Accordingly, omega-3 FAs and their downstream SPMs represent promising adjunctive therapies capable of restoring inflammatory balance and promoting tissue homeostasis. While mechanistic and early clinical data on SPMs are compelling, further research is needed to comprehensively characterize SPMs profiles in inflammatory skin disorders. Additional studies are also required to evaluate the therapeutic potential of SPM analogues and receptor agonists, particularly given their superior stability and receptor specificity compared with endogenous mediators. The limited available human data suggest that deficiencies in omega-3–derived pro-resolving mediators, together with a shift toward omega-6–dominant inflammatory profiles, may contribute to disease chronicity and severity. This imbalance highlights the relevance of targeting lipid mediator pathways as a complementary strategy to conventional anti-inflammatory therapies. To sum up, omega-3 FAs and SPMs represent promising adjuncts for managing several inflammatory skin conditions through their capacity to rebalance inflammatory pathways and promote resolution pathways.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/antiox15010009/s1, The full PubMed search strategy, including MeSH terms, keywords, and Boolean operators, is provided in Supplementary File S1. Ref [178] was cited in the Supplementary Materials.
Author Contributions
Conceptualization, A.B.-B. and O.J.-Z.; methodology, A.B.-B. and H.M.; software, N/A; validation, N/A; formal analysis, N/A; investigation, N/A; resources, A.B.-B. and O.J.-Z.; data curation, A.B.-B. and O.J.-Z.; writing—original draft preparation, A.B.-B. and O.J.-Z.; writing—review and editing, H.M. and I.F.; visualization, A.B.-B. and O.J.-Z.; supervision, I.F.; project administration, H.M.; funding acquisition, N/A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Publication Fund of the Medical University of Bialystok.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data analyzed in this systematic review are publicly available from the cited publications. The full study protocol and search strategy are archived and accessible at [https://doi.org/10.17605/OSF.IO/FV9PR]. No new primary data were generated.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 14,15-EpETE/17,18-EpETE | 14,15-Epoxyeicosatetraenoic Acid/17,18-Epoxyeicosatetraenoic Acid |
| 14-HDHA/17-HDHA | 14-Hydroxydocosahexaenoic Acid/17-Hydroxydocosahexaenoic Acid |
| 5-HETE/12-HETE/12-HEPE/15-HEPE | 5-Hydroxyeicosatetraenoic Acid/12-Hydroxyeicosatetraenoic Acid/12-Hydroxyeicosapentaenoic Acid/15-Hydroxyeicosapentaenoic Acid |
| 5-oxo-ETE | 5-Oxo-Eicosatetraenoic Acid |
| AA | Arachidonic Acid |
| AC | Acne Comedonica |
| AD/AD-lika | Atopic dermatitis/Atopic dermatitis like |
| ALA | α-Linolenic Acid |
| ALX/FPR2 | Lipoxin A4 Receptor/Formyl Peptide Receptor 2 |
| AP | Acne Papulopustulosa |
| AT/ATL | Aspirin triggered/Aspirin triggered lipoxin |
| BALB/c | Bagg Albino Laboratory-bred Mouse Strain c |
| BLT1 | Leukotriene B4 Receptor 1 |
| BSA | Body Surface Area |
| CBMC | Cord blood mononuclear cell |
| CCL2 | C-C motif chemokine ligand 2 |
| CD80/86 | Co-stimulatory molecules expressed on antigen-presenting cells |
| CI | Confidence Interval |
| COX/COX-2/acetyl-COX-2 | Cyclooxygenase/Cyclooxygenase-2/aspirin-acetylated cyclooxygenase-2 |
| CRP | C-Reactive Protein |
| CTLA-4 | Cytotoxic T-lymphocyte-associated protein 4 |
| CYP-450 | Cytochrome P450 Enzyme System |
| CXCL10 | C-X-C motif chemokine ligand 10 |
| DGLA | Dihomo-γ-Linolenic Acid |
| DHA | Docosahexaenoic Acid |
| DHA-M2 | Docosahexaenoic acid–induced M2 macrophages |
| DHET | Dihydroxyeicosatrienoic Acid |
| DLQI | Dermatology Life Quality Index |
| DNCB | 2,4-dinitrochlorobenzene |
| DNFB | 2,4-dinitrofluorobenzene |
| DPA | Docosapentaenoic Acid |
| EASI | Eczema Area and Severity Index |
| ECM | Extracellular Matrix |
| ELOVL | Elongation of Very Long Chain Fatty Acids Protein (family) |
| EPA/EPA-E | Eicosapentaenoic Acid/eicosapentaenoic acid ethyl ester |
| ERK1/2 | Extracellular Signal–Regulated Kinases 1/2 |
| FAs | Fatty acids |
| FABP | Fatty Acid–Binding Protein |
| FADS2 | Fatty acid desaturase 2 |
| FDLQI | Family Dermatology Life Quality Index |
| FFA4R/GPR120 | Free Fatty Acid Receptor 4 |
| FK506 | Tacrolimus |
| FMT | Fecal Microbiota Transplantation |
| GLA | γ-linolenic acid |
| GPCR | G Protein–Coupled Receptor |
| HDL | High-density lipoprotein |
| HETE | Hydroxyeicosatetraenoic Acid |
| HEPE | Hydroxy-eicosapentaenoic Acid |
| HMGB | High-Mobility Group Box protein |
| HpEPE | Hydroperoxy-eicosapentaenoic Acid |
| HpETE | Hydroxyperoxy-eicosatetraenoic Acid |
| HR-AD | Hairless atopic dermatitis |
| HRO | Herring Roe Oil |
| HS | Hidradenitis Suppurativa |
| IgE | Immunoglobulin E |
| IL | Interleukin |
| IRI/I–R | Ischemia–Reperfusion Injury |
| ISAAC | International Study of Asthma and Allergies in Childhood |
| IV | Intravenous |
| IMQ | Imiquimod |
| KO | Knockout |
| LCE3B–C | Late Cornified Envelope Genes 3B and 3C |
| LC-PUFA | Long-chain polyunsaturated fatty acid |
| LN | Lymph node |
| LOX/5-LOX/12-LOX/15-LOX | Lipoxygenase/5- Lipoxygenase/12- Lipoxygenase/15- Lipoxygenase |
| LPL | Lipoprotein Lipase |
| LPS | Lipopolysaccharide |
| LT/LTA4/5/LTE4/5 | Leukotriene A4/5—E4/5 |
| LXA4/LXA5 | Lipoxin A4/A5 |
| M2 | Alternatively Activated Macrophage Phenotype |
| MAPK | Mitogen-Activated Protein Kinase |
| MaR1/MaR2 | Maresin-1/Maresin 2 |
| MCT | Medium-chain triglyceride |
| MCTR | Maresin Conjugates in Tissue Regeneration |
| MD | Mean difference |
| MR | Mendelian Randomization |
| MUFA | Monounsaturated fatty acid |
| NF-κB | Nuclear Factor kappa-light-chain-enhancer of activated B cells |
| NHDF | Normal Human Dermal Fibroblasts |
| NHEK | Normal Human Epidermal Keratinocytes |
| NK | Natural Killer Cells |
| NRF2 | Nuclear Factor Erythroid 2–Related Factor 2 |
| NSAID | Non-Steroidal Anti-Inflammatory Drug |
| PASI | Psoriasis Area and Severity Index |
| PAF | Platelet-activating factor |
| PBMC | peripheral blood mononuclear cell |
| PG/PGE2/PGE3/PGH2/PGG2 | Prostaglandin/E2/E3/H2/G2 |
| PGI2 | Prostacyclin |
| PSO | Psoriasis |
| PUFA | Polyunsaturated Fatty Acid |
| PD1 | Protectin D1 |
| PDX | Protectin DX |
| PLA2/PLA2G4E/sPLA2 | Phospholipase A2/Phospholipase A2 Group IVE/Secretory Phospholipase A2 |
| PPAR | Peroxisome Proliferator–Activated Receptor |
| PRS | Polygenic risk score |
| RCT | Randomized Controlled Trial |
| QoL | Quality of life |
| RvD1-RvD6 | Resolvin D1-D6 |
| RvE/RvE1–RvE4 | Resolvin E-series/E1/E2/E3/E4 |
| S100A12 | S100 Calcium-Binding Protein A12 |
| SCORAD | SCORing Atopic Dermatitis |
| SFA | Saturated fatty acid |
| SNP | Single-nucleotide polymorphism |
| SPMs | Specialized Pro-Resolving Mediators |
| SSS | Symptom Severity Score |
| STAT1 | Signal Transducer and Activator of Transcription 1 |
| TCM | Central Memory T cells |
| TCS | Topical corticosteroid |
| TEWL | Transepidermal Water Loss |
| TG | Trigliceride |
| TGF-β | Transforming growth factor-β |
| Th/Th2/Th17 | Helper T Cell/Type 2 Helper T Cell/Type 17 Helper T Cell |
| TN | Naïve T cells |
| TNF-α | Tumor Necrosis Factor α |
| Treg | Regulatory T cell |
| TSLP | Thymic stromal lymphopoietin |
| TXA2/TXB2 | Thromboxane A2/B2 |
| UFA | Unsaturated Fatty Acid |
| UV | Ultraviolet |
| VDR | Vitamin D Receptor |
| WT | Wild-Type |
| ω-3 | Omega-3 Fatty Acids |
| ω-6 | Omega-6 Fatty Acids |
References
- Serhan, C.N.; Savill, J. Resolution of Inflammation: The Beginning Programs the End. Nat. Immunol. 2005, 6, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Clish, C.B.; Brannon, J.; Colgan, S.P.; Chiang, N.; Gronert, K. Novel Functional Sets of Lipid-Derived Mediators with Antiinflammatory Actions Generated from Omega-3 Fatty Acids via Cyclooxygenase 2-Nonsteroidal Antiinflammatory Drugs and Transcellular Processing. J. Exp. Med. 2000, 192, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Maddox, J.F.; Petasis, N.A.; Akritopoulou-Zanze, I.; Papayianni, A.; Brady, H.R.; Colgan, S.P.; Madara, J.L. Design of Lipoxin A4 Stable Analogs That Block Transmigration and Adhesion of Human Neutrophils. Biochemistry 1995, 34, 14609–14615. [Google Scholar] [CrossRef] [PubMed]
- Dalli, J.; Serhan, C. Macrophage Proresolving Mediators-the When and Where. Microbiol. Spectr. 2016, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. A Search for Endogenous Mechanisms of Anti-Inflammation Uncovers Novel Chemical Mediators: Missing Links to Resolution. Histochem. Cell Biol. 2004, 122, 305–321. [Google Scholar] [CrossRef]
- Fierro, I.M.; Colgan, S.P.; Bernasconi, G.; Petasis, N.A.; Clish, C.B.; Arita, M.; Serhan, C.N. Lipoxin A4 and Aspirin-Triggered 15-Epi-Lipoxin A4 Inhibit Human Neutrophil Migration: Comparisons Between Synthetic 15 Epimers in Chemotaxis and Transmigration with Microvessel Endothelial Cells and Epithelial Cells. J. Immunol. 2003, 170, 2688–2694. [Google Scholar] [CrossRef]
- Hosseini, Z.; Marinello, M.; Decker, C.; Sansbury, B.E.; Sadhu, S.; Gerlach, B.D.; Bossardi Ramos, R.; Adam, A.P.; Spite, M.; Fredman, G. Resolvin D1 Enhances Necroptotic Cell Clearance Through Promoting Macrophage Fatty Acid Oxidation and Oxidative Phosphorylation. Arter. Thromb. Vasc. Biol. 2021, 41, 1062–1075. [Google Scholar] [CrossRef]
- Chiang, N.; Libreros, S.; Norris, P.C.; De La Rosa, X.; Serhan, C.N. Maresin 1 Activates LGR6 Receptor Promoting Phagocyte Immunoresolvent Functions. J. Clin. Investig. 2019, 129, 5294–5311. [Google Scholar] [CrossRef]
- Fredman, G.; Khan, S. Specialized Pro-resolving Mediators Enhance the Clearance of Dead Cells. Immunol. Rev. 2023, 319, 151–157. [Google Scholar] [CrossRef]
- Qiao, N.; Lin, Y.; Wang, Z.; Chen, J.-Y.; Ge, Y.-Y.; Yao, S.-L.; Gong, J. Maresin1 Promotes M2 Macrophage Polarization Through Peroxisome Proliferator–Activated Receptor-γ Activation to Expedite Resolution of Acute Lung Injury. J. Surg. Res. 2020, 256, 584–594. [Google Scholar] [CrossRef]
- Basil, M.C.; Levy, B.D. Specialized Pro-Resolving Mediators: Endogenous Regulators of Infection and Inflammation. Nat. Rev. Immunol. 2016, 16, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Perez-Hernandez, J.; Chiurchiù, V.; Perruche, S.; You, S. Regulation of T-Cell Immune Responses by Pro-Resolving Lipid Mediators. Front. Immunol. 2021, 12, 768133. [Google Scholar] [CrossRef] [PubMed]
- Ramon, S.; Bancos, S.; Serhan, C.N.; Phipps, R.P. Lipoxin A4 Modulates Adaptive Immunity by Decreasing Memory B-cell Responses via an ALX/FPR 2-dependent Mechanism. Eur. J. Immunol. 2014, 44, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Duffney, P.F.; Falsetta, M.L.; Rackow, A.R.; Thatcher, T.H.; Phipps, R.P.; Sime, P.J. Key Roles for Lipid Mediators in the Adaptive Immune Response. J. Clin. Investig. 2018, 128, 2724–2731. [Google Scholar] [CrossRef]
- Jin, Z.; Song, Y.; He, L. A Review of Skin Immune Processes in Acne. Front. Immunol. 2023, 14, 1324930. [Google Scholar] [CrossRef]
- Schön, M.P. Adaptive and Innate Immunity in Psoriasis and Other Inflammatory Disorders. Front. Immunol. 2019, 10, 1764. [Google Scholar] [CrossRef]
- Burke, O.M.; Frerichs, V.R.; Garcia, D.F.; Stone, R.C.; Lev-Tov, H.; Czarnowicki, T.; Keane, R.W.; Ojeh, N.; Marjanovic, J.; Pastar, I.; et al. The Impact of Innate Immunity and Epigenetics in the Pathogenesis of Hidradenitis Suppurativa. Front. Immunol. 2025, 16, 1593253. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, Y.; Xu, M.; Zhong, M.; Peng, X.; Zeng, K.; Huang, X. The Roles of Innate Immune Cells in Atopic Dermatitis. J. Innate Immun. 2024, 16, 385–396. [Google Scholar] [CrossRef]
- Orzan, O.A.; Tutunaru, C.V.; Ianoși, S.L. Understanding the Intricate Pathophysiology of Psoriasis and Related Skin Disorders. Int. J. Mol. Sci. 2025, 26, 749. [Google Scholar] [CrossRef]
- Spiteri, J.; Mintoff, D.; Grech, L.; Pace, N.P. Transcriptomic Signatures and Molecular Pathways in Hidradenitis Suppurativa—A Narrative Review. Int. J. Mol. Sci. 2025, 26, 7704. [Google Scholar] [CrossRef]
- Furue, M.; Furue, K.; Tsuji, G.; Nakahara, T. Interleukin-17A and Keratinocytes in Psoriasis. Int. J. Mol. Sci. 2020, 21, 1275. [Google Scholar] [CrossRef] [PubMed]
- Schell, S.L.; Cong, Z.; Sennett, M.L.; Gettle, S.L.; Longenecker, A.L.; Goldberg, S.R.; Kirby, J.S.; Helm, M.F.; Nelson, A.M. Keratinocytes and Immune Cells in the Epidermis Are Key Drivers of Inflammation in Hidradenitis Suppurativa Providing a Rationale for Novel Topical Therapies. Br. J. Dermatol. 2023, 188, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Chieosilapatham, P.; Kiatsurayanon, C.; Umehara, Y.; Trujillo-Paez, J.V.; Peng, G.; Yue, H.; Nguyen, L.T.H.; Niyonsaba, F. Keratinocytes: Innate Immune Cells in Atopic Dermatitis. Clin. Exp. Immunol. 2021, 204, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Orsmond, A.; Bereza-Malcolm, L.; Lynch, T.; March, L.; Xue, M. Skin Barrier Dysregulation in Psoriasis. Int. J. Mol. Sci. 2021, 22, 10841. [Google Scholar] [CrossRef] [PubMed]
- Andersen, A.D.; Almegaard, A.; Schultz, A.N.; Zibert, J.R. Effect of Omega-3 Supplementation on Quality of Life in Patients with Psoriasis: A Digital Survey-Based Study. Acta Derm. Venereol. 2019, 99, 907–908. [Google Scholar] [CrossRef] [PubMed]
- Jerzyńska, A.; Polańska, A.; Trafalska, E.; Jankowska, A.; Podlecka, D.; Brzozowska, A. Prenatal Polyunsaturated Fatty Acids and Atopic Dermatitis and Food Allergy in Children from Polish Mother and Child Cohort Study. Int. J. Occup. Med. Environ. Health 2023, 36, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Sasaki, S.; Tanaka, K.; Ohya, Y.; Matsunaga, I.; Yoshida, T.; Hirota, Y.; Oda, H. Relationship between Dietary Fat and Fish Intake and the Prevalence of Atopic Eczema in Pregnant Japanese Females: Baseline Data from the Osaka Maternal and Child Health Study. Asia Pac. J. Clin. Nutr. 2008, 17, 612–619. [Google Scholar]
- Nwaru, B.I.; Erkkola, M.; Lumia, M.; Kronberg-Kippilä, C.; Ahonen, S.; Kaila, M.; Ilonen, J.; Simell, O.; Knip, M.; Veijola, R.; et al. Maternal Intake of Fatty Acids during Pregnancy and Allergies in the Offspring. Br. J. Nutr. 2012, 108, 720–732. [Google Scholar] [CrossRef]
- Trak-Fellermeier, M.A.; Brasche, S.; Winkler, G.; Koletzko, B.; Heinrich, J. Food and Fatty Acid Intake and Atopic Disease in Adults. Eur. Respir. J. 2004, 23, 575–582. [Google Scholar] [CrossRef]
- Weiland, S.K.; von Mutius, E.; Hüsing, A.; Asher, M.I. Intake of Trans Fatty Acids and Prevalence of Childhood Asthma and Allergies in Europe. ISAAC Steering Committee. Lancet 1999, 353, 2040–2041. [Google Scholar] [CrossRef]
- Collier, P.M.; Ursell, A.; Zaremba, K.; Payne, C.M.; Staughton, R.C.; Sanders, T. Effect of Regular Consumption of Oily Fish Compared with White Fish on Chronic Plaque Psoriasis. Eur. J. Clin. Nutr. 1993, 47, 251–254. [Google Scholar] [PubMed]
- Dotterud, C.K.; Storrø, O.; Simpson, M.R.; Johnsen, R.; Øien, T. The Impact of Pre- and Postnatal Exposures on Allergy Related Diseases in Childhood: A Controlled Multicentre Intervention Study in Primary Health Care. BMC Public Health 2013, 13, 123. [Google Scholar] [CrossRef] [PubMed]
- Fabbrocini, G.; Cameli, N.; Lorenzi, S.; De Padova, M.P.; Marasca, C.; Izzo, R.; Monfrecola, G. A Dietary Supplement to Reduce Side Effects of Oral Isotretinoin Therapy in Acne Patients. G. Ital. Dermatol. Venereol. 2014, 149, 441–445. [Google Scholar] [PubMed]
- Li, Y.; Li, X. PUFA, Psoriasis and Atherosclerotic CVD: Evidence from the National Health and Nutrition Examination Survey Database 2003–2006 and 2009–2014. Br. J. Nutr. 2025, 133, 1331–1341. [Google Scholar] [CrossRef]
- Mihrshahi, S.; Peat, J.K.; Webb, K.; Tovey, E.R.; Marks, G.B.; Mellis, C.M.; Leeder, S.R. The Childhood Asthma Prevention Study (CAPS): Design and Research Protocol of a Randomized Trial for the Primary Prevention of Asthma. Control Clin. Trials 2001, 22, 333–354. [Google Scholar] [CrossRef]
- Ordnung, M.; Mank, M.; Stahl, B.; Kurz, D.; Marosvölgyi, T.; Decsi, T.; Rothenbacher, D.; Genuneit, J.; Siziba, L.P. Potential Sex Differences in Human Milk Fatty Acids and Their Association with Atopic Dermatitis: Results of the Ulm SPATZ Health Study. Pediatr. Allergy Immunol. 2023, 34, e13992. [Google Scholar] [CrossRef]
- Thijs, C.; Müller, A.; Rist, L.; Kummeling, I.; Snijders, B.E.P.; Huber, M.; van Ree, R.; Simões-Wüst, A.P.; Dagnelie, P.C.; van den Brandt, P.A. Fatty Acids in Breast Milk and Development of Atopic Eczema and Allergic Sensitisation in Infancy. Allergy 2011, 66, 58–67. [Google Scholar] [CrossRef]
- Wright, S.; Bolton, C. Breast Milk Fatty Acids in Mothers of Children with Atopic Eczema. Br. J. Nutr. 1989, 62, 693–697. [Google Scholar] [CrossRef]
- Desbois, A.P.; Lawlor, K.C. Antibacterial Activity of Long-Chain Polyunsaturated Fatty Acids against Propionibacterium Acnes and Staphylococcus Aureus. Mar. Drugs 2013, 11, 4544–4557. [Google Scholar] [CrossRef]
- Sarandi, E.; Krueger-Krasagakis, S.; Tsoukalas, D.; Evangelou, G.; Sifaki, M.; Kyriakakis, M.; Paramera, E.; Papakonstantinou, E.; Rudofsky, G.; Tsatsakis, A. Novel Fatty Acid Biomarkers in Psoriasis and the Role of Modifiable Factors: Results from the METHAP Clinical Study. Biomolecules 2024, 14, 1114. [Google Scholar] [CrossRef]
- Wang, X.; Ma, R.; Shi, R.; Qin, H.; Chen, W.; Yu, Z.; Ding, Y.; Peng, C.; Shi, Y. Sex Differences in the Association between Plasma Polyunsaturated Fatty Acids Levels and Moderate-to-Severe Plaque Psoriasis Severity: A Cross-Sectional and Longitudinal Study. J. Transl. Med. 2023, 21, 834. [Google Scholar] [CrossRef] [PubMed]
- Rubin, M.G.; Kim, K.; Logan, A.C. Acne Vulgaris, Mental Health and Omega-3 Fatty Acids: A Report of Cases. Lipids Health Dis. 2008, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Ashcroft, F.J.; Mahammad, N.; Midtun Flatekvål, H.; Jullumstrø Feuerherm, A.; Johansen, B. cPLA2α Enzyme Inhibition Attenuates Inflammation and Keratinocyte Proliferation. Biomolecules 2020, 10, 1402. [Google Scholar] [CrossRef] [PubMed]
- Omland, S.H.; Habicht, A.; Damsbo, P.; Wilms, J.; Johansen, B.; Gniadecki, R. A Randomized, Double-Blind, Placebo-Controlled, Dose-Escalation First-in-Man Study (Phase 0) to Assess the Safety and Efficacy of Topical Cytosolic Phospholipase A2 Inhibitor, AVX001, in Patients with Mild to Moderate Plaque Psoriasis. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 1161–1167. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Takahara, M.; Xie, L.; Takeuchi, S.; Tu, Y.; Nakahara, T.; Uchi, H.; Moroi, Y.; Furue, M. Lipoxin A(4), a Potential Anti-Inflammatory Drug Targeting the Skin. J. Dermatol. Sci. 2011, 62, 67–69. [Google Scholar] [CrossRef]
- Liu, X.; Wang, X.; Duan, X.; Poorun, D.; Xu, J.; Zhang, S.; Gan, L.; He, M.; Zhu, K.; Ming, Z.; et al. Lipoxin A4 and Its Analog Suppress Inflammation by Modulating HMGB1 Translocation and Expression in Psoriasis. Sci. Rep. 2017, 7, 7100. [Google Scholar] [CrossRef]
- Xu, J.; Duan, X.; Hu, F.; Poorun, D.; Liu, X.; Wang, X.; Zhang, S.; Gan, L.; He, M.; Zhu, K.; et al. Resolvin D1 Attenuates Imiquimod-Induced Mice Psoriasiform Dermatitis through MAPKs and NF-κB Pathways. J. Dermatol. Sci. 2018, 89, 127–135. [Google Scholar] [CrossRef]
- Sawada, Y.; Honda, T.; Nakamizo, S.; Otsuka, A.; Ogawa, N.; Kobayashi, Y.; Nakamura, M.; Kabashima, K. Resolvin E1 Attenuates Murine Psoriatic Dermatitis. Sci. Rep. 2018, 8, 11873. [Google Scholar] [CrossRef]
- Park, K.-D.; Kim, N.; Kang, J.; Dhakal, H.; Kim, J.Y.; Jang, Y.H.; Lee, W.J.; Lee, S.-J.; Kim, S.-H. Protectin D1 Reduces Imiquimod-Induced Psoriasiform Skin Inflammation. Int. Immunopharmacol. 2021, 98, 107883. [Google Scholar] [CrossRef]
- Sorokin, A.V.; Arnardottir, H.; Svirydava, M.; Ng, Q.; Baumer, Y.; Berg, A.; Pantoja, C.J.; Florida, E.M.; Teague, H.L.; Yang, Z.-H.; et al. Comparison of the Dietary Omega-3 Fatty Acids Impact on Murine Psoriasis-like Skin Inflammation and Associated Lipid Dysfunction. J. Nutr. Biochem. 2023, 117, 109348. [Google Scholar] [CrossRef]
- Sorokin, A.V.; Domenichiello, A.F.; Dey, A.K.; Yuan, Z.-X.; Goyal, A.; Rose, S.M.; Playford, M.P.; Ramsden, C.E.; Mehta, N.N. Bioactive Lipid Mediator Profiles in Human Psoriasis Skin and Blood. J. Investig. Dermatol. 2018, 138, 1518–1528. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, A.V.; Norris, P.C.; English, J.T.; Dey, A.K.; Chaturvedi, A.; Baumer, Y.; Silverman, J.; Playford, M.P.; Serhan, C.N.; Mehta, N.N. Identification of Proresolving and Inflammatory Lipid Mediators in Human Psoriasis. J. Clin. Lipidol. 2018, 12, 1047–1060. [Google Scholar] [CrossRef] [PubMed]
- Karrys, A.; Rady, I.; Chamcheu, R.-C.N.; Sabir, M.S.; Mallick, S.; Chamcheu, J.C.; Jurutka, P.W.; Haussler, M.R.; Whitfield, G.K. Bioactive Dietary VDR Ligands Regulate Genes Encoding Biomarkers of Skin Repair That Are Associated with Risk for Psoriasis. Nutrients 2018, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Wannick, M.; Bezdek, S.; Guillen, N.; Thieme, M.; Meshrkey, F.; Mousavi, S.; Seeling, M.; Nimmerjahn, F.; Mócsai, A.; Zillikens, D.; et al. Oral Administration of the Selective GPR120/FFA4 Agonist Compound A Is Not Effective in Alleviating Tissue Inflammation in Mouse Models of Prototypical Autoimmune Diseases. Pharmacol. Res. Perspect. 2018, 6, e00438. [Google Scholar] [CrossRef]
- Morin, S.; Simard, M.; Flamand, N.; Pouliot, R. Biological Action of Docosahexaenoic Acid in a 3D Tissue-Engineered Psoriatic Skin Model: Focus on the PPAR Signaling Pathway. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 159032. [Google Scholar] [CrossRef]
- Simard, M.; Rioux, G.; Morin, S.; Martin, C.; Guérin, S.L.; Flamand, N.; Julien, P.; Fradette, J.; Pouliot, R. Investigation of Omega-3 Polyunsaturated Fatty Acid Biological Activity in a Tissue-Engineered Skin Model Involving Psoriatic Cells. J. Investig. Dermatol. 2021, 141, 2391–2401.e13. [Google Scholar] [CrossRef]
- Son, S.-E.; Koh, J.-M.; Im, D.-S. Free Fatty Acid Receptor 4 (FFA4) Activation Ameliorates Imiquimod-Induced Psoriasis in Mice. Int. J. Mol. Sci. 2022, 23, 4482. [Google Scholar] [CrossRef]
- Morin, S.; Tremblay, A.; Dumais, E.; Julien, P.; Flamand, N.; Pouliot, R. Eicosapentaenoic Acid Influences the Lipid Profile of an In Vitro Psoriatic Skin Model Produced with T Cells. Biomolecules 2023, 13, 1413. [Google Scholar] [CrossRef]
- Gupta, A.K.; Ellis, C.N.; Tellner, D.C.; Anderson, T.F.; Voorhees, J.J. Double-Blind, Placebo-Controlled Study to Evaluate the Efficacy of Fish Oil and Low-Dose UVB in the Treatment of Psoriasis. Br. J. Dermatol. 1989, 120, 801–807. [Google Scholar] [CrossRef]
- Dewsbury, C.E.; Graham, P.; Darley, C.R. Topical Eicosapentaenoic Acid (EPA) in the Treatment of Psoriasis. Br. J. Dermatol. 1989, 120, 581. [Google Scholar] [CrossRef]
- Kojima, T.; Terano, T.; Tanabe, E.; Okamoto, S.; Tamura, Y.; Yoshida, S. Long-Term Administration of Highly Purified Eicosapentaenoic Acid Provides Improvement of Psoriasis. Dermatologica 1991, 182, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Grimminger, F.; Mayser, P.; Papavassilis, C.; Thomas, M.; Schlotzer, E.; Heuer, K.U.; Führer, D.; Hinsch, K.D.; Walmrath, D.; Schill, W.B. A Double-Blind, Randomized, Placebo-Controlled Trial of n-3 Fatty Acid Based Lipid Infusion in Acute, Extended Guttate Psoriasis. Rapid Improvement of Clinical Manifestations and Changes in Neutrophil Leukotriene Profile. Clin. Investig. 1993, 71, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Søyland, E.; Funk, J.; Rajka, G.; Sandberg, M.; Thune, P.; Rustad, L.; Helland, S.; Middelfart, K.; Odu, S.; Falk, E.S. Effect of Dietary Supplementation with Very-Long-Chain n-3 Fatty Acids in Patients with Psoriasis. N. Engl. J. Med. 1993, 328, 1812–1816. [Google Scholar] [CrossRef] [PubMed]
- Henneicke-von Zepelin, H.H.; Mrowietz, U.; Färber, L.; Bruck-Borchers, K.; Schober, C.; Huber, J.; Lutz, G.; Kohnen, R.; Christophers, E.; Welzel, D. Highly Purified Omega-3-Polyunsaturated Fatty Acids for Topical Treatment of Psoriasis. Results of a Double-Blind, Placebo-Controlled Multicentre Study. Br. J. Dermatol. 1993, 129, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Mayser, P.; Mrowietz, U.; Arenberger, P.; Bartak, P.; Buchvald, J.; Christophers, E.; Jablonska, S.; Salmhofer, W.; Schill, W.B.; Krämer, H.J.; et al. Omega-3 Fatty Acid-Based Lipid Infusion in Patients with Chronic Plaque Psoriasis: Results of a Double-Blind, Randomized, Placebo-Controlled, Multicenter Trial. J. Am. Acad. Dermatol. 1998, 38, 539–547. [Google Scholar] [CrossRef]
- Danno, K.; Sugie, N. Combination Therapy with Low-Dose Etretinate and Eicosapentaenoic Acid for Psoriasis Vulgaris. J. Dermatol. 1998, 25, 703–705. [Google Scholar] [CrossRef] [PubMed]
- Guida, B.; Napoleone, A.; Trio, R.; Nastasi, A.; Balato, N.; Laccetti, R.; Cataldi, M. Energy-Restricted, n-3 Polyunsaturated Fatty Acids-Rich Diet Improves the Clinical Response to Immuno-Modulating Drugs in Obese Patients with Plaque-Type Psoriasis: A Randomized Control Clinical Trial. Clin. Nutr. 2014, 33, 399–405. [Google Scholar] [CrossRef]
- Kristensen, S.; Schmidt, E.B.; Schlemmer, A.; Rasmussen, C.; Johansen, M.B.; Christensen, J.H. Beneficial Effect of N-3 Polyunsaturated Fatty Acids on Inflammation and Analgesic Use in Psoriatic Arthritis: A Randomized, Double Blind, Placebo-Controlled Trial. Scand. J. Rheumatol. 2018, 47, 27–36. [Google Scholar] [CrossRef]
- Petrovic, A.; Bueide, I.; Tveit, K.S.; Hallaråker, H.; Bjørndal, B.; Holmes, T.D.; Davies, R.; Brokstad, K.A.; Bergum, B.; Appel, S. Herring Roe Oil in Treatment of Psoriasis—Influence on Immune Cells and Cytokine Network. Front. Immunol. 2023, 14, 1128986. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, Z.; Wu, X.; Chen, C.; Cai, L. The Effects of Fatty Acids on Psoriasis: A Two-Sample Mendelian Randomization Study. J. Cosmet. Dermatol. 2024, 23, 2716–2725. [Google Scholar] [CrossRef]
- Khayef, G.; Young, J.; Burns-Whitmore, B.; Spalding, T. Effects of Fish Oil Supplementation on Inflammatory Acne. Lipids Health Dis. 2012, 11, 165. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Kwon, H.H.; Hong, J.S.; Yoon, J.Y.; Park, M.S.; Jang, M.Y.; Suh, D.H. Effect of Dietary Supplementation with Omega-3 Fatty Acid and Gamma-Linolenic Acid on Acne Vulgaris: A Randomised, Double-Blind, Controlled Trial. Acta Derm. Venereol. 2014, 94, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Aslan, İ.; Özcan, F.; Karaarslan, T.; Kıraç, E.; Aslan, M. Decreased Eicosapentaenoic Acid Levels in Acne Vulgaris Reveals the Presence of a Proinflammatory State. Prostaglandins Other Lipid Mediat. 2017, 128–129, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zainab, Z.; Malik, N.A.; Obaid, S.; Malik, S.; Aftab, K.; Mumtaz, M.; Pervez, A.; Syed, Z. Effectiveness of Oral Omega 3 In Reducing Mucocutaneous Side Effects of Oral Isotretinoin in Patients with Acne Vulgaris. J. Ayub Med. Coll. Abbottabad 2021, 33, 60–63. [Google Scholar]
- Guertler, A.; Neu, K.; Lill, D.; Clanner-Engelshofen, B.; French, L.E.; Reinholz, M. Exploring the Potential of Omega-3 Fatty Acids in Acne Patients: A Prospective Intervention Study. J. Cosmet. Dermatol. 2024, 23, 3295–3304. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, F.; Lai, J.; Jiang, S.; Tan, X.; Chen, L.; Xu, Y.; Xiong, X.; Deng, Y. The Adjuvant Treatment Role of ω-3 Fatty Acids by Regulating Gut Microbiota Positively in the Acne Vulgaris. J. Dermatol. Treat. 2024, 35, 2299107. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Y.; Pu, Y.; Dang, T.; Shi, Q.; Wu, W. Exploring Clinical and Genetic Evidence in Association between Unsaturated Fatty Acids and Acne. Eur. J. Nutr. 2025, 64, 130. [Google Scholar] [CrossRef]
- Weise, C.; Heunemann, C.; Loddenkemper, C.; Herz, U.; van Tol, E.A.F.; Worm, M. Dietary Docosahexaenoic Acid in Combination with Arachidonic Acid Ameliorates Allergen-Induced Dermatitis in Mice. Pediatr. Allergy Immunol. 2011, 22, 497–504. [Google Scholar] [CrossRef]
- Kim, T.-H.; Kim, G.-D.; Jin, Y.-H.; Park, Y.S.; Park, C.-S. Omega-3 Fatty Acid-Derived Mediator, Resolvin E1, Ameliorates 2,4-Dinitrofluorobenzene-Induced Atopic Dermatitis in NC/Nga Mice. Int. Immunopharmacol. 2012, 14, 384–391. [Google Scholar] [CrossRef]
- Han, S.-C.; Koo, D.-H.; Kang, N.-J.; Yoon, W.-J.; Kang, G.-J.; Kang, H.-K.; Yoo, E.-S. Docosahexaenoic Acid Alleviates Atopic Dermatitis by Generating Tregs and IL-10/TGF-β-Modified Macrophages via a TGF-β-Dependent Mechanism. J. Investig. Dermatol. 2015, 135, 1556–1564. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Yasutomo, K.; Watanabe, T. Treatment with DHA/EPA Ameliorates Atopic Dermatitis-like Skin Disease by Blocking LTB4 Production. J. Med. Investig. 2016, 63, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Fujii, M.; Ohyanagi, C.; Kawaguchi, N.; Matsuda, H.; Miyamoto, Y.; Ohya, S.; Nabe, T. Eicosapentaenoic Acid Ethyl Ester Ameliorates Atopic Dermatitis-like Symptoms in Special Diet-Fed Hairless Mice, Partly by Restoring Covalently Bound Ceramides in the Stratum Corneum. Exp. Dermatol. 2018, 27, 837–840. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-C.; Tominaga, M.; Yasukawa, K.; Ohba, M.; Takahashi, N.; Honda, K.; Okuno, T.; Takamori, K.; Yokomizo, T. Dietary Supplementation of Omega-3 Fatty Acid Eicosapentaenoic Acid Does Not Ameliorate Pruritus in Murine Models of Atopic Dermatitis and Psoriasis. J. Dermatol. Sci. 2019, 95, 130–133. [Google Scholar] [CrossRef]
- Sato, A.; Sato, G.; Yabuki, A.; Ogawa, Y.; Nemoto, H.; Ohira, M. Ameliorative Effects of Squid Phospholipids from Todarodes Pacificus on Atopic Dermatitis-like Lesions in NC/Nga Mice. Arch. Dermatol. Res. 2025, 317, 239. [Google Scholar] [CrossRef]
- Montes, R.; Chisaguano, A.M.; Castellote, A.I.; Morales, E.; Sunyer, J.; López-Sabater, M.C. Fatty-Acid Composition of Maternal and Umbilical Cord Plasma and Early Childhood Atopic Eczema in a Spanish Cohort. Eur. J. Clin. Nutr. 2013, 67, 658–663. [Google Scholar] [CrossRef]
- Mihály, J.; Marosvölgyi, T.; Szegedi, A.; Köröskényi, K.; Lucas, R.; Törőcsik, D.; Garcia, A.L.; Decsi, T.; Rühl, R. Increased FADS2-Derived n-6 PUFAs and Reduced n-3 PUFAs in Plasma of Atopic Dermatitis Patients. Ski. Pharmacol. Physiol. 2014, 27, 242–248. [Google Scholar] [CrossRef]
- Gardner, K.G.; Gebretsadik, T.; Hartman, T.J.; Rosa, M.J.; Tylavsky, F.A.; Adgent, M.A.; Moore, P.E.; Kocak, M.; Bush, N.R.; Davis, R.L.; et al. Prenatal Omega-3 and Omega-6 Polyunsaturated Fatty Acids and Childhood Atopic Dermatitis. J. Allergy Clin. Immunol. Pract. 2020, 8, 937–944. [Google Scholar] [CrossRef]
- Mao, R.; Meng, X.; Zhang, T.; Wang, F.; Zhong, Y.; Li, J. Evaluating the Impact of Omega-3 Fatty Acids and Genetic Susceptibility on Atopic Dermatitis in Adults. Mol. Nutr. Food Res. 2025, 69, e70002. [Google Scholar] [CrossRef]
- Bjørneboe, A.; Søyland, E.; Bjørneboe, G.E.; Rajka, G.; Drevon, C.A. Effect of Dietary Supplementation with Eicosapentaenoic Acid in the Treatment of Atopic Dermatitis. Br. J. Dermatol. 1987, 117, 463–469. [Google Scholar] [CrossRef]
- Berth-Jones, J.; Graham-Brown, R.A. Placebo-Controlled Trial of Essential Fatty Acid Supplementation in Atopic Dermatitis. Lancet 1993, 341, 1557–1560. [Google Scholar] [CrossRef]
- Watanabe, T.; Kuroda, Y. The Effect of a Newly Developed Ointment Containing Eicosapentaenoic Acid and Docosahexaenoic Acid in the Treatment of Atopic Dermatitis. J. Med. Investig. 1999, 46, 173–177. [Google Scholar]
- Mayser, P.; Mayer, K.; Mahloudjian, M.; Benzing, S.; Krämer, H.-J.; Schill, W.-B.; Seeger, W.; Grimminger, F. A Double-Blind, Randomized, Placebo-Controlled Trial of n-3 versus n-6 Fatty Acid-Based Lipid Infusion in Atopic Dermatitis. J. Parenter. Enter. Nutr. 2002, 26, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Dunstan, J.A.; Mori, T.A.; Barden, A.; Beilin, L.J.; Taylor, A.L.; Holt, P.G.; Prescott, S.L. Fish Oil Supplementation in Pregnancy Modifies Neonatal Allergen-Specific Immune Responses and Clinical Outcomes in Infants at High Risk of Atopy: A Randomized, Controlled Trial. J. Allergy Clin. Immunol. 2003, 112, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Koch, C.; Dölle, S.; Metzger, M.; Rasche, C.; Jungclas, H.; Rühl, R.; Renz, H.; Worm, M. Docosahexaenoic Acid (DHA) Supplementation in Atopic Eczema: A Randomized, Double-Blind, Controlled Trial. Br. J. Dermatol. 2008, 158, 786–792. [Google Scholar] [CrossRef]
- Furuhjelm, C.; Warstedt, K.; Fagerås, M.; Fälth-Magnusson, K.; Larsson, J.; Fredriksson, M.; Duchén, K. Allergic Disease in Infants up to 2 Years of Age in Relation to Plasma Omega-3 Fatty Acids and Maternal Fish Oil Supplementation in Pregnancy and Lactation. Pediatr. Allergy Immunol. 2011, 22, 505–514. [Google Scholar] [CrossRef]
- Noakes, P.S.; Vlachava, M.; Kremmyda, L.-S.; Diaper, N.D.; Miles, E.A.; Erlewyn-Lajeunesse, M.; Williams, A.P.; Godfrey, K.M.; Calder, P.C. Increased Intake of Oily Fish in Pregnancy: Effects on Neonatal Immune Responses and on Clinical Outcomes in Infants at 6 Mo. Am. J. Clin. Nutr. 2012, 95, 395–404. [Google Scholar] [CrossRef]
- Palmer, D.J.; Sullivan, T.; Gold, M.S.; Prescott, S.L.; Heddle, R.; Gibson, R.A.; Makrides, M. Effect of N-3 Long Chain Polyunsaturated Fatty Acid Supplementation in Pregnancy on Infants’ Allergies in First Year of Life: Randomised Controlled Trial. BMJ 2012, 344, e184. [Google Scholar] [CrossRef]
- Wu, S.-H.; Chen, X.-Q.; Liu, B.; Wu, H.-J.; Dong, L. Efficacy and Safety of 15(R/S)-Methyl-Lipoxin A(4) in Topical Treatment of Infantile Eczema. Br. J. Dermatol. 2013, 168, 172–178. [Google Scholar] [CrossRef]
- Komulainen, M.; Saros, L.; Vahlberg, T.; Nermes, M.; Jartti, T.; Laitinen, K. Maternal Fish Oil and/or Probiotics Intervention: Allergic Diseases in Children up to Two Years Old. Pediatr. Allergy Immunol. 2023, 34, e14004. [Google Scholar] [CrossRef]
- Figueroa-Garduño, I.; Escamilla-Núñez, C.; Barraza-Villarreal, A.; Hernández-Cadena, L.; Onofre-Pardo, E.N.; Romieu, I. Docosahexaenoic Acid Effect on Prenatal Exposure to Arsenic and Atopic Dermatitis in Mexican Preschoolers. Biol. Trace Elem. Res. 2023, 201, 3152–3161. [Google Scholar] [CrossRef]
- Niseteo, T.; Hojsak, I.; Ožanić Bulić, S.; Pustišek, N. Effect of Omega-3 Polyunsaturated Fatty Acid Supplementation on Clinical Outcome of Atopic Dermatitis in Children. Nutrients 2024, 16, 2829. [Google Scholar] [CrossRef] [PubMed]
- Harwood, J.L. Polyunsaturated Fatty Acids: Conversion to Lipid Mediators, Roles in Inflammatory Diseases and Dietary Sources. Int. J. Mol. Sci. 2023, 24, 8838. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Hamberg, M.; Samuelsson, B. Lipoxins: Novel Series of Biologically Active Compounds Formed from Arachidonic Acid in Human Leukocytes. Proc. Natl. Acad. Sci. USA 1984, 81, 5335–5339. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Pro-Resolving Lipid Mediators Are Leads for Resolution Physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef]
- Jerab, D.; Blangero, F.; Da Costa, P.C.T.; De Brito Alves, J.L.; Kefi, R.; Jamoussi, H.; Morio, B.; Eljaafari, A. Beneficial Effects of Omega-3 Fatty Acids on Obesity and Related Metabolic and Chronic Inflammatory Diseases. Nutrients 2025, 17, 1253. [Google Scholar] [CrossRef]
- Yokoyama, M.; Origasa, H.; Matsuzaki, M.; Matsuzawa, Y.; Saito, Y.; Ishikawa, Y.; Oikawa, S.; Sasaki, J.; Hishida, H.; Itakura, H.; et al. Effects of Eicosapentaenoic Acid on Major Coronary Events in Hypercholesterolaemic Patients (JELIS): A Randomised Open-Label, Blinded Endpoint Analysis. Lancet 2007, 369, 1090–1098. [Google Scholar] [CrossRef]
- Liang, J.; Chen, L.; Yang, S.; Zhang, H.; Li, L.; Chen, Z.; Zhang, S.; Xian, H.; Tang, Y.; Deng, Y.; et al. A 12-Hour Rapid Titration Method for Cancer Pain: A Randomized, Controlled, Open-Label Study. Ann. Palliat. Med. 2021, 10, 88–96. [Google Scholar] [CrossRef]
- The ASCEND Study Collaborative Group. Effects of N−3 Fatty Acid Supplements in Diabetes Mellitus. N. Engl. J. Med. 2018, 379, 1540–1550. [Google Scholar] [CrossRef]
- Delpino, F.M.; Figueiredo, L.M.; Da Silva, B.G.C.; Da Silva, T.G.; Mintem, G.C.; Bielemann, R.M.; Gigante, D.P. Omega-3 Supplementation and Diabetes: A Systematic Review and Meta-Analysis. Crit. Rev. Food Sci. Nutr. 2022, 62, 4435–4448. [Google Scholar] [CrossRef]
- Colas, R.A.; Shinohara, M.; Dalli, J.; Chiang, N.; Serhan, C.N. Identification and Signature Profiles for Pro-Resolving and Inflammatory Lipid Mediators in Human Tissue. Am. J. Physiol. Cell Physiol. 2014, 307, C39–C54. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Dalli, J.; Levy, B.D. Lipid Mediators in the Resolution of Inflammation. Cold Spring Harb. Perspect. Biol. 2015, 7, a016311. [Google Scholar] [CrossRef] [PubMed]
- Bannenberg, G.; Serhan, C.N. Specialized Pro-Resolving Lipid Mediators in the Inflammatory Response: An Update. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2010, 1801, 1260–1273. [Google Scholar] [CrossRef] [PubMed]
- Ariel, A.; Chiang, N.; Arita, M.; Petasis, N.A.; Serhan, C.N. Aspirin-Triggered Lipoxin A4 and B4 Analogs Block Extracellular Signal-Regulated Kinase-Dependent TNF-Alpha Secretion from Human T Cells. J. Immunol. 2003, 170, 6266–6272. [Google Scholar] [CrossRef] [PubMed]
- Godson, C.; Mitchell, S.; Harvey, K.; Petasis, N.A.; Hogg, N.; Brady, H.R. Cutting Edge: Lipoxins Rapidly Stimulate Nonphlogistic Phagocytosis of Apoptotic Neutrophils by Monocyte-Derived Macrophages. J. Immunol. 2000, 164, 1663–1667. [Google Scholar] [CrossRef]
- Roach, K.M.; Feghali-Bostwick, C.A.; Amrani, Y.; Bradding, P. Lipoxin A4 Attenuates Constitutive and TGF-Β1–Dependent Profibrotic Activity in Human Lung Myofibroblasts. J. Immunol. 2015, 195, 2852–2860. [Google Scholar] [CrossRef]
- Wu, L.; Li, H.-H.; Wu, Q.; Miao, S.; Liu, Z.-J.; Wu, P.; Ye, D.-Y. Lipoxin A4 Activates Nrf2 Pathway and Ameliorates Cell Damage in Cultured Cortical Astrocytes Exposed to Oxygen-Glucose Deprivation/Reperfusion Insults. J. Mol. Neurosci. 2015, 56, 848–857. [Google Scholar] [CrossRef]
- Levy, B.D. Myocardial 15-Epi-Lipoxin A4 Generation Provides a New Mechanism for the Immunomodulatory Effects of Statins and Thiazolidinediones. Circulation 2006, 114, 873–875. [Google Scholar] [CrossRef]
- Clària, J.; Serhan, C.N. Aspirin Triggers Previously Undescribed Bioactive Eicosanoids by Human Endothelial Cell-Leukocyte Interactions. Proc. Natl. Acad. Sci. USA 1995, 92, 9475–9479. [Google Scholar] [CrossRef]
- József, L.; Zouki, C.; Petasis, N.A.; Serhan, C.N.; Filep, J.G. Lipoxin A4 and Aspirin-Triggered 15-Epi-Lipoxin A4 Inhibit Peroxynitrite Formation, NF-κB and AP-1 Activation, and IL-8 Gene Expression in Human Leukocytes. Proc. Natl. Acad. Sci. USA 2002, 99, 13266–13271. [Google Scholar] [CrossRef]
- Serhan, C.N.; Chiang, N.; Dalli, J. The Resolution Code of Acute Inflammation: Novel pro-Resolving Lipid Mediators in Resolution. Semin. Immunol. 2015, 27, 200–215. [Google Scholar] [CrossRef]
- Pirault, J.; Bäck, M. Lipoxin and Resolvin Receptors Transducing the Resolution of Inflammation in Cardiovascular Disease. Front. Pharmacol. 2018, 9, 1273. [Google Scholar] [CrossRef]
- Arita, M.; Yoshida, M.; Hong, S.; Tjonahen, E.; Glickman, J.N.; Petasis, N.A.; Blumberg, R.S.; Serhan, C.N. Resolvin E1, an Endogenous Lipid Mediator Derived from Omega-3 Eicosapentaenoic Acid, Protects against 2,4,6-Trinitrobenzene Sulfonic Acid-Induced Colitis. Proc. Natl. Acad. Sci. USA 2005, 102, 7671–7676. [Google Scholar] [CrossRef] [PubMed]
- Libreros, S.; Shay, A.E.; Nshimiyimana, R.; Fichtner, D.; Martin, M.J.; Wourms, N.; Serhan, C.N. A New E-Series Resolvin: RvE4 Stereochemistry and Function in Efferocytosis of Inflammation-Resolution. Front. Immunol. 2021, 11, 631319. [Google Scholar] [CrossRef] [PubMed]
- Dalli, J.; Winkler, J.W.; Colas, R.A.; Arnardottir, H.; Cheng, C.-Y.C.; Chiang, N.; Petasis, N.A.; Serhan, C.N. Resolvin D3 and Aspirin-Triggered Resolvin D3 Are Potent Immunoresolvents. Chem. Biol. 2013, 20, 188–201. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Hong, S.; Gronert, K.; Colgan, S.P.; Devchand, P.R.; Mirick, G.; Moussignac, R.-L. Resolvins. J. Exp. Med. 2002, 196, 1025–1037. [Google Scholar] [CrossRef]
- Krishnamoorthy, S.; Recchiuti, A.; Chiang, N.; Fredman, G.; Serhan, C.N. Resolvin D1 Receptor Stereoselectivity and Regulation of Inflammation and Proresolving MicroRNAs. Am. J. Pathol. 2012, 180, 2018–2027. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Marcheselli, V.L.; Serhan, C.N.; Bazan, N.G. Neuroprotectin D1: A Docosahexaenoic Acid-Derived Docosatriene Protects Human Retinal Pigment Epithelial Cells from Oxidative Stress. Proc. Natl. Acad. Sci. USA 2004, 101, 8491–8496. [Google Scholar] [CrossRef]
- Morita, M.; Kuba, K.; Ichikawa, A.; Nakayama, M.; Katahira, J.; Iwamoto, R.; Watanebe, T.; Sakabe, S.; Daidoji, T.; Nakamura, S.; et al. The Lipid Mediator Protectin D1 Inhibits Influenza Virus Replication and Improves Severe Influenza. Cell 2013, 153, 112–125. [Google Scholar] [CrossRef]
- Liu, W.-C.; Yang, Y.-H.; Wang, Y.-C.; Chang, W.-M.; Wang, C.-W. Maresin: Macrophage Mediator for Resolving Inflammation and Bridging Tissue Regeneration—A System-Based Preclinical Systematic Review. Int. J. Mol. Sci. 2023, 24, 11012. [Google Scholar] [CrossRef]
- Serhan, C.N.; Yang, R.; Martinod, K.; Kasuga, K.; Pillai, P.S.; Porter, T.F.; Oh, S.F.; Spite, M. Maresins: Novel Macrophage Mediators with Potent Antiinflammatory and Proresolving Actions. J. Exp. Med. 2009, 206, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Dalli, J.; Karamnov, S.; Choi, A.; Park, C.; Xu, Z.; Ji, R.; Zhu, M.; Petasis, N.A. Macrophage Proresolving Mediator Maresin 1 Stimulates Tissue Regeneration and Controls Pain. FASEB J. 2012, 26, 1755–1765. [Google Scholar] [CrossRef] [PubMed]
- Berdyshev, E. Skin Lipid Barrier: Structure, Function and Metabolism. Allergy Asthma Immunol. Res. 2024, 16, 445–461. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G.; Yada, Y.; Higuchi, K.; Okuda, M.; Ohashi, Y.; Kawamata, A. Pseudo-Acylceramide with Linoleic Acid Produces Selective Recovery of Diminished Cutaneous Barrier Function in Essential Fatty Acid-Deficient Rats and Has an Inhibitory Effect on Epidermal Hyperplasia. J. Clin. Investig. 1994, 94, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Harauma, A.; Enomoto, Y.; Endo, S.; Hariya, H.; Moriguchi, T. Omega-3 Fatty Acids Mitigate Skin Damage Caused by Ultraviolet-B Radiation. Prostaglandins Leukot. Essent. Fat. Acids 2024, 203, 102641. [Google Scholar] [CrossRef]
- Sawada, Y.; Saito-Sasaki, N.; Nakamura, M. Omega 3 Fatty Acid and Skin Diseases. Front. Immunol. 2020, 11, 623052. [Google Scholar] [CrossRef]
- Töröcsik, D.; Weise, C.; Gericke, J.; Szegedi, A.; Lucas, R.; Mihaly, J.; Worm, M.; Rühl, R. Transcriptomic and Lipidomic Profiling of Eicosanoid/Docosanoid Signalling in Affected and Non-Affected Skin of Human Atopic Dermatitis Patients. Exp. Dermatol. 2019, 28, 177–189. [Google Scholar] [CrossRef]
- Liu, M.; Diaz-Torres, S.; Mitchell, B.L.; Toledo-Flores, D.; Gharhakhani, P.; Simpson, M.A.; Zhang, H.; Ong, J.-S.; Li, J.; Rentería, M.E. The Role of Lipid Metabolism in Acne Risk: Integrating Blood Metabolite and Genetic Insights. Ski. Health Dis. 2025, 5, 124–129. [Google Scholar] [CrossRef]
- McCusker, M.M.; Grant-Kels, J.M. Healing Fats of the Skin: The Structural and Immunologic Roles of the ω-6 and ω-3 Fatty Acids. Clin. Dermatol. 2010, 28, 440–451. [Google Scholar] [CrossRef]
- De Spirt, S.; Stahl, W.; Tronnier, H.; Sies, H.; Bejot, M.; Maurette, J.-M.; Heinrich, U. Intervention with Flaxseed and Borage Oil Supplements Modulates Skin Condition in Women. Br. J. Nutr. 2008, 101, 440–445. [Google Scholar] [CrossRef]
- Nowowiejska, J.; Baran, A.; Flisiak, I. Aberrations in Lipid Expression and Metabolism in Psoriasis. Int. J. Mol. Sci. 2021, 22, 6561. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C.; Waitzberg, D.L.; Klek, S.; Martindale, R.G. Lipids in Parenteral Nutrition: Biological Aspects. J. Parenter. Enter. Nutr. 2020, 44, S21–S27. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Eicosapentaenoic and Docosahexaenoic Acid Derived Specialised Pro-Resolving Mediators: Concentrations in Humans and the Effects of Age, Sex, Disease and Increased Omega-3 Fatty Acid Intake. Biochimie 2020, 178, 105–123. [Google Scholar] [CrossRef] [PubMed]
- Blauvelt, A.; Chiricozzi, A. The Immunologic Role of IL-17 in Psoriasis and Psoriatic Arthritis Pathogenesis. Clin. Rev. Allergy Immunol. 2018, 55, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Tawada, C.; Kanoh, H.; Nakamura, M.; Mizutani, Y.; Fujisawa, T.; Banno, Y.; Seishima, M. Interferon-γ Decreases Ceramides with Long-Chain Fatty Acids: Possible Involvement in Atopic Dermatitis and Psoriasis. J. Investig. Dermatol. 2014, 134, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; Chen, J.; Swindell, W.R.; Tsoi, L.C.; Xing, X.; Ma, F.; Uppala, R.; Sarkar, M.K.; Plazyo, O.; Billi, A.C.; et al. Phospholipase A2 Enzymes Represent a Shared Pathogenic Pathway in Psoriasis and Pityriasis Rubra Pilaris. J. Clin. Investig. 2021, 6, e151911. [Google Scholar] [CrossRef]
- Myśliwiec, H.; Baran, A.; Harasim-Symbor, E.; Myśliwiec, P.; Milewska, A.J.; Chabowski, A.; Flisiak, I. Serum Fatty Acid Profile in Psoriasis and Its Comorbidity. Arch. Dermatol. Res. 2017, 309, 371–380. [Google Scholar] [CrossRef]
- Marchlewicz, M.; Polakowska, Z.; Maciejewska-Markiewicz, D.; Stachowska, E.; Jakubiak, N.; Kiedrowicz, M.; Rak-Załuska, A.; Duchnik, M.; Wajs-Syrenicz, A.; Duchnik, E. Fatty Acid Profile of Erythrocyte Membranes in Patients with Psoriasis. Nutrients 2024, 16, 1799. [Google Scholar] [CrossRef]
- Kozłowska, D.; Harasim-Symbor, E.; Myśliwiec, H.; Milewska, A.; Chabowski, A.; Flisiak, I. Lipid Profile Disturbances May Predispose Psoriatic Patients to Liver Dysfunction. Adv. Dermatol. Allergol. Postępy Dermatol. I Alergol. 2021, 38, 310–318. [Google Scholar] [CrossRef]
- Serhan, C.N.; Bäck, M.; Chiurchiù, V.; Hersberger, M.; Mittendorfer, B.; Calder, P.C.; Waitzberg, D.L.; Stoppe, C.; Klek, S.; Martindale, R.G.; et al. Expert Consensus Report on Lipid Mediators: Role in Resolution of Inflammation and Muscle Preservation. FASEB J. 2024, 38, e23699. [Google Scholar] [CrossRef]
- Yang, S.-J.; Chi, C.-C. Effects of Fish Oil Supplement on Psoriasis: A Meta-Analysis of Randomized Controlled Trials. BMC Complement. Altern. Med. 2019, 19, 354. [Google Scholar] [CrossRef] [PubMed]
- Riveira-Munoz, E.; He, S.-M.; Escaramís, G.; Stuart, P.E.; Hüffmeier, U.; Lee, C.; Kirby, B.; Oka, A.; Giardina, E.; Liao, W.; et al. Meta-Analysis Confirms the LCE3C_LCE3B Deletion as a Risk Factor for Psoriasis in Several Ethnic Groups and Finds Interaction with HLA-Cw6. J. Investig. Dermatol. 2011, 131, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Dou, Y.; Lin, Y.; Chu, G.; Wang, J.; Ma, L. HMGB1 Regulates Th17 Cell Differentiation and Function in Patients with Psoriasis. Immun. Inflamm. Dis. 2024, 12, e1205. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.S.L.E.; Bloise, G.; Moltrasio, C.; Coelho, A.; Agrelli, A.; Moura, R.; Tricarico, P.M.; Jamain, S.; Marzano, A.V.; Crovella, S.; et al. Transcriptome Meta-Analysis Confirms the Hidradenitis Suppurativa Pathogenic Triad: Upregulated Inflammation, Altered Epithelial Organization, and Dysregulated Metabolic Signaling. Biomolecules 2022, 12, 1371. [Google Scholar] [CrossRef] [PubMed]
- Sabat, R.; Alavi, A.; Wolk, K.; Wortsman, X.; McGrath, B.; Garg, A.; Szepietowski, J.C. Hidradenitis Suppurativa. Lancet 2025, 405, 420–438. [Google Scholar] [CrossRef]
- Krajewski, P.K.; Matusiak, Ł.; Von Stebut, E.; Schultheis, M.; Kirschner, U.; Nikolakis, G.; Szepietowski, J.C. Quality-of-Life Impairment among Patients with Hidradenitis Suppurativa: A Cross-Sectional Study of 1795 Patients. Life 2021, 11, 34. [Google Scholar] [CrossRef]
- Gold, D.A.; Reeder, V.J.; Mahan, M.G.; Hamzavi, I.H. The Prevalence of Metabolic Syndrome in Patients with Hidradenitis Suppurativa. J. Am. Acad. Dermatol. 2014, 70, 699–703. [Google Scholar] [CrossRef]
- Dany, M.; Elston, D. Gene Expression of Sphingolipid Metabolism Pathways Is Altered in Hidradenitis Suppurativa. J. Am. Acad. Dermatol. 2017, 77, 268–273.e6. [Google Scholar] [CrossRef]
- Choi, E.; Mir, S.A.; Ji, S.; Ooi, X.T.; Chua, E.W.L.; Wei, Y.Y.; Wenk, M.R.; Bendt, A.K.; Chandran, N.S. Understanding the Systemic Burden of Disease in Hidradenitis Suppurativa from Plasma Lipidomic Analysis. J. Dermatol. Sci. 2022, 107, 133–141. [Google Scholar] [CrossRef]
- Penno, C.A.; Jäger, P.; Laguerre, C.; Hasler, F.; Hofmann, A.; Gass, S.K.; Wettstein-Ling, B.; Schaefer, D.J.; Avrameas, A.; Raulf, F.; et al. Lipidomics Profiling of Hidradenitis Suppurativa Skin Lesions Reveals Lipoxygenase Pathway Dysregulation and Accumulation of Proinflammatory Leukotriene B4. J. Investig. Dermatol. 2020, 140, 2421–2432.e10. [Google Scholar] [CrossRef]
- Geng, R.; Sibbald, R.G. Acne Vulgaris: Clinical Aspects and Treatments. Adv. Ski. Wound Care 2024, 37, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Sutaria, A.H.; Masood, S.; Saleh, H.M.; Schlessinger, J. Acne Vulgaris. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2025. [Google Scholar]
- Smith, R.N.; Mann, N.J.; Braue, A.; Mäkeläinen, H.; Varigos, G.A. A Low-Glycemic-Load Diet Improves Symptoms in Acne Vulgaris Patients: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2007, 86, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Vasam, M.; Korutla, S.; Bohara, R.A. Acne Vulgaris: A Review of the Pathophysiology, Treatment, and Recent Nanotechnology Based Advances. Biochem. Biophys. Rep. 2023, 36, 101578. [Google Scholar] [CrossRef] [PubMed]
- Tanghetti, E.A. The Role of Inflammation in the Pathology of Acne. J. Clin. Aesthet. Dermatol. 2013, 6, 27–35. [Google Scholar] [PubMed]
- Jeremy, A.H.T.; Holland, D.B.; Roberts, S.G.; Thomson, K.F.; Cunliffe, W.J. Inflammatory Events Are Involved in Acne Lesion Initiation. J. Investig. Dermatol. 2003, 121, 20–27. [Google Scholar] [CrossRef]
- Leyden, J.J.; McGinley, K.J.; Mills, O.H.; Kligman, A.M. Propionibacterium Levels In Patients With And Without Acne Vulgaris. J. Investig. Dermatol. 1975, 65, 382–384. [Google Scholar] [CrossRef]
- Sobhan, M.; Seif Rabiei, M.A.; Amerifar, M. Correlation Between Lipid Profile and Acne Vulgaris. Clin. Cosmet. Investig. Dermatol. 2020, 13, 67–71. [Google Scholar] [CrossRef]
- Jiang, H.; Li, C.Y.; Zhou, L.; Lu, B.; Lin, Y.; Huang, X.; Wei, B.; Wang, Q.; Wang, L.; Lu, J. Acne Patients Frequently Associated with Abnormal Plasma Lipid Profile. J. Dermatol. 2015, 42, 296–299. [Google Scholar] [CrossRef]
- Gokdemir, G.S.; Alp, S.; Nas, C.; Ecevit, H. Evaluation of the Relationship between Lipid Profile and Inflammatory Parameters in Patients with Acne Vulgaris. J. Med. Dent. Investig. 2024, 5, e240384. [Google Scholar] [CrossRef]
- Kelhälä, H.-L.; Palatsi, R.; Fyhrquist, N.; Lehtimäki, S.; Väyrynen, J.P.; Kallioinen, M.; Kubin, M.E.; Greco, D.; Tasanen, K.; Alenius, H.; et al. IL-17/Th17 Pathway Is Activated in Acne Lesions. PLoS ONE 2014, 9, e105238. [Google Scholar] [CrossRef]
- Kistowska, M.; Meier, B.; Proust, T.; Feldmeyer, L.; Cozzio, A.; Kuendig, T.; Contassot, E.; French, L.E. Propionibacterium Acnes Promotes Th17 and Th17/Th1 Responses in Acne Patients. J. Investig. Dermatol. 2015, 135, 110–118. [Google Scholar] [CrossRef]
- Fasseeh, A.N.; Elezbawy, B.; Korra, N.; Tannira, M.; Dalle, H.; Aderian, S.; Abaza, S.; Kaló, Z. Burden of Atopic Dermatitis in Adults and Adolescents: A Systematic Literature Review. Dermatol. Ther. 2022, 12, 2653–2668. [Google Scholar] [CrossRef]
- Mirrahimi, B.; Moazemi, M.; Eslami, N.; Jamshidi, E.; Mir, M.; Mohebbi, R.; Esmaily, H. Evaluating the Effect of Eicosapentaenoic Acid in Children With Atopic Dermatitis: A Randomized Triple-Blind Clinical Trial. J. Pediatr. Pharmacol. Ther. 2023, 28, 29–35. [Google Scholar] [CrossRef] [PubMed]
- DiNicolantonio, J.J.; O’Keefe, J. The Importance of Maintaining a Low Omega-6/Omega-3 Ratio for Reducing the Risk of Autoimmune Diseases, Asthma, and Allergies. Mo. Med. 2021, 118, 453–459. [Google Scholar] [PubMed]
- Fania, L.; Moretta, G.; Antonelli, F.; Scala, E.; Abeni, D.; Albanesi, C.; Madonna, S. Multiple Roles for Cytokines in Atopic Dermatitis: From Pathogenic Mediators to Endotype-Specific Biomarkers to Therapeutic Targets. Int. J. Mol. Sci. 2022, 23, 2684. [Google Scholar] [CrossRef] [PubMed]
- Livshits, G.; Kalinkovich, A. Resolution of Chronic Inflammation, Restoration of Epigenetic Disturbances and Correction of Dysbiosis as an Adjunctive Approach to the Treatment of Atopic Dermatitis. Cells 2024, 13, 1899. [Google Scholar] [CrossRef]
- Jia, Y.; Huang, Y.; Wang, H.; Jiang, H. Effect of Prenatal Omega-3 Polyunsaturated Fatty Acid Supplementation on Childhood Eczema: A Systematic Review and Meta-Analysis. Int. Arch. Allergy Immunol. 2023, 184, 21–32. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.