
Synergistic Antioxidant and Anti-Inflammatory Activities of Kale Juice Fermented with Limosilactobacills reuteri EFEL6901 or Limosilactobacills fermentum EFEL6800


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Characteristics of Fermented Kale Juices
2.3. DPPH Radical Scavenging Activity Assay
2.4. ABTS Radical Scavenging Activity Assay
2.5. Quercetin and Kaempferol Analysis
2.6. 1H-NMR Analysis
2.7. Cytotoxicity of Fermented Kale Juice
2.8. Measurement of Nitric Oxide (NO) Production
2.9. mRNA Expression Level of Nitric Oxide Synthase (iNOS), Cyclooxygenase-2 (COX-2), and Cytokine
2.10. Enzyme and Related Genes
2.11. Statistical Analysis
3. Results
3.1. Quality Characteristics of Fermented Kale Juice
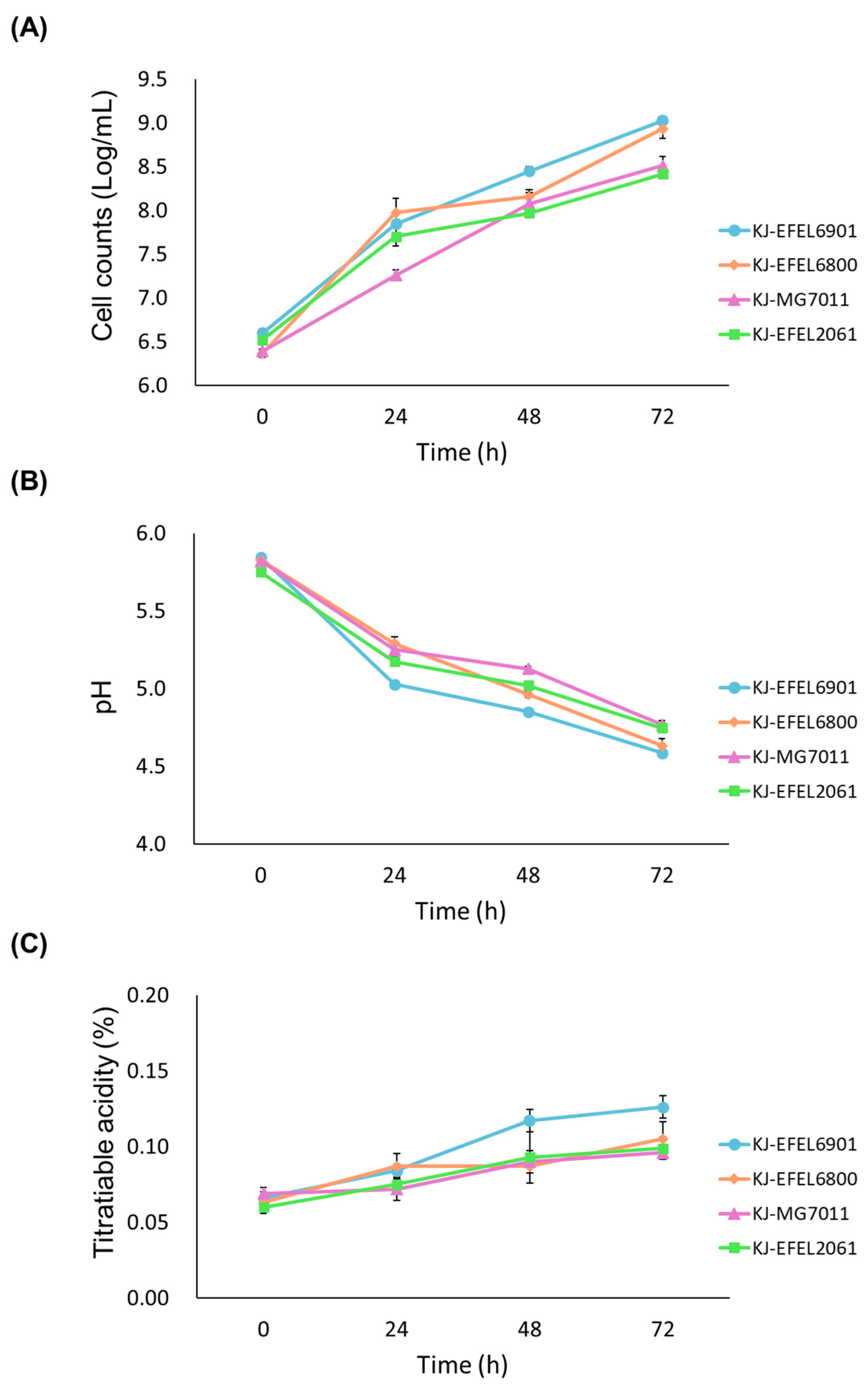
3.1.1. Microbial Growth in Kale Juice
3.1.2. Chromaticity Analysis
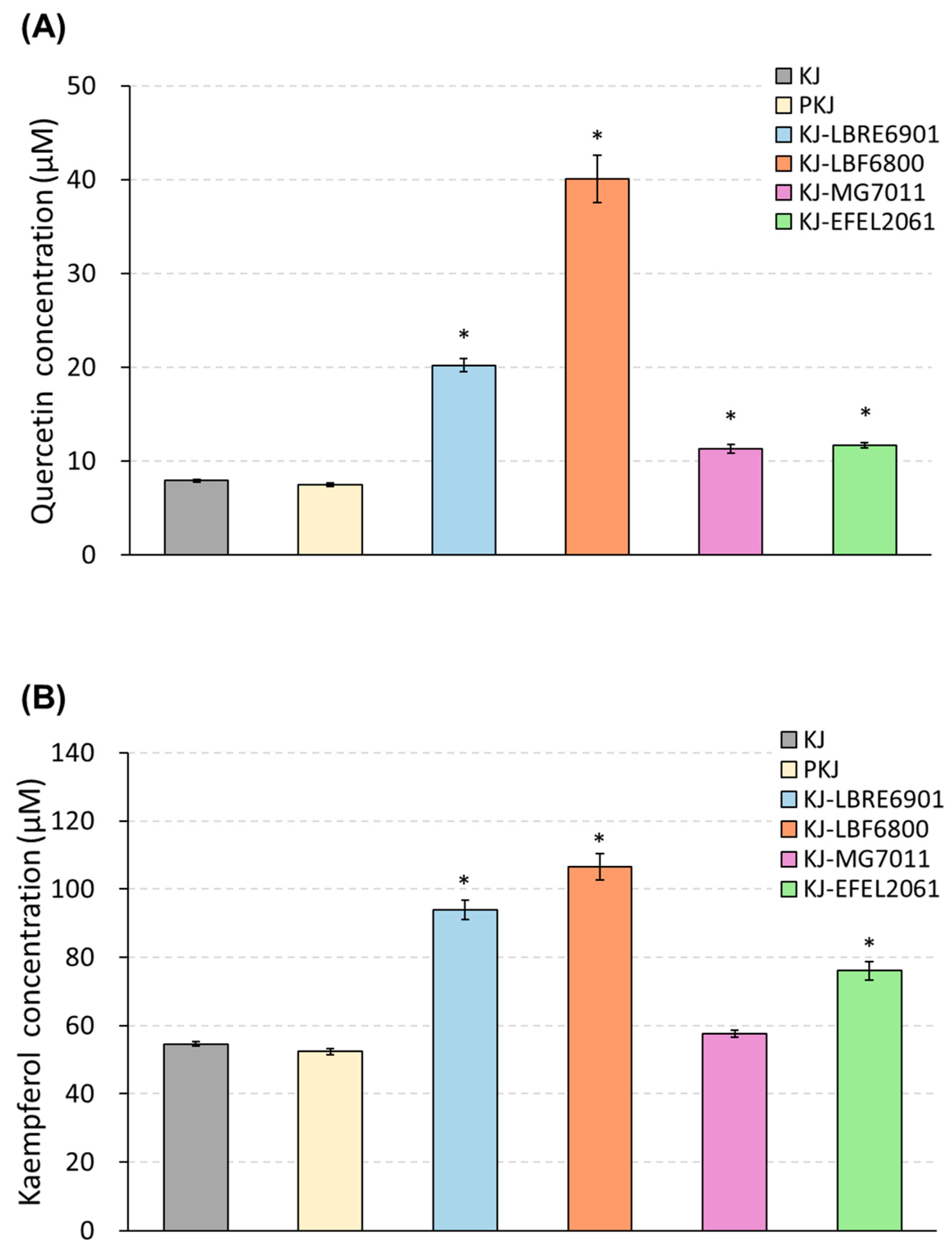
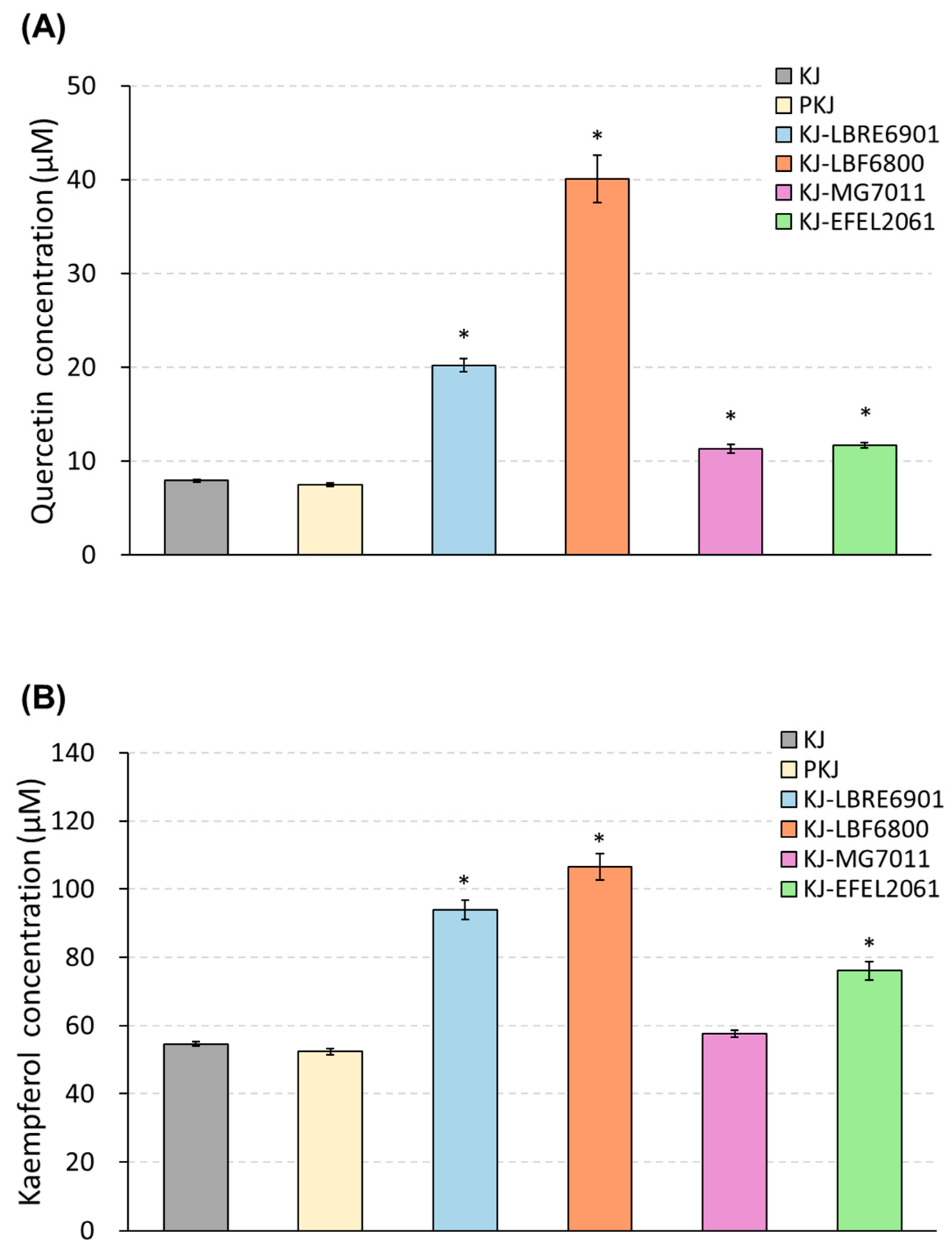
3.1.3. Quantification of Quercetin and Kaempferol
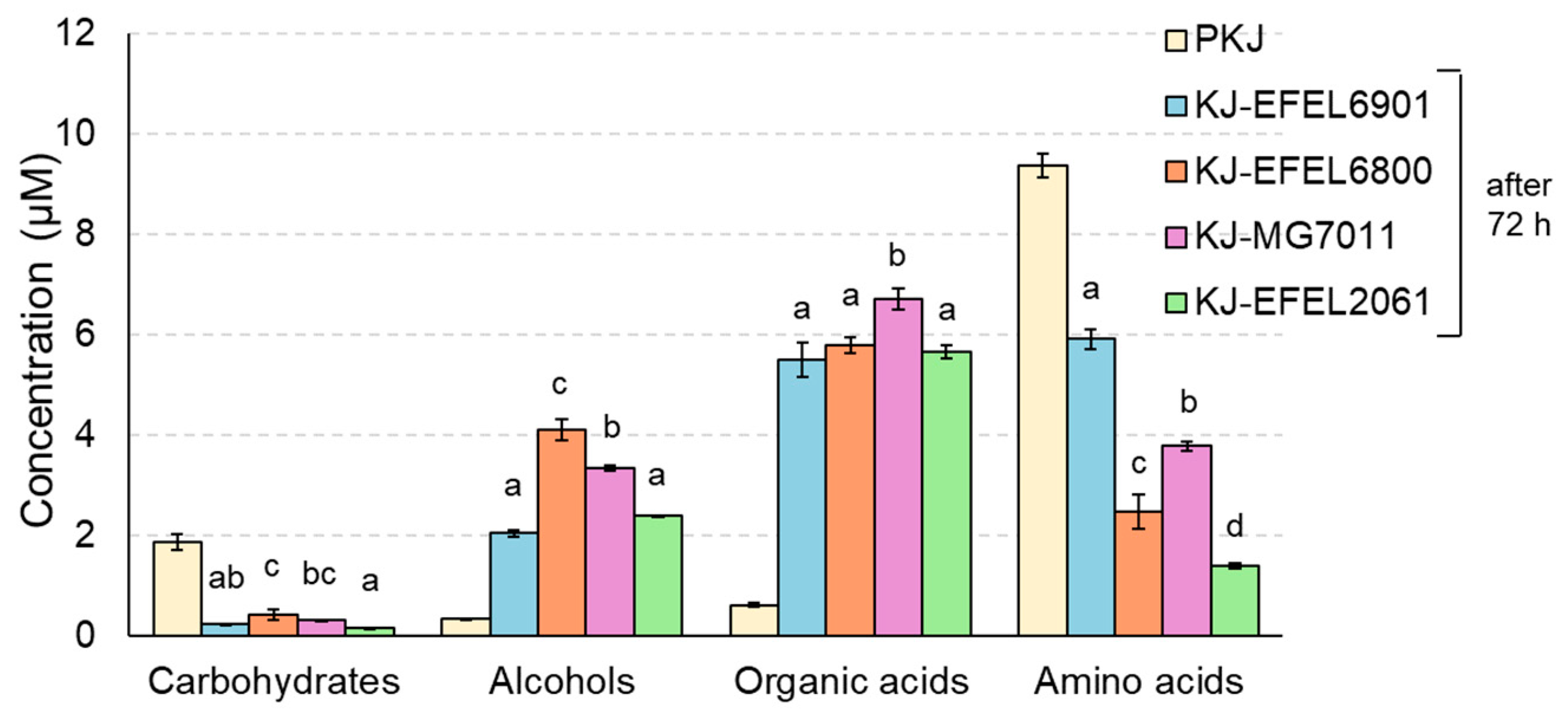
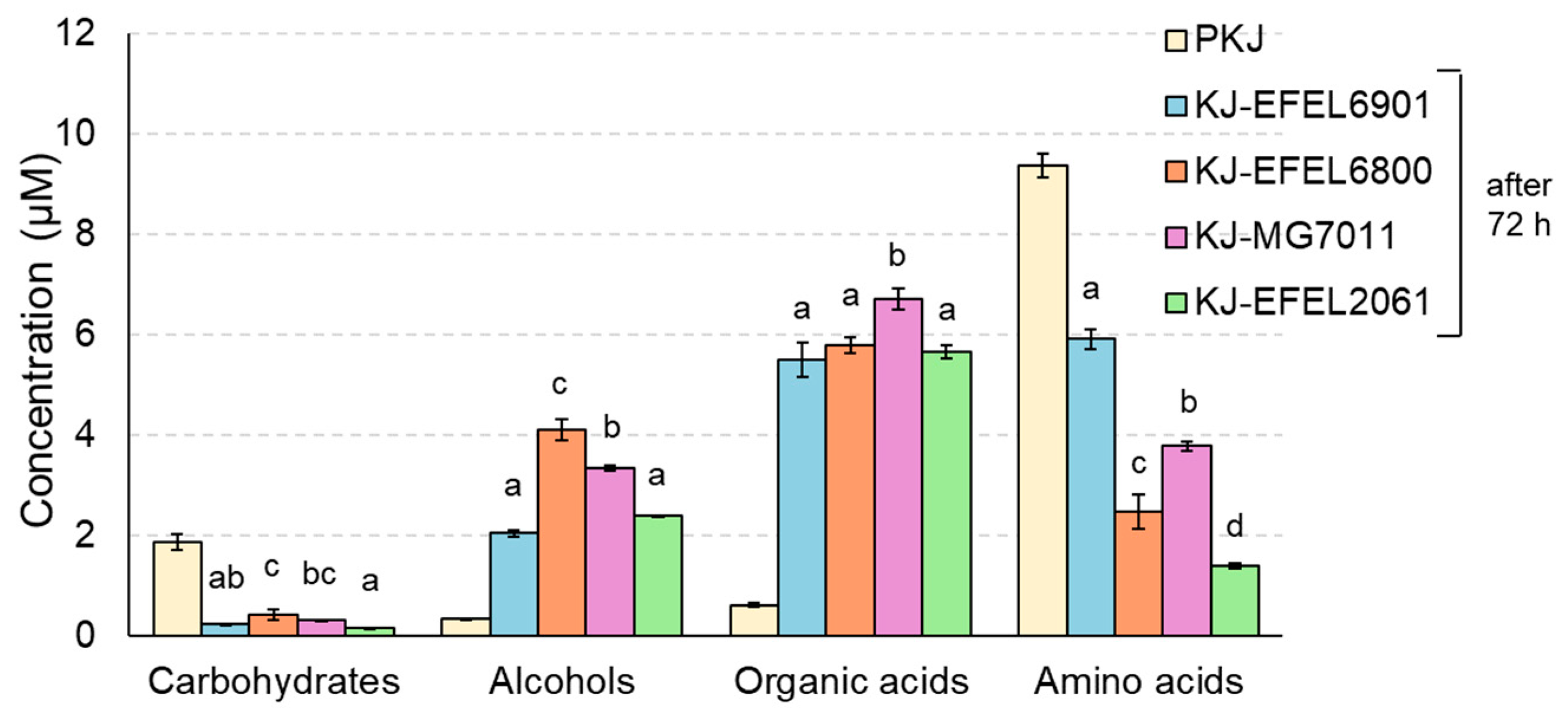
3.1.4. H-NMR Analysis
3.1.5. Enzymes and Genes
3.2. Health-Related Characteristics of Fermented Kale Juice
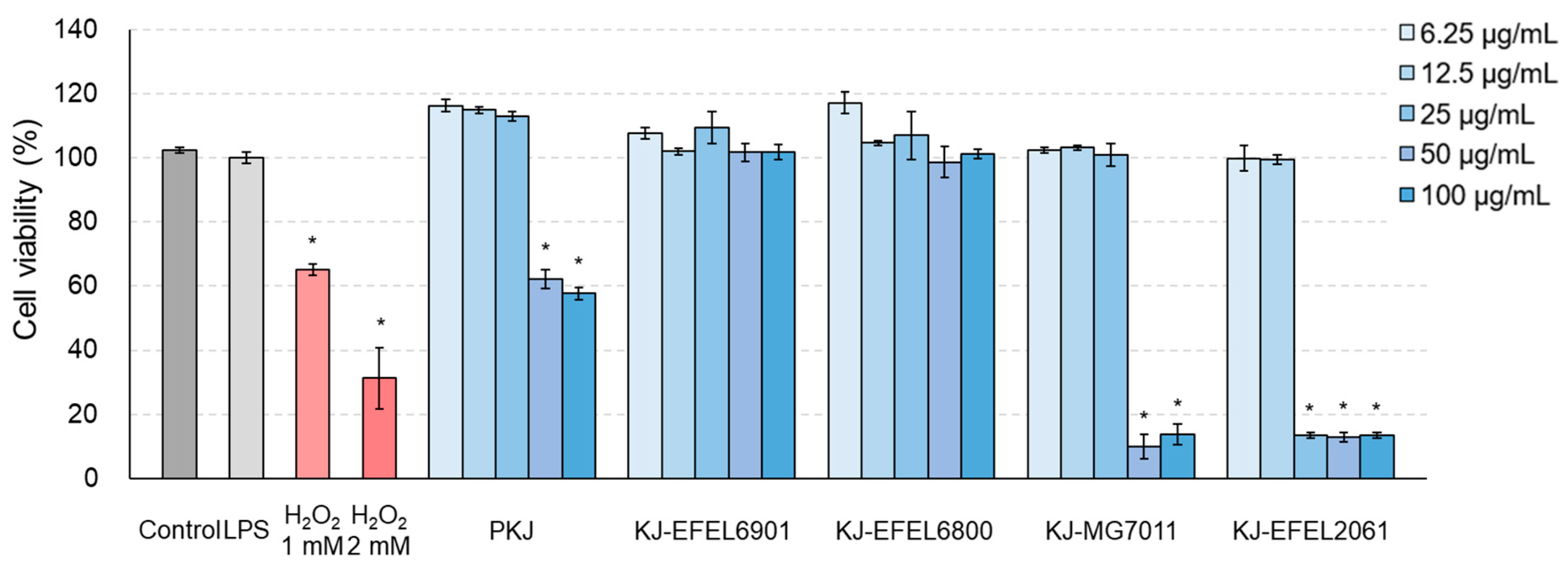
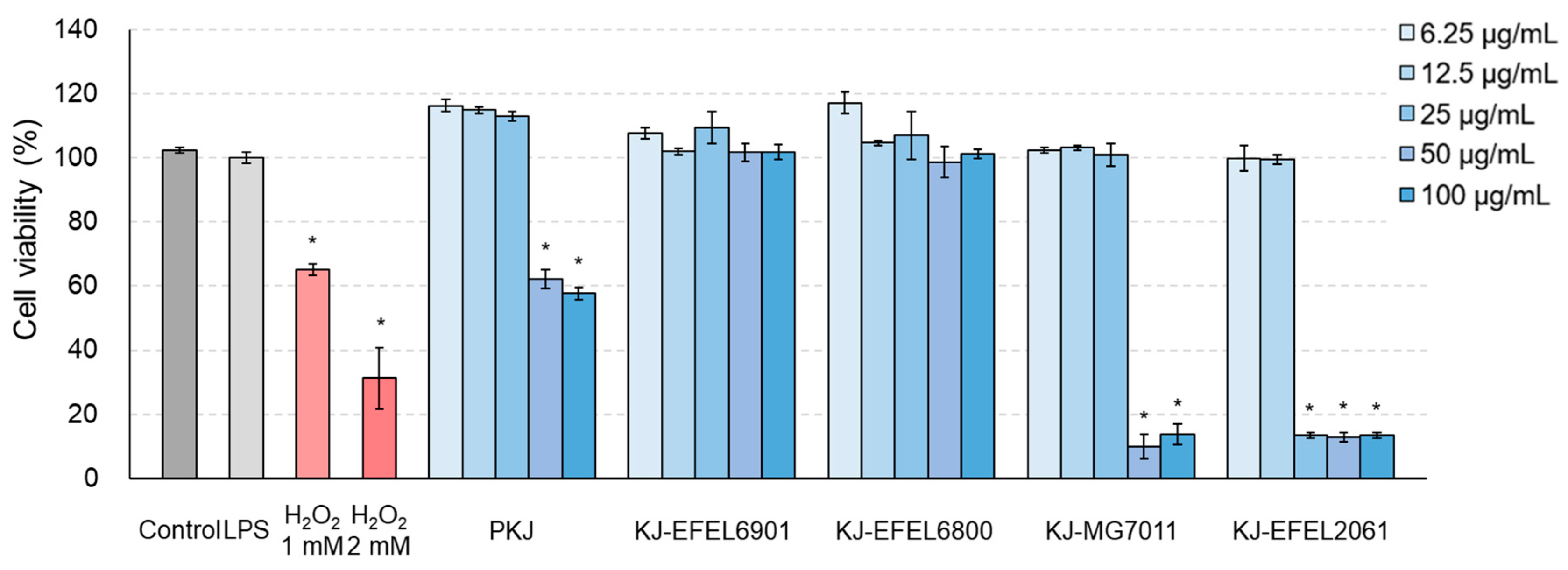
3.2.1. Cytotoxicity
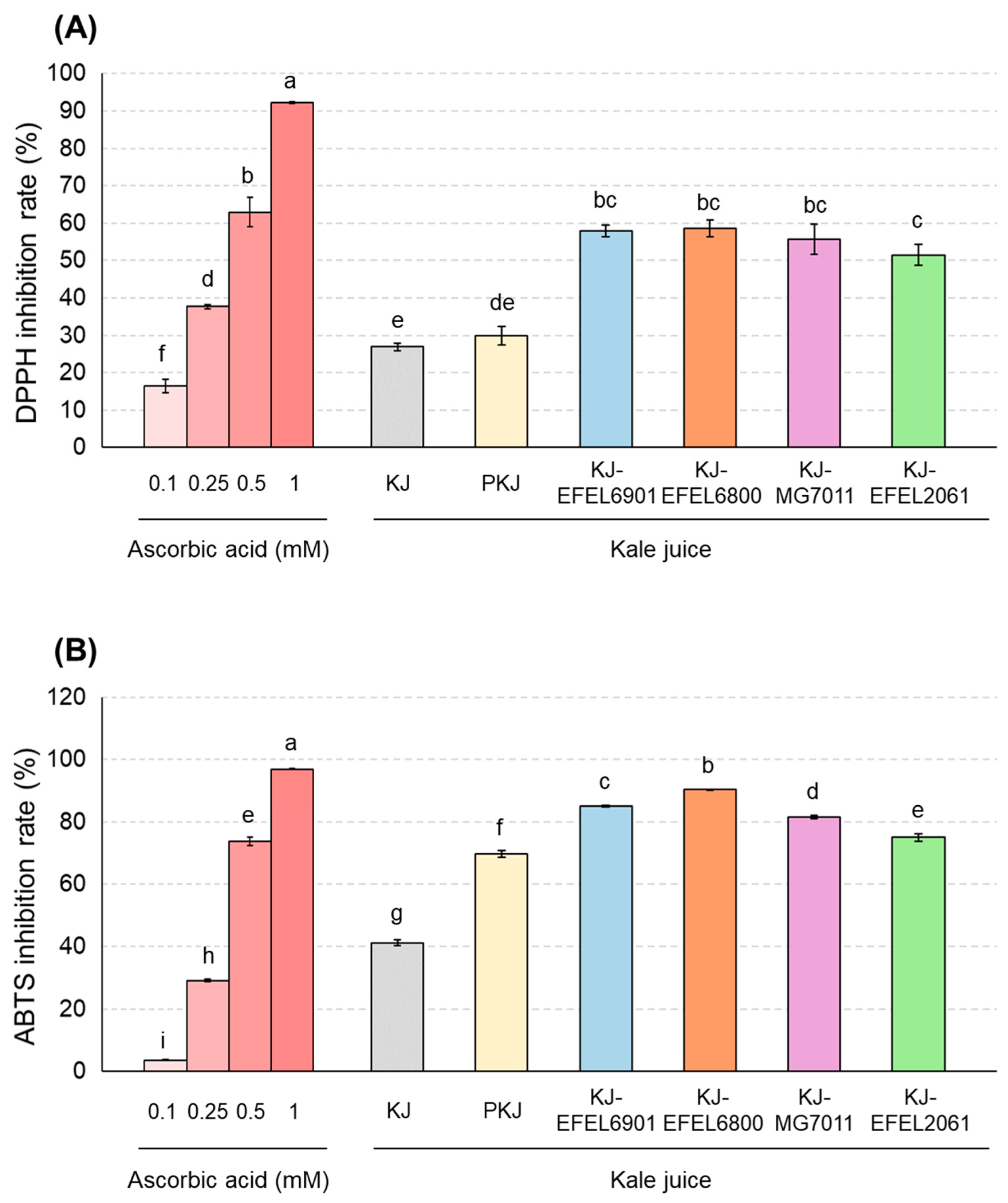
3.2.2. Antioxidant Activities
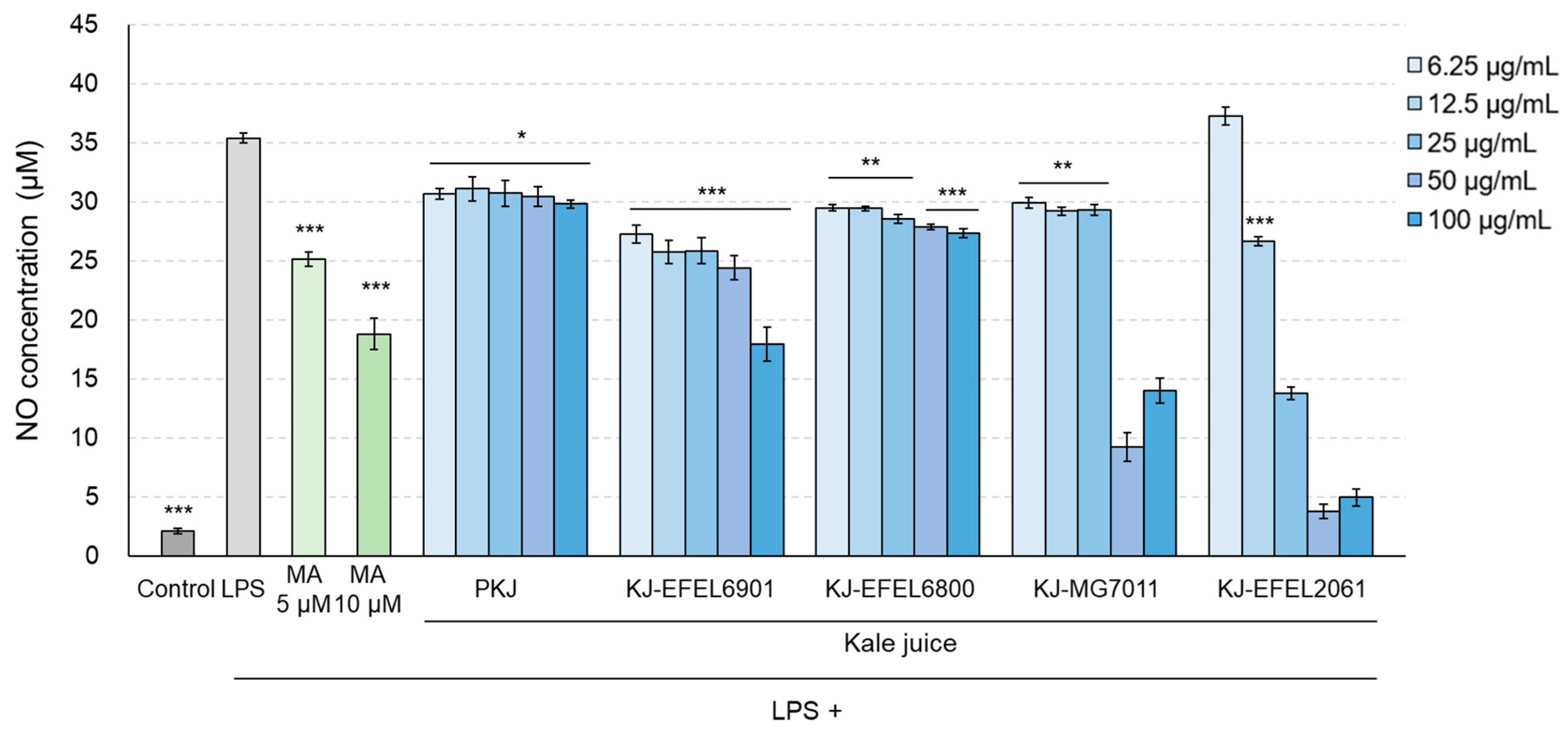
3.2.3. NO Production
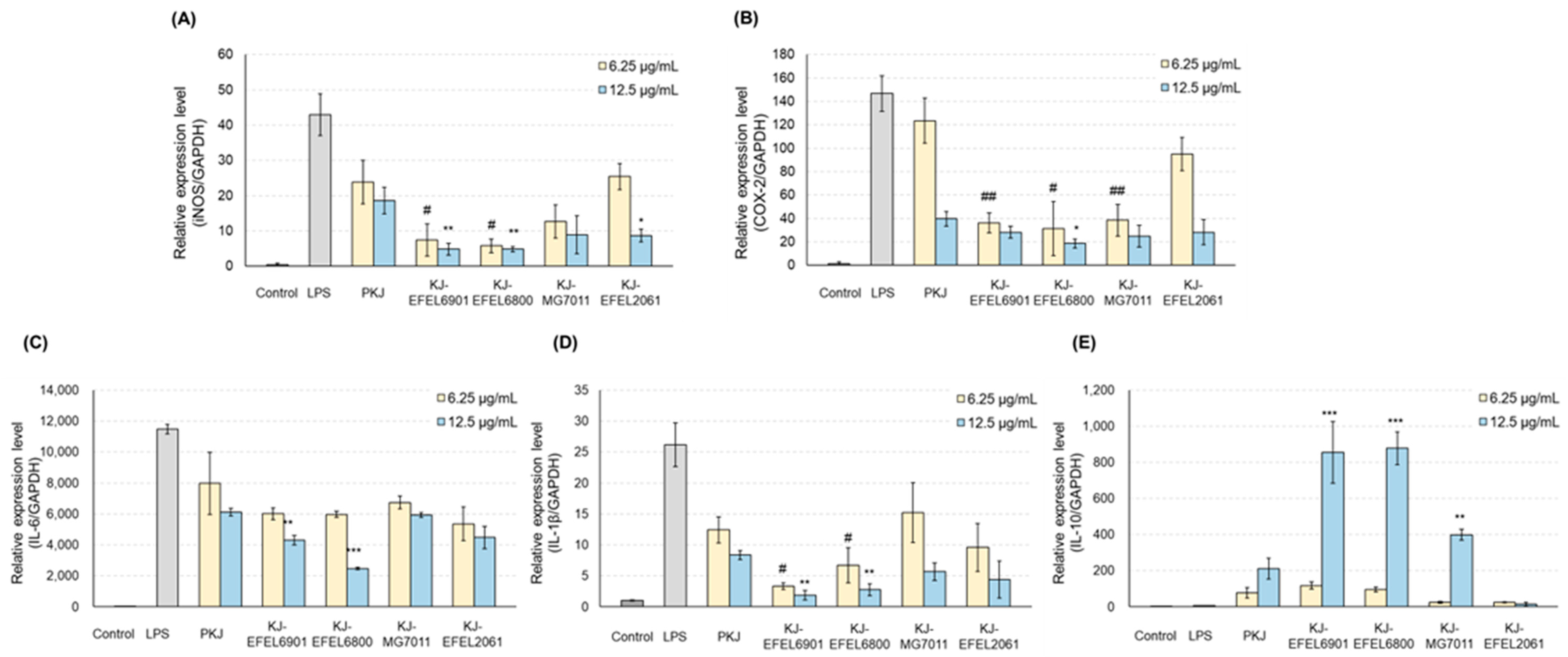
3.2.4. mRNA Levels of iNOS and COX-2
3.2.5. mRNA Levels of Cytokines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marrero, S.C.; Martínez-Rodríguez, A.; Pérez, S.E.M.; Moya, S.P. New Trends and Applications in Fermented Beverages. In Fermented Beverages; Elsevier: Amsterdam, The Netherlands, 2019; pp. 31–66. [Google Scholar] [CrossRef]
- Septembre-Malaterre, A.; Remize, F.; Poucheret, P. Fruits and Vegetables, as a Source of Nutritional Compounds and Phytochemicals: Changes in Bioactive Compounds during Lactic Fermentation. Food Res. Int. 2018, 104, 86–99. [Google Scholar] [CrossRef]
- Michalak, M.; Kubik-Komar, A.; Waśko, A.; Polak-Berecka, M. Starter Culture for Curly Kale Juice Fermentation Selected using Principal Component Analysis. Food Biosci. 2020, 35, 100602. [Google Scholar] [CrossRef]
- Szutowska, J. Functional Properties of Lactic Acid Bacteria in Fermented Fruit and Vegetable Juices: A Systematic Literature Review. Eur. Food Res. Technol. 2020, 246, 357–372. [Google Scholar] [CrossRef]
- Chung, H.-J.; Lee, H.; Na, G.; Jung, H.; Kim, D.-G.; Shin, S.-I.; Jung, S.-E.; Choi, I.-D.; Lee, J.-H.; Sim, J.-H. Metabolic and Lipidomic Profiling of Vegetable Juices Fermented with Various Probiotics. Biomolecules 2020, 10, 725. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y. Production of Fermented Kale Juices with Lactobacillus Strains and Nutritional Composition. Prev. Nutr. Food Sci. 2017, 22, 231. [Google Scholar] [CrossRef] [PubMed]
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a Superfood: Review of the Scientific Evidence behind the Statement. Crit. Rev. Food Sci. Nutr. 2019, 59, 2411–2422. [Google Scholar] [CrossRef] [PubMed]
- Han, J.-H.; Lee, H.-J.; Kim, T.-S.; Kang, M.-H. The Effect of Glutathione S-Transferase M1 and T1 Polymorphisms on Blood Pressure, Blood Glucose, and Lipid Profiles following the Supplementation of Kale (Brassica oleracea acephala) Juice in South Korean Subclinical Hypertensive Patients. Nutr. Res. Pract. 2015, 9, 49–56. [Google Scholar] [CrossRef]
- Huang, Z.; Wang, B.; Eaves, D.H.; Shikany, J.M.; Pace, R.D. Phenolic Compound Profile of Selected Vegetables Frequently Consumed by African Americans in the Southeast United States. Food Chem. 2007, 103, 1395–1402. [Google Scholar] [CrossRef]
- Zielińska, M.; Lewandowska, U.; Podsędek, A.; Cygankiewicz, A.I.; Jacenik, D.; Sałaga, M.; Kordek, R.; Krajewska, W.M.; Fichna, J. Orally Available Extract from Brassica Oleracea var. Capitata Rubra Attenuates Experimental Colitis in Mouse Models of Inflammatory Bowel Diseases. J. Funct. Foods 2015, 17, 587–599. [Google Scholar] [CrossRef]
- Leroy, F.; De Vuyst, L. Lactic acid Bacteria as Functional Starter Cultures for the Food Fermentation Industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Tamang, J.P.; Cotter, P.D.; Endo, A.; Han, N.S.; Kort, R.; Liu, S.Q.; Mayo, B.; Westerik, N.; Hutkins, R. Fermented Foods in a Global Age: East Meets West. Compr. Rev. Food Sci. Food Saf. 2020, 19, 184–217. [Google Scholar] [CrossRef] [PubMed]
- Hess, J.M. Role of Lactic Acid Bacteria in Impacting Nutrient Bioavailability. Lact. Acid Bact. A Funct. Approach 2020, 3, 35–57. [Google Scholar]
- Jiang, X.; Liu, X.; Xu, H.; Sun, Y.; Zhang, Y.; Wang, Y. Improvement of the Nutritional, Antioxidant and Bioavailability Properties of Corn Gluten-Wheat Bran Mixture Fermented with Lactic Acid Bacteria and Acid Protease. LWT 2021, 144, 111161. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, L.; Wen, R.; Chen, Q.; Kong, B. Role of Lactic Acid Bacteria in Flavor Development in Traditional Chinese Fermented Foods: A Review. Crit. Rev. Food Sci. Nutr. 2022, 62, 2741–2755. [Google Scholar] [CrossRef] [PubMed]
- Khatami, M. Unresolved Inflammation: ‘Immune Tsunami’ or Erosion of Integrity in Immune-Privileged and Immune-Responsive Tissues and Acute and Chronic Inflammatory Diseases or Cancer. Expert Opin. Biol. Ther. 2011, 11, 1419–1432. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W. Chronic Inflammation in the Etiology of Disease across the Life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef]
- Shahinozzaman, M.; Raychaudhuri, S.; Fan, S.; Obanda, D.N. Kale Attenuates Inflammation and Modulates Gut Microbial Composition and Function in C57BL/6J Mice with Diet-Induced Obesity. Microorganisms 2021, 9, 238. [Google Scholar] [CrossRef] [PubMed]
- Hämäläinen, M.; Nieminen, R.; Asmawi, M.Z.; Vuorela, P.; Vapaatalo, H.; Moilanen, E. Effects of Flavonoids on Prostaglandin E2 Production and on COX-2 and mPGES-1 Expressions in Activated Macrophages. Planta Medica 2011, 77, 1504–1511. [Google Scholar] [CrossRef] [PubMed]
- De Marco, S.; Sichetti, M.; Muradyan, D.; Piccioni, M.; Traina, G.; Pagiotti, R.; Pietrella, D. Probiotic Cell-Free Supernatants Exhibited Anti-Inflammatory and Antioxidant Activity on Human Gut Epithelial Cells and Macrophages Stimulated with LPS. Evid.-Based Complement. Altern. Med. 2018, 2018, 1756308. [Google Scholar] [CrossRef] [PubMed]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: A Door to the Body. Front. Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef]
- Plaza-Díaz, J.; Ruiz-Ojeda, F.J.; Vilchez-Padial, L.M.; Gil, A. Evidence of the Anti-Inflammatory Effects of Probiotics and Synbiotics in Intestinal Chronic Diseases. Nutrients 2017, 9, 555. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Szwajgier, D.; Paduch, R.; Kukula-Koch, W.; Waśko, A.; Polak-Berecka, M. Fermented Curly Kale as a New Source of Gentisic and Salicylic Acids with Antitumor Potential. J. Func. Foods 2020, 67, 103866. [Google Scholar] [CrossRef]
- Seo, H.; Seong, H.; Kim, G.Y.; Jo, Y.M.; Cheon, S.W.; Song, Y.; Ryu, B.H.; Kang, H.; Han, N.S. Development of Anti-Inflammatory Probiotic Limosilactobacillus reuteri EFEL 6901 as Kimchi Starter: In vitro and In vivo Evidence. Front. Microb. 2021, 12, 760476. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.M.; Kim, G.Y.; Kim, S.-A.; Cheon, S.W.; Kang, C.-H.; Han, N.S. Limosilactobacillus fermentum MG7011: An Amylase and Phytase Producing Starter for the Preparation of Rice-based Probiotic Beverages. Front. Microb. 2021, 12, 745952. [Google Scholar] [CrossRef]
- Kang, H.; Moon, J.S.; Lee, M.-G.; Han, N.S. Immunomodulatory Effects of Leuconostoc citreum EFEL2061 isolated from Kimchi, a Traditional Korean Food, on the Th2 Type-Dominant Immune Response in vitro and in vivo. J. Func. Foods 2016, 20, 79–87. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.-C.; Choi, I.; Kim, G.-B. Effect of Fermentation on the Antioxidant Activity in Plant-based Foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Biglari, F.; AlKarkhi, A.F.; Easa, A.M. Antioxidant activity and Phenolic Content of Various Date Palm (Phoenix dactylifera) Fruits from Iran. Food Chem. 2008, 107, 1636–1641. [Google Scholar] [CrossRef]
- Hagen, S.F.; Borge, G.I.A.; Solhaug, K.A.; Bengtsson, G.B. Effect of Cold Storage and Harvest Date on Bioactive Compounds in Curly Kale (Brassica oleracea L. var. acephala). Postharvest Biol. Technol. 2009, 51, 36–42. [Google Scholar] [CrossRef]
- Rha, C.-S.; Seong, H.; Jung, Y.S.; Jang, D.; Kwak, J.-G.; Kim, D.-O.; Han, N.S. Stability and Fermentability of Green Tea Flavonols in in-vitro-Simulated Gastrointestinal Digestion and Human Fecal Fermentation. Int. J. Mol. Sci. 2019, 20, 35890. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.-S.; Lee, N.-K.; Choi, A.-J.; Choe, J.-S.; Bae, C.H.; Paik, H.-D. Anti-Inflammatory Potential of Probiotic Strain Weissella cibaria JW15 Isolated from Kimchi through Regulation of NF-κB and MAPKs Pathways in LPS-Induced RAW 264.7 cells. J. Microbiol. Biotechnol. 2019, 29, 1022–1032. [Google Scholar] [CrossRef]
- Cheon, S.; Kim, G.; Bae, J.H.; Lee, D.H.; Seong, H.; Kim, D.H.; Han, N.S. Comparative Analysis of Prebiotic Effects of Four Oligosaccharides using in vitro Gut Model: Digestibility, Microbiome, and Metabolome Changes. FEMS Microbiol. Ecol. 2023, 99, fiad002. [Google Scholar] [CrossRef]
- Tangyu, M.; Muller, J.; Bolten, C.J.; Wittmann, C. Fermentation of Plant-based Milk Alternatives for Improved Flavour and Nutritional Value. Appl. Microbiol. Biotechnol. 2019, 103, 9263–9275. [Google Scholar] [CrossRef] [PubMed]
- Dave, R.I.; Shah, N.P. Viability of Yoghurt and Probiotic Bacteria in Yoghurts Made from Commercial Starter Cultures. Int. Dairy J. 1997, 7, 31–41. [Google Scholar] [CrossRef]
- Kim, S.A.; Lee, D.H.; Ryu, B.H.; Han, N.S. Strain-Specific Barcode PCR and Quantitative PCR Assay for Identification and Enumeration of Kimchi Starter, Leuconostoc mesenteroides DRC1506. LWT 2023, 174, 114460. [Google Scholar] [CrossRef]
- Pinelo, M.; Arnous, A.; Meyer, A.S. Upgrading of Grape Skins: Significance of Plant Cell-Wall Structural Components and Extraction Techniques for Phenol Release. Trends Food Sci. Technol. 2006, 17, 579–590. [Google Scholar] [CrossRef]
- Martins, S.; Mussatto, S.I.; Martínez-Avila, G.; Montañez-Saenz, J.; Aguilar, C.N.; Teixeira, J.A. Bioactive Phenolic Compounds: Production and Extraction by Solid-State Fermentation. A Review. Biotechnol. Adv. 2011, 29, 365–373. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound Phenolics in Foods, a Review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yao, B.; Su, X. Linking Enzymatic Oxidative Degradation of Lignin to Organics Detoxification. Int. J. Mol. Sci. 2018, 19, 13373. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Rho, H.S.; Kim, D.H.; Chang, I.S. Enzymatic Preparation of Kaempferol from Green Tea Seed and its Antioxidant Activity. J. Agric. Food Chem. 2006, 54, 2951–2956. [Google Scholar] [CrossRef] [PubMed]
- Nguyen Thai, H.; Van Camp, J.; Smagghe, G.; Raes, K. Improved Release and Metabolism of Flavonoids by Steered Fermentation Processes: A Review. Int. J. Mol. Sci. 2014, 15, 19369–19388. [Google Scholar] [CrossRef]
- Gan, R.Y.; Shah, N.P.; Wang, M.F.; Lui, W.Y.; Corke, H. Lactobacillus plantarum WCFS1 Fermentation Differentially Affects Antioxidant Capacity and Polyphenol Content in Mung Bean (Vigna radiata) and Soya Bean (Glycine max) milks. J. Food Process Preserv. 2017, 41, e12944. [Google Scholar] [CrossRef]
- Landete, J.M.; Curiel, J.A.; Rodríguez, H.; de las Rivas, B.; Munoz, R. Aryl Glycosidases from Lactobacillus plantarum Increase Antioxidant Activity of Phenolic Compounds. J. Func. Foods 2014, 7, 322–329. [Google Scholar] [CrossRef]
- Lee, Y.G.; Cho, J.Y.; Kim, Y.M.; Moon, J.H. Change in Flavonoid Composition and Antioxidative Activity during Fermentation of Onion (Allium cepa L.) by Leuconostoc mesenteroides with Different Salt Concentrations. J. Food Sci. 2016, 81, C1385–C1393. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, S.M.; Chua, L.S.; El-Enshasy, H.A.; Abd Majid, F.A.; Hanapi, S.Z. Kinetic profile and anti-diabetic potential of fermented Punica granatum juice using Lactobacillus casei. Process Biochem. 2020, 92, 224–231. [Google Scholar] [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of Lactobacillus Strains on Phenolic Profile, Color Attributes and Antioxidant Activities of Lactic-Acid-Fermented Mulberry Juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef]
- Stübler, A.-S.; Lesmes, U.; Heinz, V.; Rauh, C.; Shpigelman, A.; Aganovic, K. Digestibility, Antioxidative Activity and Stability of Plant Protein-Rich Products after Processing and Formulation with Polyphenol-Rich Juices: Kale and Kale–Strawberry as a Model. Eur. Food Res. Technol. 2019, 245, 2499–2514. [Google Scholar] [CrossRef]
- Jeganathan, B.; Punyasiri, P.; Kottawa-Arachchi, J.D.; Ranatunga, M.A.; Abeysinghe, I.S.B.; Gunasekare, M.; Bandara, B. Genetic Variation of Flavonols Quercetin, Myricetin, and Kaempferol in the Sri Lankan Tea (Camellia sinensis L.) and Their Health-Promoting Aspects. Int. J. Food Sci. 2016, 2016, 6057434. [Google Scholar] [CrossRef] [PubMed]
- Lampiasi, N.; Montana, G. The Molecular Events behind Ferulic Acid Mediated Modulation of IL-6 Expression in LPS-Activated Raw 264.7 Cells. Immunobiology 2016, 221, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Blonska, M.; Czuba, Z.; Krol, W. Effect of Flavone Derivatives on Interleukin-1β (IL-1β) mRNA Expression and IL-1β Protein Synthesis in Stimulated RAW 264.7 Macrophages. Scand. J. Immunol. 2003, 57, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Saiki, P.; Nakajima, Y.; Van Griensven, L.J.; Miyazaki, K. Real-Time Monitoring of IL-6 and IL-10 Reporter Expression for Anti-Inflammation Activity in Live RAW 264.7 Cells. Biochem. Biophys. Res. Commun. 2018, 505, 885–890. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Collection | Abbreviation | Culture Condition |
|---|---|---|---|
| Limosilactobacillus reuteri EFEL6901 | KACC 81105BP | EFEL6901 | 37 °C, MRS |
| L. fermentum EFEL6800 | KACC 81106BP | EFEL6800 | 37 °C, MRS |
| L. fermentum MG7011 | KACC 81147BP | MG7011 | 37 °C, MRS |
| Leuconostoc citreum EFEL2061 | KACC 92070P | EFEL2601 | 37 °C, MRS |
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Gene Accession Code |
|---|---|---|---|
| GAPDH | TTGTCTCCTGCGACTTCAACA | GCTGTAGCCGTATTCATTGTCATA | XM_059184619.1 |
| iNOS | ACCATGGAGCATCCCAAGTA | CCATGTACCAACCATTGAAGG | NM_010927.4 |
| COX-2 | AGCATTCATTCCTCTACATAAGC | GTAACAACACTCACATATTCATACAT | AH002100.2 |
| IL-6 | GAGGATACCACTCCCAACAG | AAGTGCATCATCGTTGTTCATA | BC132458.1 |
| IL-1β | GTTGACGGACCCCAAAAGAT | CACACACCAGCAGGTTATCA | M15131.1 |
| IL-10 | GGACAACATACTGCTAACCGACTC | AAAATCACTCTTCACCTGCTCCAC | XM_036162094.1 |
| Sample | L* | a* | b* | ΔE* |
|---|---|---|---|---|
| Kale juice | 29.95 ± 0.02 c | −6.56 ± 0.02 e | 10.91 ± 0.02 a | 6.30 ± 0.04 a |
| PKJ | 34.61 ± 0.00 a | −6.80 ± 0.01 e | 15.14 ± 0.03 b | - |
| KJ-EFEL6901 | 34.02 ± 0.01 b | −0.01 ± 0.01 a | 13.73 ± 0.02 b | 6.80 ± 0.02 c |
| KJ-EFEL6800 | 34.59 ± 0.03 a | −1.36 ± 0.01 d | 15.25 ± 0.09 b | 5.67 ± 0.04 c |
| KJ-MG7011 | 34.61 ± 0.04 a | −0.76 ± 0.04 b | 15.70 ± 1.44 b | 6.49 ± 0.47 b |
| KJ-EFEL2061 | 34.64 ± 0.02 a | −0.85 ± 0.01 c | 14.65 ± 0.01 b | 6.07 ± 0.04 c |
| Group | Metabolites | PKJ | KJ-EFEL6901 | KJ-EFEL6800 | KJ-MG7011 | KJ-EFEL2061 |
|---|---|---|---|---|---|---|
| Carbohydrates | Fructose | 0.65 ± 0.12 a | 0.08 ± 0.01 b | 0.20 ± 0.03 b | 0.14 ± 0.00 b | 0.04 ± 0.00 b |
| Glucose | 0.90 ± 0.04 a | 0.06 ± 0.01 b | 0.06 ± 0.01 b | 0.05 ± 0.00 b | 0.04 ± 0.00 b | |
| Mannose | 0.16 ± 0.05 a | 0.09 ± 0.01 b | 0.13 ± 0.04 b | 0.09 ± 0.00 b | 0.07 ± 0.00 b | |
| Sucrose | 0.14 ± 0.01 a | 0.02 ± 0.00 b | 0.05 ± 0.04 b | 0.05 ± 0.00 b | 0.02 ± 0.00 b | |
| Alcohols | Mannitol | 0.10 ± 0.01 ab | 1.23 ± 0.03 ab | 1.14 ± 0.05 a | 1.10 ± 0.02 a | 0.98 ± 0.02 ab |
| Ethanol | 0.24 ± 0.01 c | 1.74 ± 0.06 b | 2.98 ± 0.24 a | 2.25 ± 0.05 b | 1.42 ± 0.01 ab | |
| Organic acids | Acetate | 0.19 ± 0.01 d | 3.24 ± 0.33 ab | 3.54 ± 0.11 a | 2.63 ± 0.13 c | 1.76 ± 0.25 b |
| Succinate | 0.09 ± 0.01 c | 0.85 ± 0.04 b | 0.30 ± 0.01 c | 1.30 ± 0.17 a | 1.13 ± 0.03 a | |
| Lactate | 0.09 ± 0.01 d | 1.69 ± 0.21 a | 1.86 ± 0.03 a | 0.97 ± 0.01 c | 0.49 ± 0.02 b | |
| Propionate | 0.03 ± 0.01 c | 0.54 ± 0.00 b | 0.71 ± 0.06 b | 0.63 ± 0.04 b | 0.84 ± 0.03 a | |
| Pyruvate | 0.07 ± 0.01 b | 0.08 ± 0.04 a | 0.07 ± 0.01 a | 0.06 ± 0.01 a | 0.04 ± 0.00 ab | |
| Butyrate | 0.04 ± 0.00 c | 0.30 ± 0.01 b | 0.18 ± 0.01 a | 0.23 ± 0.40 a | 0.19 ± 0.01 a | |
| Citrate | 0.12 ± 0.02 a | 0.02 ± 0.01 b | 0.04 ± 0.01 b | 0.02 ± 0.00 b | 0.01 ± 0.00 b | |
| Amino acids | Aspartate | 1.40 ± 0.04 b | 1.08 ± 0.08 a | 0.04 ± 0.01 c | 0.03 ± 0.01 c | 0.06 ± 0.01 c |
| Glutamate | 1.13 ± 0.11 b | 1.35 ± 0.07 a | 0.33 ± 0.02 c | 0.27 ± 0.01 c | 0.06 ± 0.00 d | |
| Cysteine | 0.06 ± 0.01 b | 0.17 ± 0.01 a | 0.12 ± 0.03 b | 0.12 ± 0.00 b | 0.07 ± 0.00 bc | |
| Glycine | 0.08 ± 0.03 bc | 0.13 ± 0.01 ab | 0.16 ± 0.02 a | 0.18 ± 0.02 a | 0.06 ± 0.00 c | |
| Histidine | 0.01 ± 0.00 a | 0.01 ± 0.01 a | 0.01 ± 0.00 a | 0.00 ± 0.00 a | 0.01 ± 0.00 a | |
| Alanine | 0.91 ± 0.03 a | 0.58 ± 0.01 b | 0.62 ± 0.13 b | 0.48 ± 0.03 b | 0.09 ± 0.01 c | |
| Serine | 1.18 ± 0.21 a | 0.09 ± 0.01 b | 0.12 ± 0.01 b | 0.12 ± 0.01 b | 0.05 ± 0.01 b | |
| Threonine | 0.30 ± 0.03 a | 0.02 ± 0.00 c | 0.08 ± 0.04 c | 0.18 ± 0.03 b | 0.03 ± 0.00 c | |
| Arginine | 0.78 ± 0.04 a | 0.05 ± 0.01 b | 0.03 ± 0.01 b | 0.02 ± 0.00 b | 0.07 ± 0.00 b | |
| Proline | 0.44 ± 0.15 a | 0.37 ± 0.10 a | 0.26 ± 0.03 a | 0.30 ± 0.04 a | 0.17 ± 0.00 a | |
| Tyrosine | 0.19 ± 0.01 a | 0.15 ± 0.00 b | 0.03 ± 0.00 c | 0.19 ± 0.01 a | 0.06 ± 0.00 d | |
| Valine | 0.53 ± 0.21 abc | 0.47 ± 0.08 bc | 0.07 ± 0.01 a | 0.40 ± 0.01 c | 0.20 ± 0.01 ab | |
| Methionine | 0.08 ± 0.01 a | 0.06 ± 0.01 a | 0.02 ± 0.01 b | 0.01 ± 0.00 b | 0.02 ± 0.00 b | |
| Isoleucine | 0.43 ± 0.05 a | 0.29 ± 0.02 b | 0.05 ± 0.04 c | 0.11 ± 0.01 c | 0.05 ± 0.01 c | |
| Leucine | 0.48 ± 0.02 a | 0.31 ± 0.07 b | 0.14 ± 0.00 c | 0.16 ± 0.00 c | 0.10 ± 0.01 c | |
| Phenylalanine | 0.56 ± 0.05 a | 0.16 ± 0.01 c | 0.05 ± 0.03 cd | 0.38 ± 0.01 b | 0.10 ± 0.01 c | |
| Tryptophan | 0.23 ± 0.02 a | 0.21 ± 0.01 a | 0.07 ± 0.00 b | 0.14 ± 0.01 c | 0.07 ± 0.00 b | |
| Lysine | 0.63 ± 0.03 b | 0.43 ± 0.05 c | 0.30 ± 0.02 c | 0.70 ± 0.01 b | 0.09 ± 0.00 a |
| Related Enzyme | Gene Annotation | Predicted Mechanism | Protein ID | |
|---|---|---|---|---|
| EFEL6901 | EFEL6800 | |||
| Glycoside hydrolase | Glycoside hydrolase family 13 protein | hydrolysis of α-glucoside linkages | WP_003668994.1 | WP_023466491.1 |
| Glycoside hydrolase family 65 protein | hydrolysis of α-glucosidic linkages (mainly phosphorylase) | WP_003669571.1 | WP_023465690.1 | |
| Glycoside hydrolase family 70 protein | hydrolysis of glycosidic linkages (transfer D-glucopyramnosyl units from sucrose onto acceptor molecules) | WP_229392148.1 | ||
| Glycoside hydrolase family 73 protein | hydrolysis of β-1,4-glycosidic linkage between N-acetylglucosaminyl (NAG) and N-acetylmuramyl (NAM) moieties | WP_003674860.1 WP_003669289.1 | WP_023466547.1 WP_003681382.1 WP_004562972.1 | |
| Glycoside hydrolase family 2 TIM barrel-domain containing protein | hydrolysis of β-glycosidic linkages (broad spectrum) | WP_003667314.1 | WP_282347715.1 WP_282348494.1 | |
| Glycoside hydrolase family 3 N-terminal domain-containing protein | hydrolysis single glycosyl residues from the non-reducing ends of substrate (exo type, broad spectrum) | - | WP_075667436.1 WP_240824081.1 | |
| Family 78 glycoside hydrolase catalytic domain | hydrolysis of α-L-rhamnosyl-linkages in L-rhamnosides | - | WP_282348481.1 | |
| Glucosidase | Alpha-glucosidase | hydrolysis of α-glucoside linkages | WP_003669620.1 | WP_023466314.1 |
| Glycosyl hydrolase 53 family protein | β-1,4-galactanase | WP_003668597.1 | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, G.Y.; Kim, S.-A.; Kong, S.Y.; Seong, H.; Bae, J.-H.; Han, N.S. Synergistic Antioxidant and Anti-Inflammatory Activities of Kale Juice Fermented with Limosilactobacills reuteri EFEL6901 or Limosilactobacills fermentum EFEL6800. Antioxidants 2023, 12, 1850. https://doi.org/10.3390/antiox12101850
Kim GY, Kim S-A, Kong SY, Seong H, Bae J-H, Han NS. Synergistic Antioxidant and Anti-Inflammatory Activities of Kale Juice Fermented with Limosilactobacills reuteri EFEL6901 or Limosilactobacills fermentum EFEL6800. Antioxidants. 2023; 12(10):1850. https://doi.org/10.3390/antiox12101850
Chicago/Turabian StyleKim, Ga Yun, Seul-Ah Kim, Sun Young Kong, Hyunbin Seong, Jae-Han Bae, and Nam Soo Han. 2023. "Synergistic Antioxidant and Anti-Inflammatory Activities of Kale Juice Fermented with Limosilactobacills reuteri EFEL6901 or Limosilactobacills fermentum EFEL6800" Antioxidants 12, no. 10: 1850. https://doi.org/10.3390/antiox12101850
APA StyleKim, G. Y., Kim, S.-A., Kong, S. Y., Seong, H., Bae, J.-H., & Han, N. S. (2023). Synergistic Antioxidant and Anti-Inflammatory Activities of Kale Juice Fermented with Limosilactobacills reuteri EFEL6901 or Limosilactobacills fermentum EFEL6800. Antioxidants, 12(10), 1850. https://doi.org/10.3390/antiox12101850