
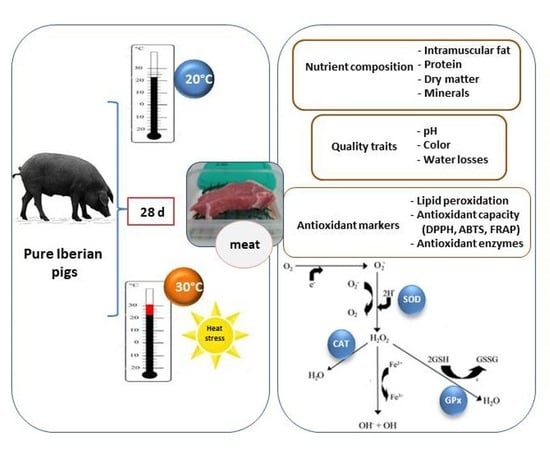
Impact of Heat Stress on Meat Quality and Antioxidant Markers in Iberian Pigs

 and
and 
Abstract
:
1. Introduction
2. Materials and Methods
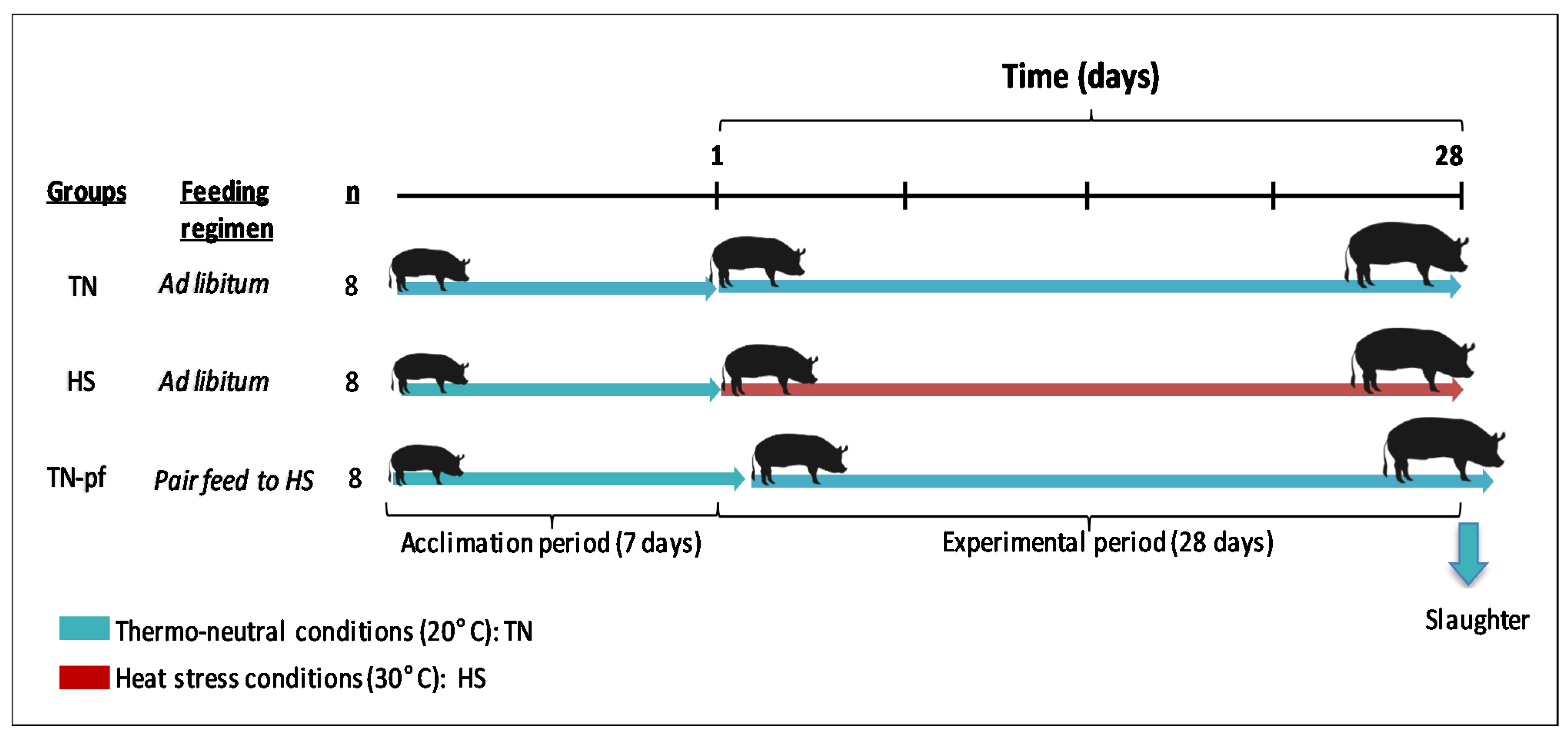
2.1. Animals and Experimental Design
2.2. Muscle Quality Traits
2.2.1. Physical Meat Quality Assessment
2.2.2. Chemical Composition
2.2.3. Fatty Acid (FA) Analysis
2.3. Antioxidant Status
2.3.1. Lipid Peroxidation
2.3.2. Antioxidant Activity
2.3.3. Antioxidant Enzyme Activity
2.4. Statistical Analysis
3. Results and Discussion
3.1. Chemical Composition and FA Profile of Muscles
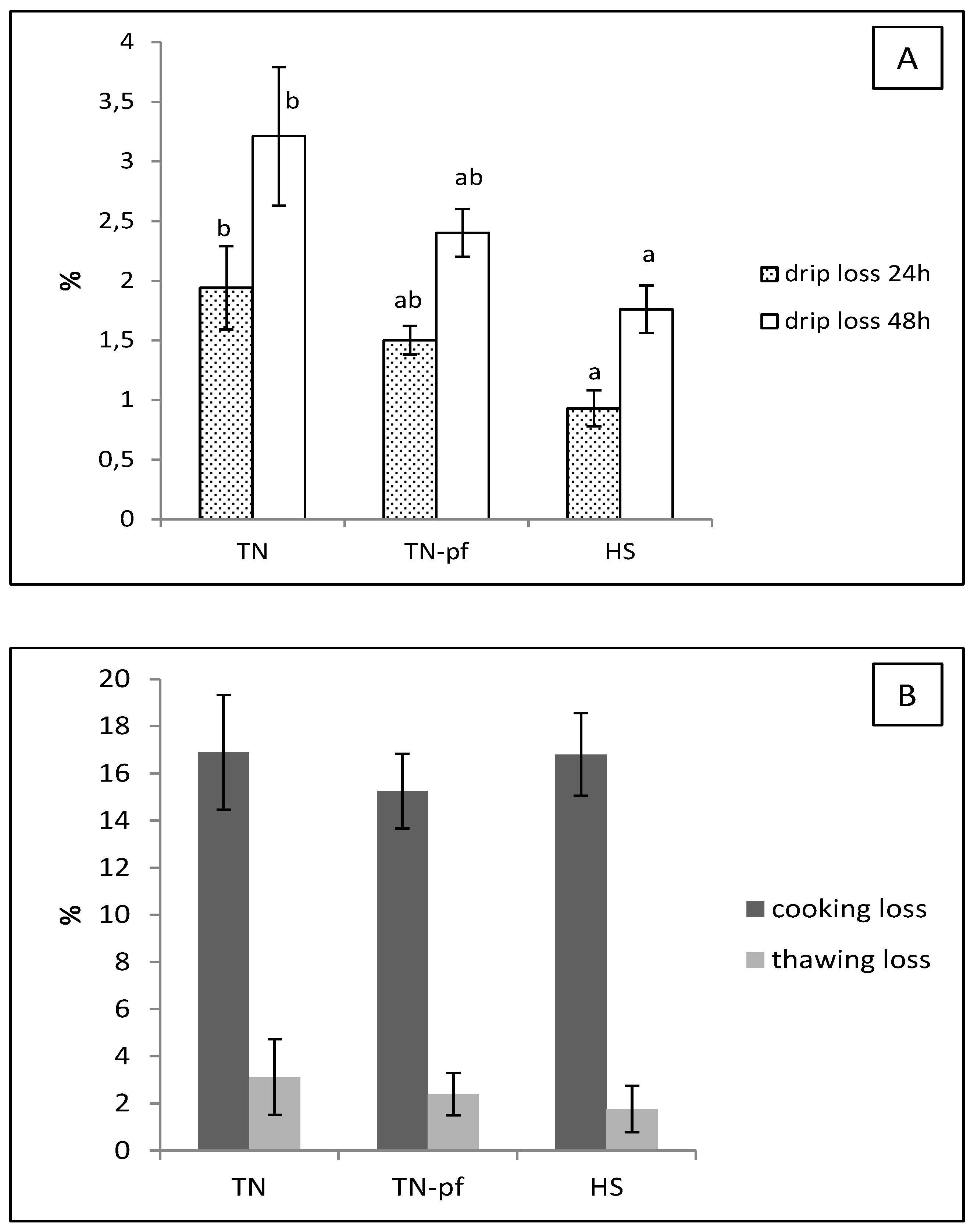
3.2. pH, Color and Water Holding Capacity
3.3. Antioxidant Markers
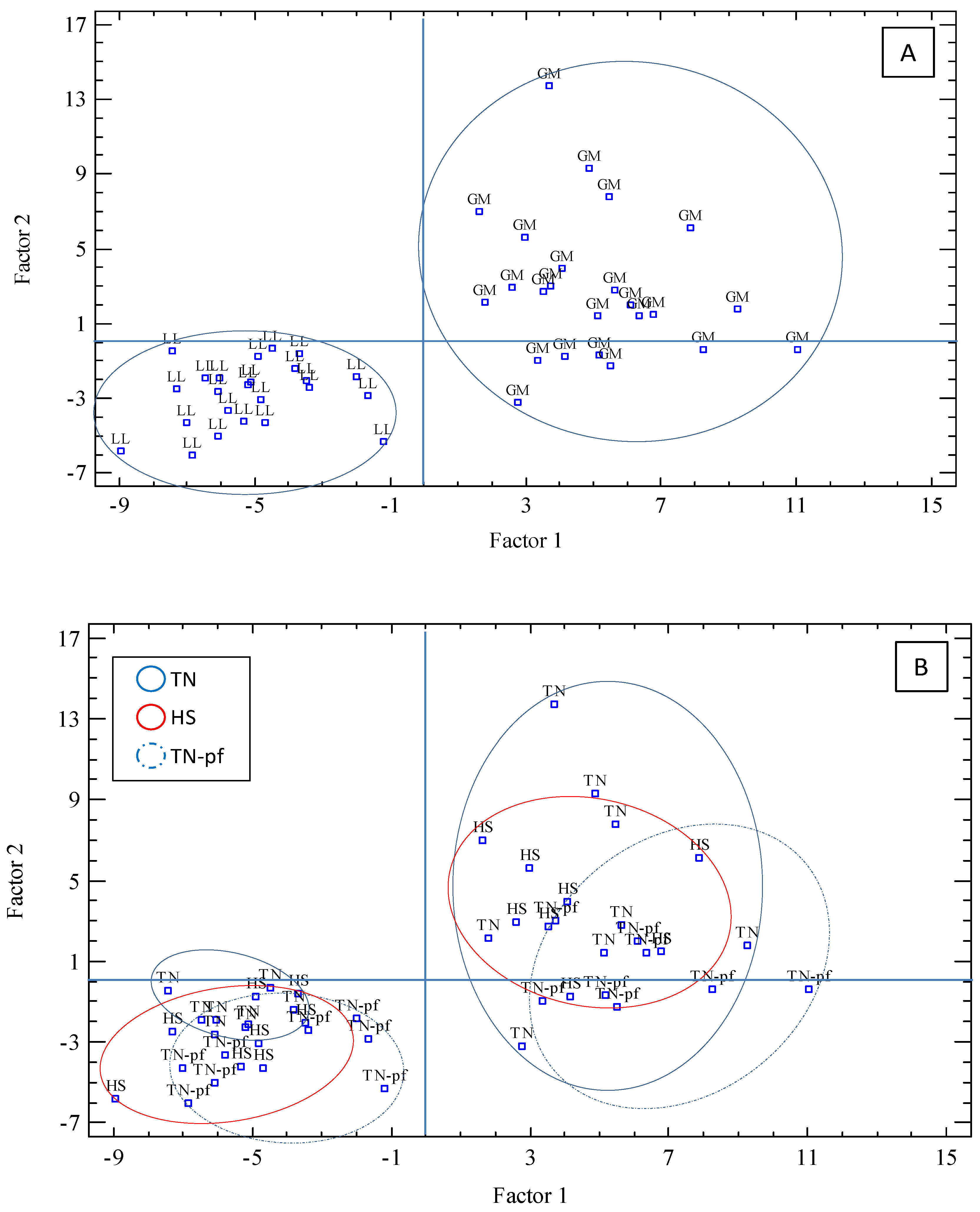
3.4. Factor Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NASA. Global Climate Change, Vital Signs of the Planet. 2021. Available online: https://climate.nasa.gov/evidence/ (accessed on 17 June 2021).
- Rojas-Downing, M.; Nejadhashemi, A.P.; Harrigan, T.; Woznicki, S.A. Climate change and livestock: Impacts, adaptation, and mitigation. Clim. Risk. Manag. 2017, 16, 145–163. [Google Scholar] [CrossRef]
- Ma, X.; Jiang, Z.; Zheng, C.; Hu, Y.; Wang, L. Nutritional regulation for meat quality and nutrient metabolism of pigs exposed to high temperature environment. J. Food Sci. Nutr. 2015, 5, 5–6. [Google Scholar] [CrossRef]
- Čobanović, N.; Stajković, S.; Blagojević, B.; Betić, N.; Vasilev, D.D.; Karabasil, N. The effect of season on health, welfare, and carcass and meat quality of slaughter pigs. Int. J. Biometeorol. 2020, 64, 1899–1909. [Google Scholar] [CrossRef]
- Zhang, M.; Dunshea, F.R.; Warner, R.D.; DiGiacomo, K.; Amponsah, R.O.; Chauchan, S.S. Impacts of heat stress on meat quality and strategies for amelioration: A review. Int. J. Biometeorol. 2020, 64, 1613–1628. [Google Scholar] [CrossRef]
- FAO. How to Feed the World in 2050. Available online: https://www.fao.org/fileadmin/templates/wsfs/docs/expert_paper/How_to_Feed_the_World_in_2050.pdf (accessed on 21 October 2021).
- Renaudeau, D.; Leclercq-smekens, M.; Herin, M. Differences in skin characteristics in European (Large White) and Caribbean (Creole) growing pigs with reference to thermoregulation. Anim. Res. 2006, 55, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Ross, J.W.; Hale, B.J.; Gabler, N.K.; Rhoads, R.P.; Keating, A.F.; Baumgard, L.H. Physiological consequences of heat stress in pigs. Anim. Prod. Sci. 2015, 55, 1381–1390. [Google Scholar] [CrossRef]
- Liu, Y.; Yin, S.; Tang, J.; Liu, Y.; Jia, G.; Liu, G.; Tian, G.; Chen, X.; Cai, J.; Kang, B.; et al. Hydroxy selenomethionine improves meat quality through optimal skeletal metabolism and functions of selenoproteins of pigs under chronic heat stress. Antioxidants 2021, 10, 1558. [Google Scholar] [CrossRef]
- Rosado Montilla, S.I.; Johnson, T.P.; Pearce, S.C.; Gardan-Salmon, D.; Gabler, N.K.; Ross, J.W.; Rhoads, R.P.; Baumgard, L.H.; Lonergan, S.M.; Selsby, J.T. Heat stress causes oxidative stress but not inflammatory signaling in porcine skeletal muscle. Temperature 2014, 1, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.; Hao, Y.; Feng, J.; Lin, H.; Feng, Y.; Wu, X.; Yang, X.; Gu, X. The expression of carnosine and its effect on the antioxidant capacity of longissimus dorsi muscle in finishing pigs exposed to constant heat stress. Asian-Australas. J. Anim. Sci. 2014, 27, 1763–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinaldo, D.; Mourot, J. Effects of tropical climate and season on growth chemical composition of muscle and adipose tissue and meat quality in pigs. Anim. Res. 2001, 50, 507–521. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Wang, L.; Shi, Z.; Chen, W.; Yang, X.; Hu, Y.; Zheng, C.; Jiang, Z. Mechanism of continuous high temperature affecting growth performance, meat quality, and muscle biochemical properties of finishing pigs. Genes Nutr. 2019, 14, 23. [Google Scholar] [CrossRef]
- Seiquer, I.; Palma-Granados, P.; Haro, A.; Lara, L.; Lachica, M.; Fernández-Fígares, I.; Nieto, R. Meat quality traits in longissimus lumborum and gluteus medius muscles from immunocastrated and surgically castrated Iberian pigs. Meat Sci. 2019, 150, 77–84. [Google Scholar] [CrossRef] [PubMed]
- McLellan, K.; Petrofsky, J.S.; Bains, G.; Zimmerman, G.; Prowse, M.; Lee, S. The effects of skin moisture and subcutaneous fat thickness on the ability of the skin to dissipate heat in young and old subjects, with and without diabetes, at three environmental room temperatures. Med. Eng. Phys. 2009, 31, 165–172. [Google Scholar] [CrossRef]
- Climate Data. Available online: http://en.climate-data.org/europe/spain-5/ (accessed on 21 October 2021).
- Usala, M.; Macciotta, N.P.P.; Bergamaschi, M.; Maltecca, C.; Fix, J.; Schwab, C.; Shull, C.; Tlezzi, F. Genetic parameters for tolerance to heat stress in crossbred swine carcass traits. Front. Genet. 2021, 11, 612815. [Google Scholar] [CrossRef]
- Nieto, R.; Martínez-Pérez, M.; Haro, A.; Lara, L.; Aguilera, J.F. Effects of protein intake on rate of growth, protein deposition, and carcass traits of heavy Iberian pigs. J. Anim. Sci. 2012, 90, 3809–3820. [Google Scholar] [CrossRef] [PubMed]
- Font-i-Furnols, M.; Čandek-Potokar, M.; Povše, M.P. A Handbook of Reference Methods for Meat Quality Assessment; European Cooperation in Science and Technology (COST): Brussels, Belgium, 2015. [Google Scholar]
- AOAC. Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2000. [Google Scholar]
- Palma-Granados, P.; Haro, A.; Seiquer, I.; Lara, L.; Aguilera, J.F.; Nieto, R. Similar effects of lysine deficiency in muscle biochemical characteristics of fatty and lean piglets. J. Anim. Sci. 2017, 95, 3025–3036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Kramer, J.K.G.; Zhou, J. Conjugated linoleic acid and octadecenoic acids: Extraction and isolation of lipids. Eur. J. Lipid Sci. Technol. 2001, 103, 594–600. [Google Scholar] [CrossRef]
- Seiquer, I.; Rueda, A.; Olalla, M.; Cabrera-Vique, C. Assesing the biobailability of polyphenols and antioxidant properties of extra virgin argan oil by simulated digestión and Caco-2 cell assays. Comparative study with extra virgin olive oil. Food Chem. 2015, 188, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Pardo, Z.; Seiquer, I. Supplemental zinc exerts a positive effect against the heat stress damage in intestinal epithelial cells: Assays in a Caco-2 model. J. Funct. Foods 2021, 83, 104569. [Google Scholar] [CrossRef]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Palma-Granados, P.; Haro, A.; Lara, L.; Aguilera, J.F.; Nieto, R.; Seiquer, I. Differences on meat color and composition between Landrace x Large White and Iberian ppigs under identical nutritional and management conditions. Anim. Prod. Sci. 2018, 58, 2132–2142. [Google Scholar] [CrossRef]
- Gonzalez-Prendes, R.; Quintanilla, R.; Mármol-Sánchez, E.; Pena, R.N.; Ballester, M.; Cardoso, T.F.; Manunza, A.; Casellas, J.; Cánovas, A.; Díaz, I.; et al. Comparing the mRNA expression profile and the genetic expression profile and the genetic determinism of intramuscular fat traits in the porcine gluteus medius and longissimus dorsi muscles. BMC Genomics. 2019, 20, 170. [Google Scholar] [CrossRef] [PubMed]
- Rinaldo, D.; Dividich, J.L. Effects of warm exposure on adipose tissue and muscle metabolism in growing pigs. Comp. Biochem. Physiol. 1991, 4, 995–1002. [Google Scholar] [CrossRef]
- Shi, Z.B.; Ma, X.Y.; Zheng, C.T.; Hu, Y.J.; Yang, X.F.; Gao, K.G.; Wang, L.; Jiang, Z.Y. Effects of high ambient temperature on meat quality, serum hormone concentrations, and gene expression in the longissimus dorsi muscle of finishing pigs. Anim. Prod. Sci. 2016, 57, 1031–1039. [Google Scholar] [CrossRef]
- Xin, W.U.; Li, Z.Y.; Jia, A.F.; Su, H.G.; Hu, C.H.; Zhang, M.H.; Feng, J.H. Effects of high ambient temperature on lipid metabolism in finishing pigs. J. Integr. Agric. 2016, 15, 391–396. [Google Scholar]
- Lu, Z.; He, X.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Chronic heat stress impairs the quality of breast muscle meat in broilers by affecting redox status and energy substance metabolism. J. Agric. Food. Chem. 2017, 65, 11251–11258. [Google Scholar] [CrossRef]
- Cardoso-Pereira, P.M.C.; Baltazar-Vicente, A.F.R. Meat nutritional composition and nutritive role in the human diet. Meat Sci. 2013, 93, 586–592. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.D.; Enser, M. Factors influencing fatty acids in meat and the role of antioxidants in improving meat quality. Brit. J. Nutr. 1997, 78, S49–S60. [Google Scholar] [CrossRef] [Green Version]
- Rant, W.; Radzik-rant, A.; Swiatek, M.; Niżnikowski, R.; Szymańska, Ż.; Bednarczyk, M.; Orlowski, E.; Morales-Villavicencio, A.; Ślęzak, M. The effect of aging and muscle type on the quality characteristics and lipid oxidation of lamb meat. Arch. Anim. Breed. 2019, 62, 383–391. [Google Scholar] [CrossRef] [Green Version]
- Andrés, A.I.; Cava, R.; Mayoral, A.I.; Tejeda, J.F.; Morcuende, D.; Ruiz, J. Oxidative stability and fatty acid composition of pig muscles as affected by rearing system, crossbreeding and metabolic type of muscle fibre. Meat Sci. 2001, 59, 39–47. [Google Scholar] [CrossRef]
- González-Domínguez, R.; Sayago, A.; Fernández-Recamales, A. Fatty acid profiling for the authentication of Iberian hams according to the feeding regime. Foods 2020, 9, 149. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.P.; Feng, D.Y.; Xiao, J.; Wu, F.; He, X.J.; Xia, M.H.; Dong, T.; Liu, Y.H.; Tan, H.Z.; Zou, S.G.; et al. Studies on meat color, myoglobin content, enzyme activities, and genes associated with oxidative potential of pigs slaughtered at different growth stages. Asian-Australas. J. Anim. Sci. 2017, 30, 1739–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonetti, A.; Perna, A.; Giudice, R.; Cappucio, A.; Gambacorta, E. The effect of high pre-slaughter environmental temperature on meat quality traits of Italian autochthonous pig Suino Nero Lucano. Anim. Sci. J. 2018, 89, 1020–1026. [Google Scholar] [CrossRef]
- Cui, Y.; Hao, Y.; Li, J.; Gao, Y.; Gu, X. Proteomic changes of the porcine skeletal muscle in response to chronic heat stress. J. Sci. Food Agric. 2018, 98, 3315–3323. [Google Scholar] [CrossRef] [PubMed]
- Mancini, R.A.; Hunt, M.C. Current research in meat color. Meat Sci. 2005, 71, 100–121. [Google Scholar] [CrossRef]
- Lindahl, G.; Lundstrom, K.; Tornberg, E. Contribution of pigment content myoglobin forms and internal reflectance to the colour of pork loin and ham from pure breed pigs. Meat Sci. 2001, 59, 141–151. [Google Scholar] [CrossRef]
- Parkunan, T.; Das, A.K.; Banerjee, D.; Mohanty, N.; Paul, A.; Nanda, P.K.; Biswas, T.K.; Naskar, S.; Bag, S.; Sarkar, M.; et al. Changes in expression of monocarboxylate transporters, heat shock proteins and meat quality of Large White Yorkshire and Ghungroo pigs during hot summer period. Asian-Australas. J. Anim. Sci. 2017, 30, 246–253. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, L.; Zhang, L.; Ying, Z.; Su, W.; Wang, T. Curcumin attenuates d-galactosamine/lipopolysaccharide-induced liver injury and mitochondrial dysfunction in mice. J. Nutr. 2014, 144, 1211–1218. [Google Scholar] [CrossRef] [Green Version]
- Hosseindoust, A.; Oh, S.M.; Ko, H.S.; Jeon, S.M.; Ha, S.H.; Jang, A.; Son, J.S.; Kim, G.Y.; Kang, H.K.; Kim, J.S. Muscle antioxidant activity and meat quality are altered by supplementation of astaxanthin in broilers exposed to high temperature. Antioxidants 2020, 9, 1032. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, Y.; Du, M.; Zhou, Y. Protective effects of dietary symbiotic supplementation on meat quality and oxidative status in broilers under heat stress. Environ. Sci. Pollut. Res. 2021, 28, 30197–30206. [Google Scholar] [CrossRef]
- Echegaray, N.; Munekata, P.E.S.; Centeno, J.A.; Domínguez, R.; Pateiro, M.; Carballo, J.; Lorenzo, J.M. Total phenol content and antioxidant activity of different celta pig carcass locations as affected by the finishing diet (Chestnuts or commercial feed). Antioxidants 2020, 10, 5. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Xiong, Y.; Yi, H.; Wu, Q.; Jiang, Z.; Wang, L. Oxidative stress and endoplasmic reticulum stress are involved in the protective effect of alpha lipoic acid against heat damage in chicken testes. Animals 2020, 10, 384. [Google Scholar] [CrossRef] [Green Version]
- Tejerina, D.; García-Torres, S.; Cabeza de Vaca, M.; Vázquez, F.M.; Cava, R. Effect of production system on physical-chemical, antioxidant and fatty acids composition of Longissimus dorsi and Serratus ventralis muscles from Iberian pig. Food Chem. 2012, 133, 293–299. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Longissimus | Gluteus | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TN | TN-pf | HS | TN | TN-pf | HS | SEM 1 | Treatment (T) | Muscle (M) | T × M | |
| Dry matter (%) | 27.6b | 25.9a | 26.4a | 30.4b | 27.9a | 30.1b | 0.21 | 0.0008 | 0.0001 | 0.2602 |
| Protein (%) | 21.6 | 22.1 | 21.2 | 23.6 | 23.5 | 23.9 | 0.35 | 0.9581 | 0.0052 | 0.7369 |
| IMF 2 (%) | 6.61b | 4.77a | 5.87b | 9.69b | 5.91a | 8.15b | 0.21 | 0.0027 | 0.0012 | 0.4516 |
| Ash (%) | 1.97 | 1.95 | 2.04 | 1.59 | 1.76 | 1.67 | 0.05 | 0.7930 | 0.0045 | 0.7226 |
| Energy (kcal/100 g) | 170b | 153a | 160a | 196b | 168a | 189ab | 2.3 | 0.0013 | 0.0001 | 0.3963 |
| Fe (mg/kg) | 7.05 | 7.82 | 7.43 | 9.08 | 10.0 | 10.1 | 0.28 | 0.4120 | 0.0002 | 0.8893 |
| Zn (mg/kg) | 15.4a | 14.4a | 16.9b | 16.2a | 15.2a | 18.9b | 0.26 | 0.0001 | 0.0288 | 0.5486 |
| Longissimus | Gluteus | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TN | TN-pf | HS | TN | TN-pf | HS | SEM 1 | Treatment (T) | Muscle (M) | T × M | |
| C12:0 | 0.065 | 0.054 | 0.061 | 0.061 | 0.053 | 0.061 | 0.002 | 0.1184 | 0.6624 | 0.9016 |
| C14:0 | 1.47 | 1.39 | 1.49 | 1.34 | 1.31 | 1.43 | 0.02 | 0.1090 | 0.0313 | 0.7780 |
| C16:0 | 27.2 | 27.4 | 27.9 | 26.6 | 26.7 | 27.6 | 0.01 | 0.2042 | 0.2051 | 0.9585 |
| C16:1 | 3.91 | 3.73 | 3.73 | 3.37 | 3.36 | 3.40 | 0.07 | 0.8480 | 0.0074 | 0.8163 |
| C17:0 | 0.26 | 0.26 | 0.27 | 0.32 | 0.31 | 0.31 | 0.01 | 0.9688 | 0.0223 | 0.9025 |
| C18:0 | 12.2 | 12.2 | 12.9 | 12.7 | 12.1 | 13.2 | 0.13 | 0.0248 | 0.3586 | 0.6256 |
| C18:1n9 | 43.1 | 41.9 | 41.0 | 41.3 | 41.3 | 41.6 | 0.30 | 0.1717 | 0.6239 | 0.5541 |
| C18:1n7 | 4.31 | 4.28 | 4.11 | 3.98 | 4.12 | 3.37 | 0.10 | 0.1288 | 0.0442 | 0.4759 |
| C18:2n6 | 4.96 | 5.99 | 5.85 | 6.45 | 7.40 | 6.25 | 0.19 | 0.1068 | 0.0057 | 0.4285 |
| C20:0 | 0.15 | 0.15 | 0.15 | 0.16 | 0.14 | 0.15 | 0.002 | 0.0967 | 0.9927 | 0.5371 |
| C20:1n9 | 0.68 | 0.64 | 0.66 | 0.72 | 0.67 | 0.75 | 0.014 | 0.3346 | 0.1095 | 0.6467 |
| C18:3n3 | 0.20 | 0.23 | 0.23 | 0.25 | 0.26 | 0.24 | 0.011 | 0.7391 | 0.1252 | 0.7529 |
| C20:2n6 | 0.18 | 0.19 | 0.20 | 0.21 | 0.24 | 0.23 | 0.005 | 0.0957 | 0.0002 | 0.6850 |
| C20:3n6 | 0.13 | 0.16 | 0.15 | 0.15 | 0.19 | 0.14 | 0.007 | 0.0977 | 0.3758 | 0.5273 |
| C20:4n6 | 0.79 | 1.00 | 0.91 | 1.07 | 1.39 | 0.81 | 0.06 | 0.1017 | 0.1540 | 0.2925 |
| C20:5n3 | 0.039 | 0.056 | 0.043 | 0.053 | 0.085 | 0.074 | 0.005 | 0.1667 | 0.0245 | 0.7908 |
| C22:4n6 | 0.14 | 0.18 | 0.16 | 0.17 | 0.21 | 0.15 | 0.009 | 0.1267 | 0.2808 | 0.5435 |
| C22:5n3 | 0.10 | 0.12 | 0.11 | 0.13 | 0.17 | 0.10 | 0.006 | 0.0443 | 0.1048 | 0.2836 |
| C22:6n3 | 0.015 | 0.018 | 0.013 | 0.017ab | 0.025b | 0.012a | 0.0001 | 0.0116 | 0.2723 | 0.3117 |
| SFA 2 | 41.4 | 41.4 | 42.8 | 41.1 | 40.6 | 42.8 | 0.32 | 0.0659 | 0.5751 | 0.8756 |
| MUFA 3 | 52.0 | 50.6 | 49.5 | 50.3 | 49.4 | 49.2 | 0.38 | 0.1526 | 0.1677 | 0.7667 |
| PUFA 4 | 6.54 | 7.95 | 7.67 | 8.51 | 9.97 | 8.02 | 0.25 | 0.0617 | 0.0064 | 0.3169 |
| n6 | 6.19 | 7.52 | 7.27 | 8.06 | 9.43 | 7.59 | 0.24 | 0.0655 | 0.0069 | 0.3129 |
| n3 | 0.35 | 0.43 | 0.39 | 0.45a | 0.54b | 0.43a | 0.014 | 0.0410 | 0.0048 | 0.496 |
| PUFA/SFA | 0.16 | 0.19 | 0.18 | 0.21a | 0.25b | 0.19a | 0.006 | 0.0494 | 0.0064 | 0.3141 |
| MUFA/SFA | 1.27 | 1.22 | 1.16 | 1.23 | 1.22 | 1.15 | 0.02 | 0.1198 | 0.6920 | 0.9242 |
| Longissimus | Gluteus | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TN | TN-pf | HS | TN | TN-pf | HS | SEM 1 | Treatment (T) | Muscle (M) | T × M | |
| pH 30 min | 6.26 | 6.16 | 6.20 | 6.30 | 6.35 | 6.33 | 0.04 | 0.9780 | 0.167 | 0.7997 |
| pH 24 h | 5.57 | 5.56 | 5.65 | 5.55 | 5.51 | 5.64 | 0.03 | 0.2789 | 0.6580 | 0.9498 |
| Lightness L* | 37.9b | 36.6a | 36.5a | 34.1b | 31.7a | 32.5a | 0.27 | 0.0190 | 0.0001 | 0.6892 |
| Redness a* | 7.07 | 8.29 | 7.11 | 12.4 | 12.8 | 11.9 | 0.21 | 0.1352 | 0.0001 | 0.6820 |
| Yellowness b* | 3.59 | 3.70 | 3.60 | 6.47 | 6.20 | 6.30 | 0.13 | 0.9593 | 0.0001 | 0.8336 |
| Chroma C* | 7.97 | 9.17 | 7.82 | 14.1 | 14.4 | 13.5 | 0.23 | 0.1337 | 0.0001 | 0.7544 |
| Hue angle h° | 25.9 | 25.4 | 28.3 | 27.8 | 27.3 | 27.8 | 0.52 | 0.4130 | 0.2870 | 0.5510 |
| Longissimus | Gluteus | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TN | TN-pf | HS | TN | TN-pf | HS | SEM 1 | Treatment (T) | Muscle (M) | T × M | |
| MDA 2 (mg/kg) | 0.17 | 0.15 | 0.20 | 0.23 | 0.29 | 0.21 | 0.04 | 0.6722 | 0.0002 | 0.0205 |
| ABTS (mM/kg) | 2.96 | 2.51 | 2.64 | 3.07 | 3.77 | 3.45 | 0.07 | 0.7480 | 0.0001 | 0.0061 |
| DPPH (mM/kg) | 0.60a | 0.59a | 0.65b | 0.64 | 0.65 | 0.64 | 0.005 | 0.0452 | 0.0061 | 0.0408 |
| FRAP (mM/kg) | 0.42 | 0.38 | 0.40 | 0.51b | 0.41a | 0.45ab | 0.011 | 0.0304 | 0.0054 | 0.4064 |
| GPx 3 (U/g prot) | 0.24 | 0.20 | 0.22 | 0.14a | 0.13a | 0.21b | 0.008 | 0.0382 | 0.0007 | 0.0752 |
| CAT 4 (U/g prot) | 332ab | 238a | 438b | 180b | 115a | 214b | 16 | 0.0016 | 0.0001 | 0.4098 |
| SOD 5 (U/mg prot) | 1.42b | 1.27a | 1.46b | 1.42 | 1.34 | 1.41 | 0.02 | 0.0668 | 0.9140 | 0.6450 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pardo, Z.; Fernández-Fígares, I.; Lachica, M.; Lara, L.; Nieto, R.; Seiquer, I. Impact of Heat Stress on Meat Quality and Antioxidant Markers in Iberian Pigs. Antioxidants 2021, 10, 1911. https://doi.org/10.3390/antiox10121911
Pardo Z, Fernández-Fígares I, Lachica M, Lara L, Nieto R, Seiquer I. Impact of Heat Stress on Meat Quality and Antioxidant Markers in Iberian Pigs. Antioxidants. 2021; 10(12):1911. https://doi.org/10.3390/antiox10121911
Chicago/Turabian StylePardo, Zaira, Ignacio Fernández-Fígares, Manuel Lachica, Luis Lara, Rosa Nieto, and Isabel Seiquer. 2021. "Impact of Heat Stress on Meat Quality and Antioxidant Markers in Iberian Pigs" Antioxidants 10, no. 12: 1911. https://doi.org/10.3390/antiox10121911
APA StylePardo, Z., Fernández-Fígares, I., Lachica, M., Lara, L., Nieto, R., & Seiquer, I. (2021). Impact of Heat Stress on Meat Quality and Antioxidant Markers in Iberian Pigs. Antioxidants, 10(12), 1911. https://doi.org/10.3390/antiox10121911