Characterisation of Seasonal Mytilus edulis By-Products and Generation of Bioactive Hydrolysates



Abstract
Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sampling
2.3. Cooked Meat Yield
2.4. By-Product Mussel Hydrolysis with Protamex
2.5. Degree of Hydrolysis
2.6. Protein and Lipid Composition
2.7. Techno-Functional Properties of Hydrolysate
2.7.1. Emulsion Activity and Emulsion Stability Assay of Mussel Hydrolysates
2.7.2. Solubility of Protein Hydrolysates
2.7.3. Water Activity
2.8. Amino Acid Composition of Peptides
2.9. Sequencing Using Mass Spectrometry (MS) and In Silico Analysis of Peptides
2.10. Gas Chromatography Mass Spectrometry for Fatty Acid Composition Analysis
2.11. Bioassays
2.11.1. Angiotensin-Converting Enzyme (ACE-I) Inhibition
2.11.2. Acetylcholinesterase (EC 3.1.1.7) (AChE) Inhibition
2.11.3. Dipeptidyl Peptidase IV (DPP-IV) Inhibition
2.12. In-Silico Analysis and Statistics
3. Results and Discussion
3.1. Sampling
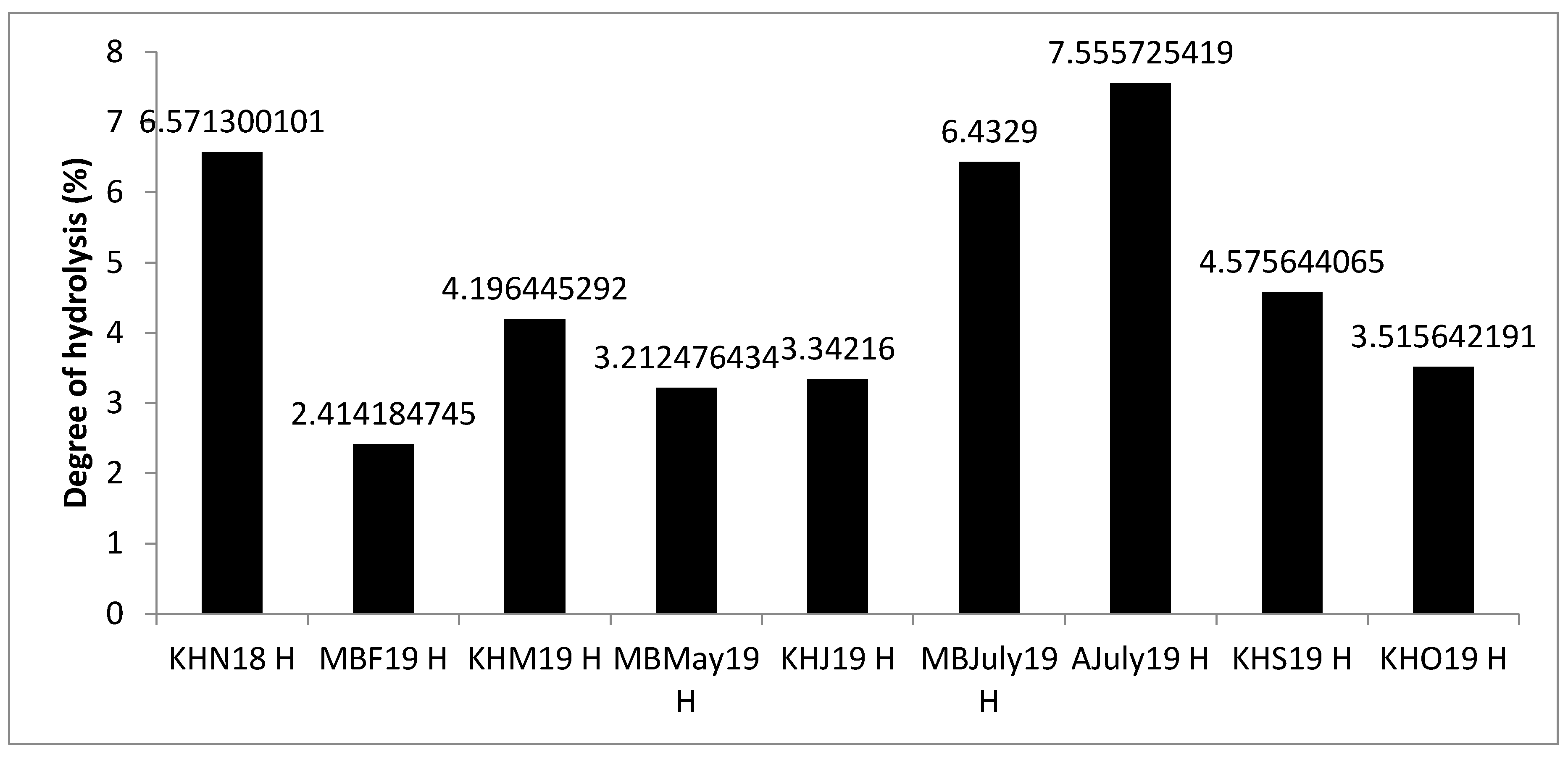
3.2. Degree of Hydrolysis and Peptide Length
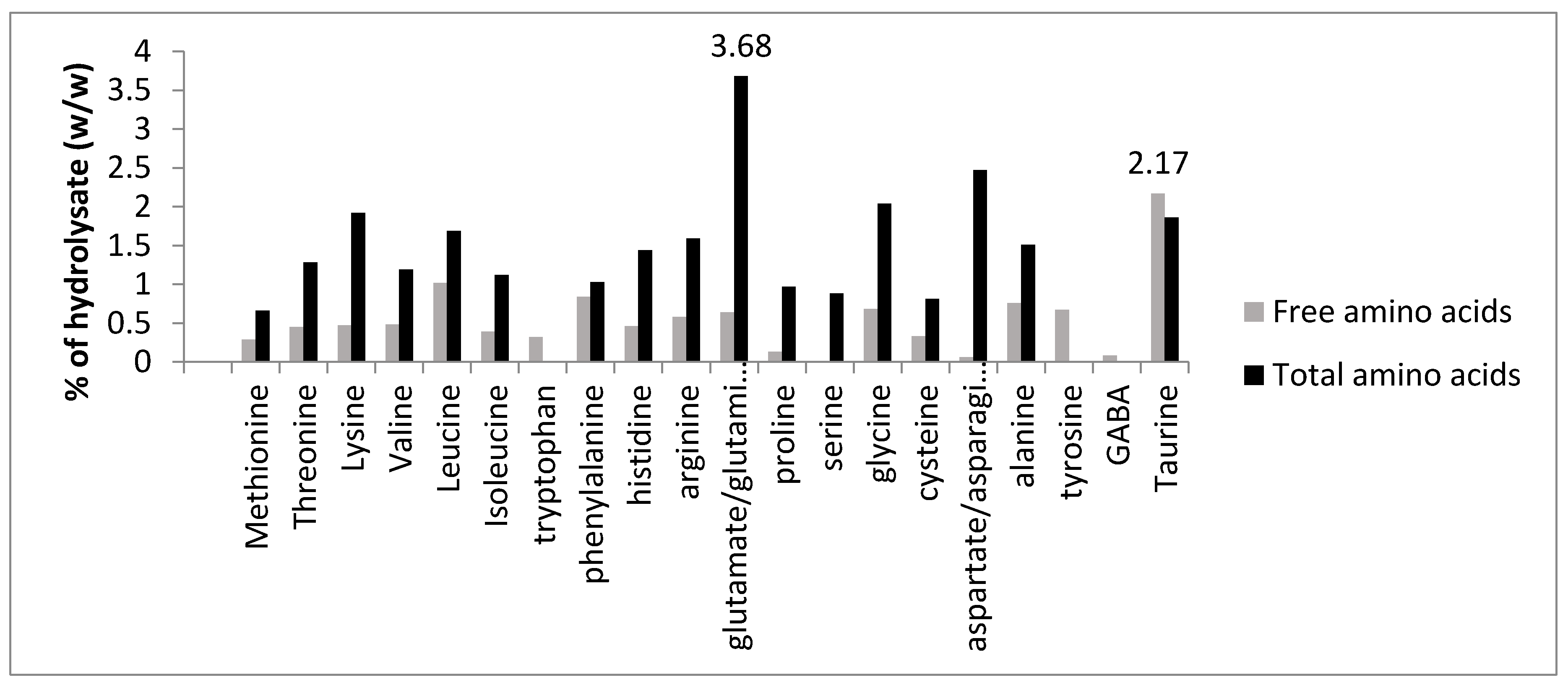
3.3. Free Amino Acid Content
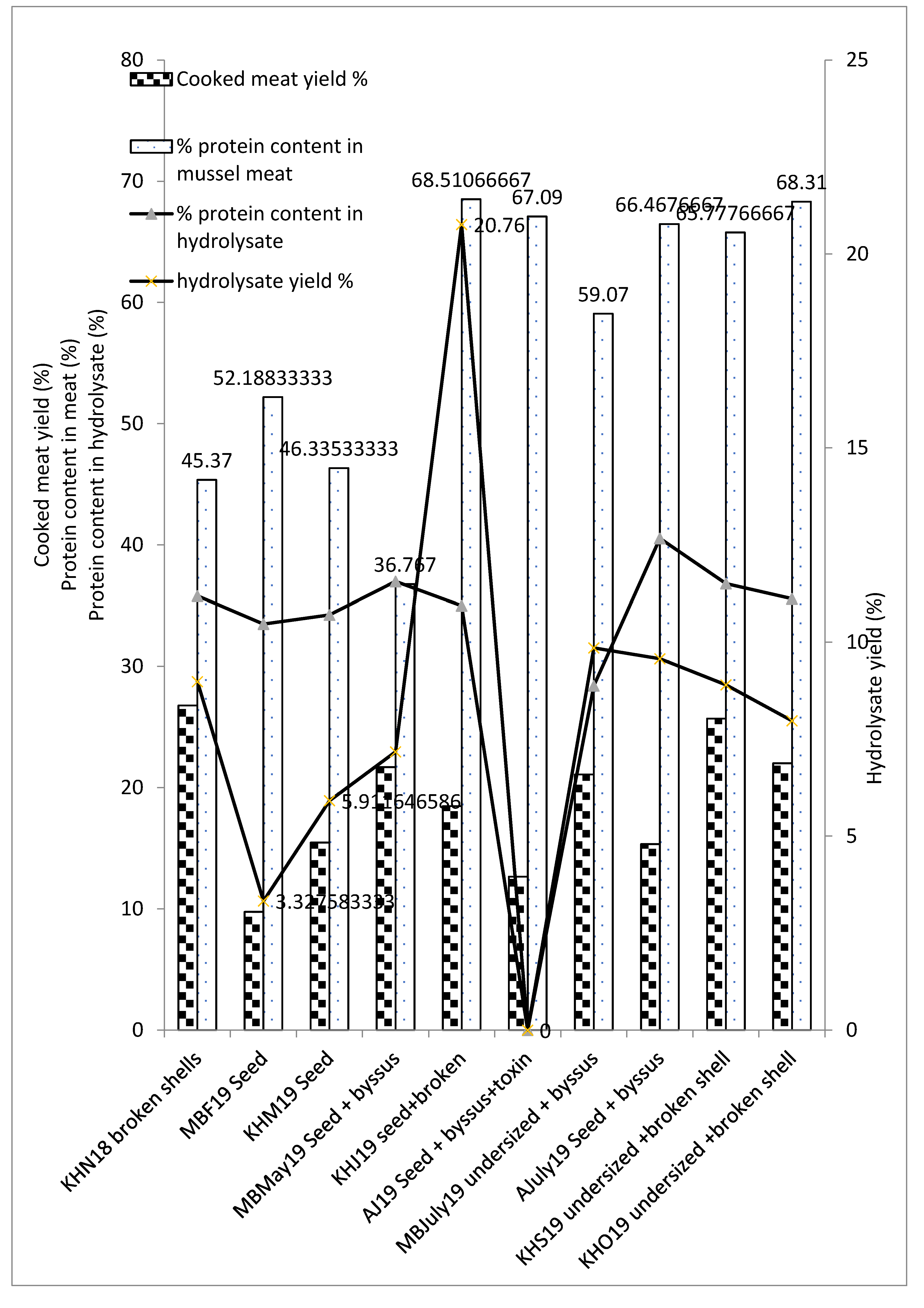
3.4. Cooked Meat Yield, Hydrolysate Yield and Protein Content across Different Seasons and By-Product Types
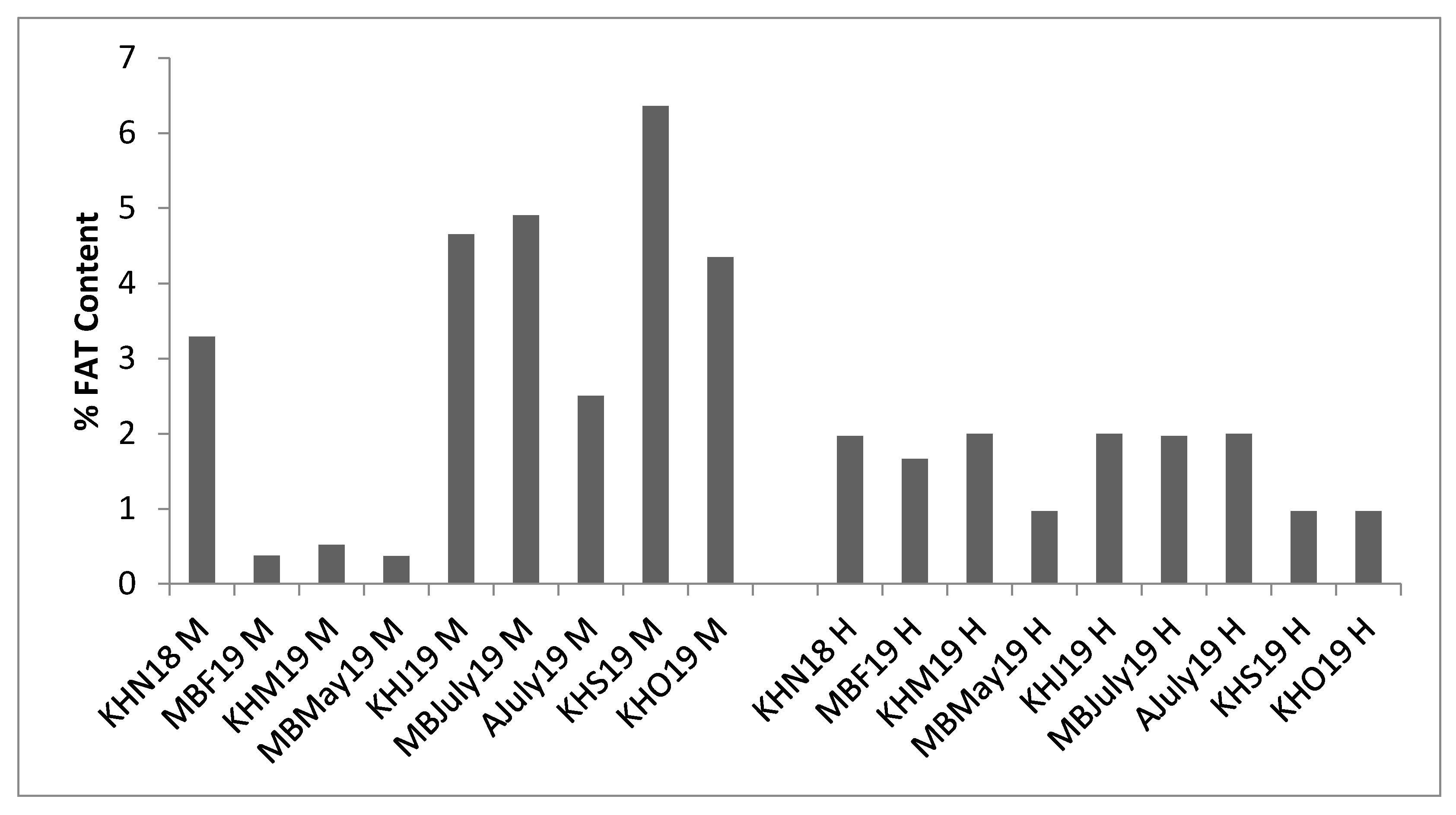
3.5. Lipid Content of Mussels and Hydrolysates
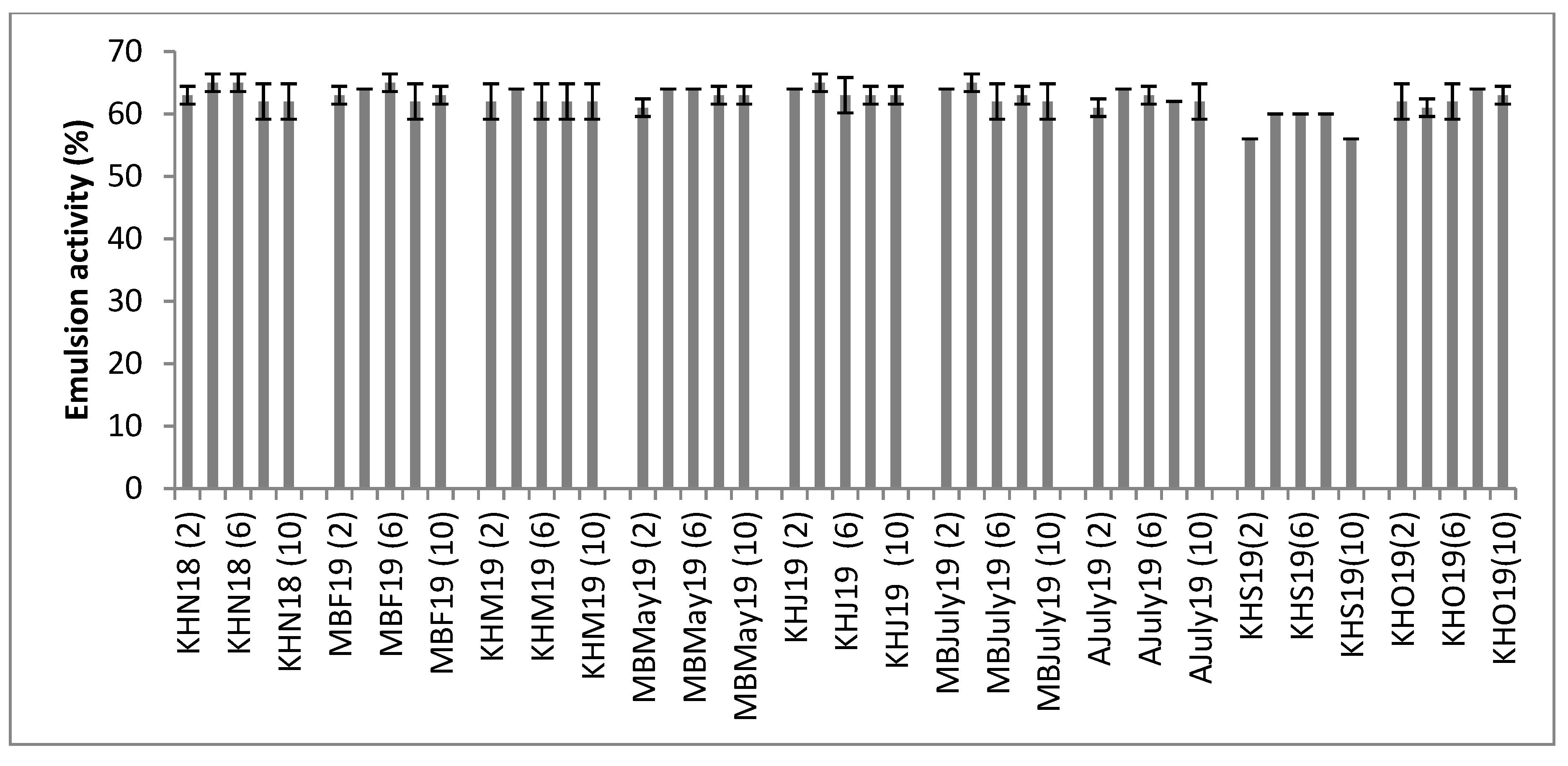
3.6. Emulsion Activity and Emulsion Stability
3.7. Water Activity
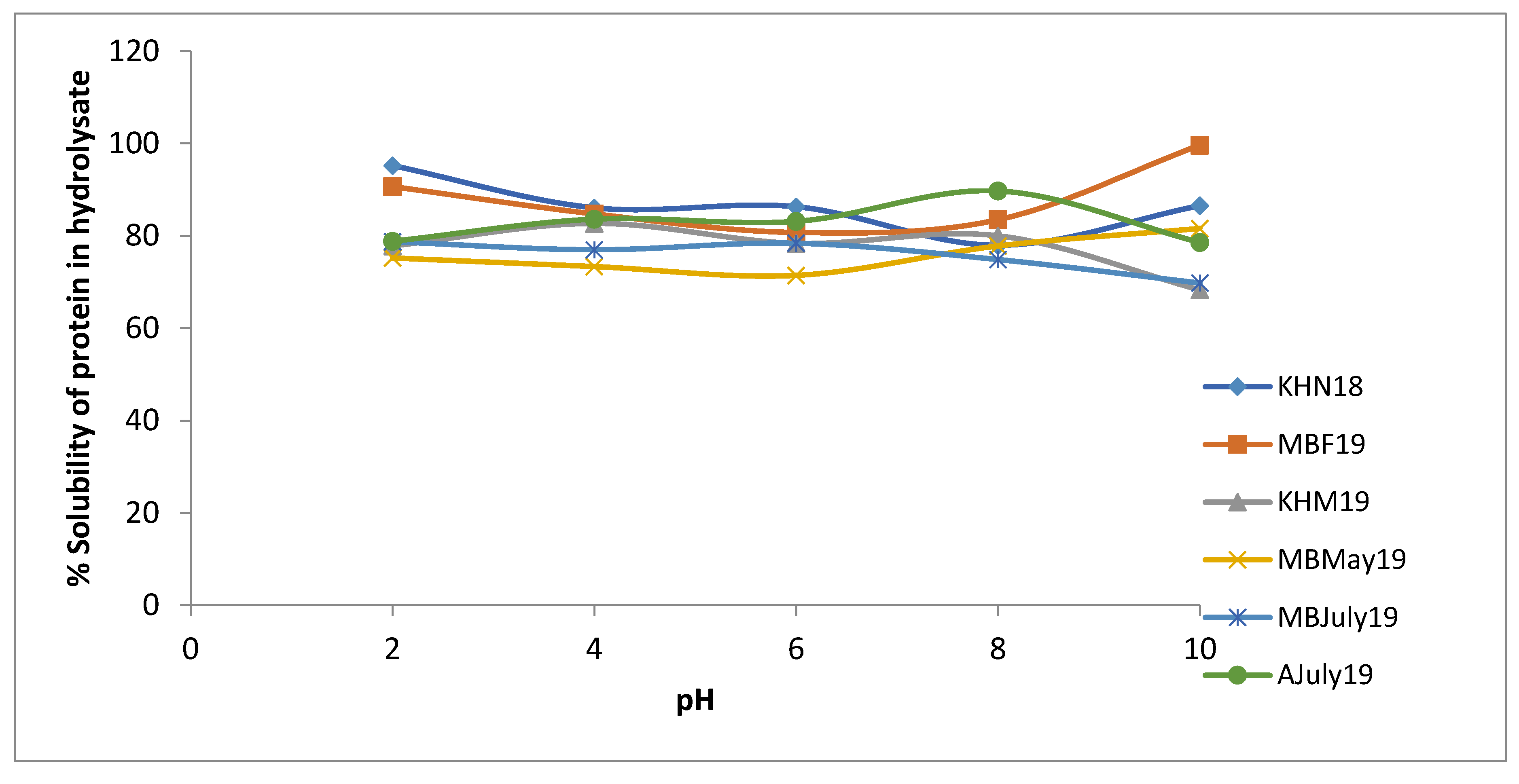
3.8. Solubility of Mussel By-Product Hydrolysates
3.9. Amino Acid Composition of Hydrolysates
3.10. Fatty Acid Analysis of Hydrolysates
3.11. Sequence, Physicochemical Nature and Bioactivity of Identified Peptides
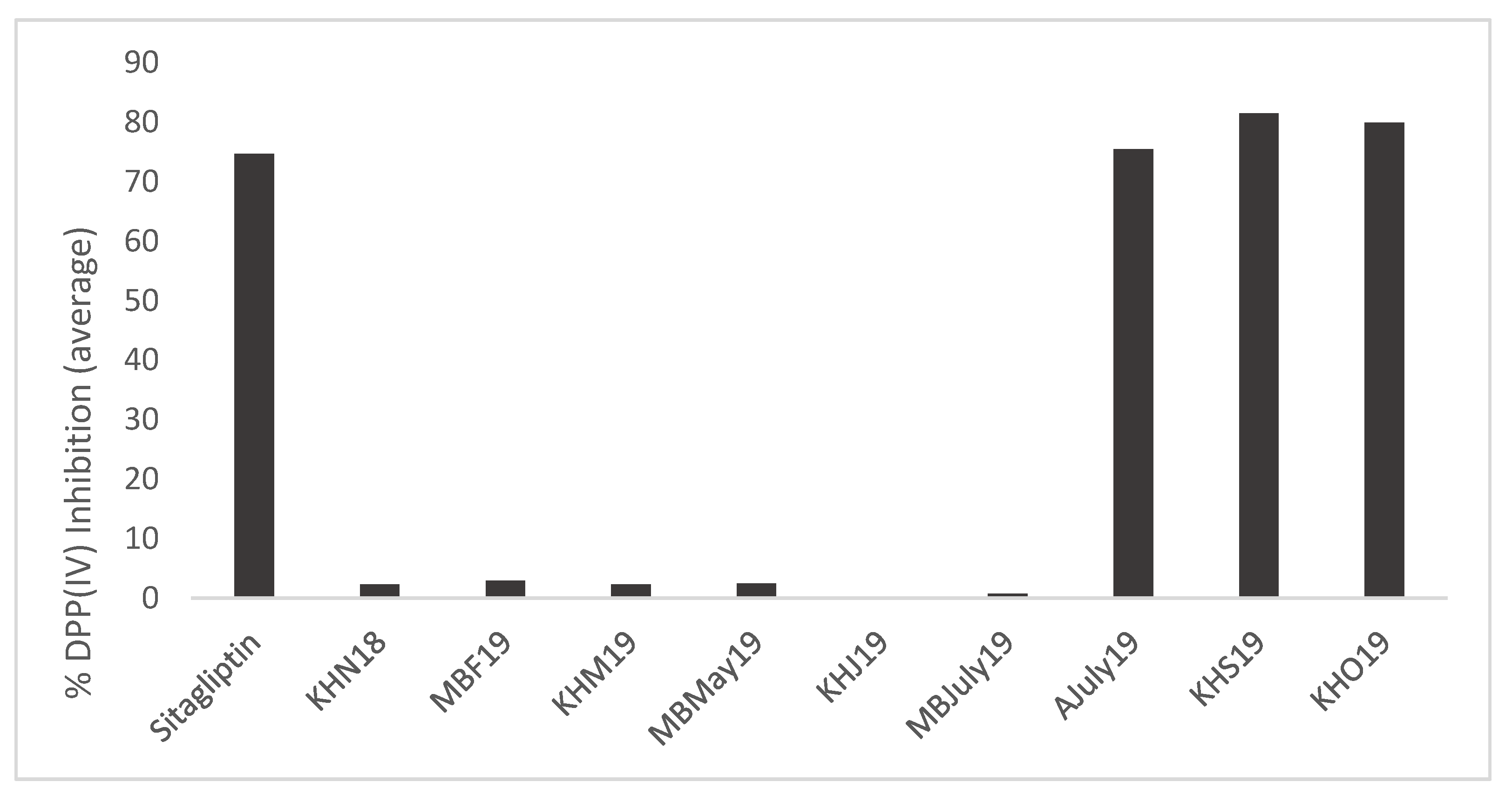
- DPP (IV) inhibition: The DPP-IV inhibitory activity at 30 min for the generated mussel by-product hydrolysates ranged from 0% to 90.51% ± 0.18%, whereas it was 81.14% ± 0.47% for sitagliptin when samples were assayed at a concentration of 1 mg/mL compared to the positive control, sitagliptin, which was assayed at a concentration of 18 nM (IC50 value) (Figure 7).
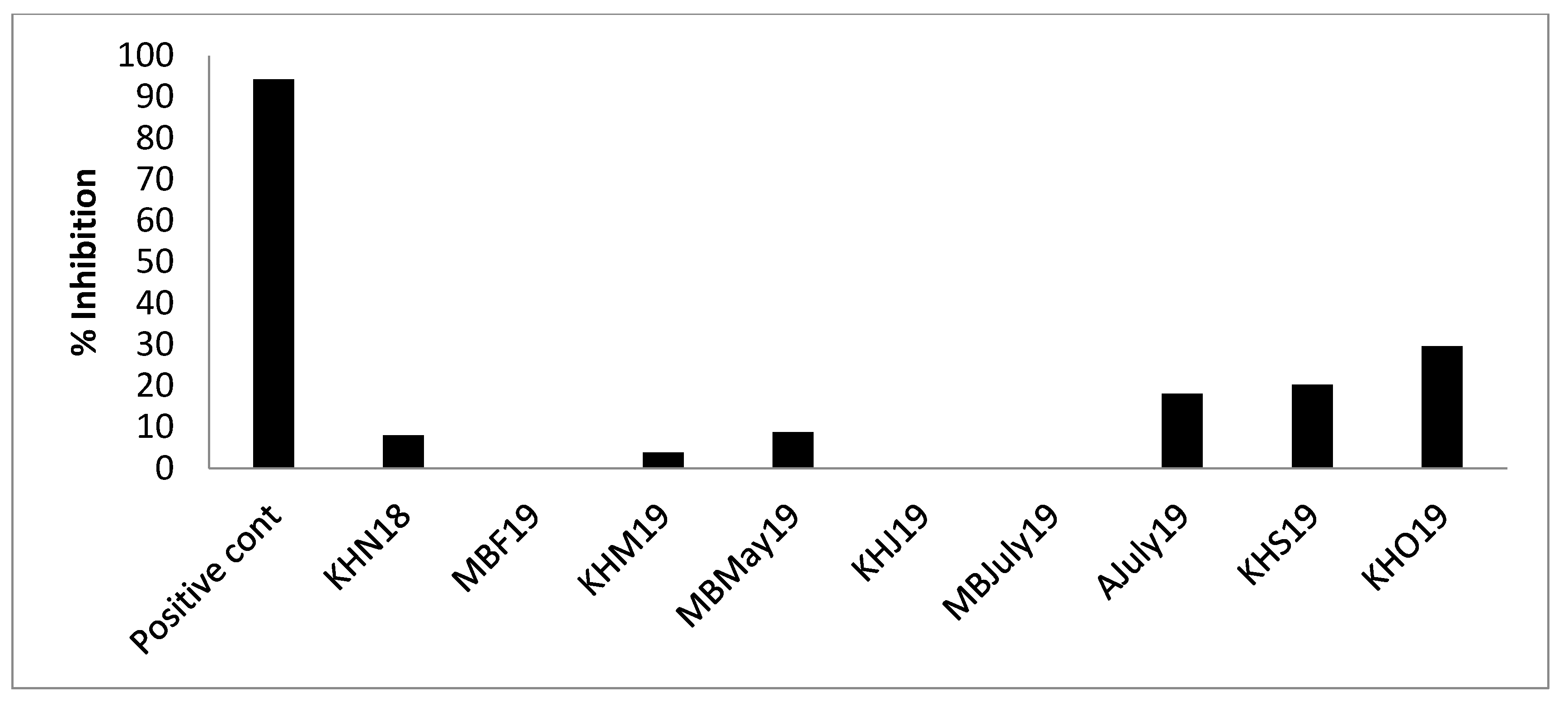
- AChE inhibition: A trend similar to DPPIV inhibition was seen. AJuly19, KHS19 and KHO19 displayed AChE inhibition (Figure 8), though the % inhibition (KHO19: 29.59% ± 1.88%) of the samples (1 mg/mL) was low as compared to positive control acotiamide dihydrochloride (94.201% ± 0.89%) assayed at 100 uM concentration (IC50 3 uM). Researchers have found AChE, ACE-I and DPPH inhibitory activity in subcritical extracts of blue mussels [47]. The obtained results warrant further studies to explore the possible potential of some of these hydrolysates in promoting mental health.
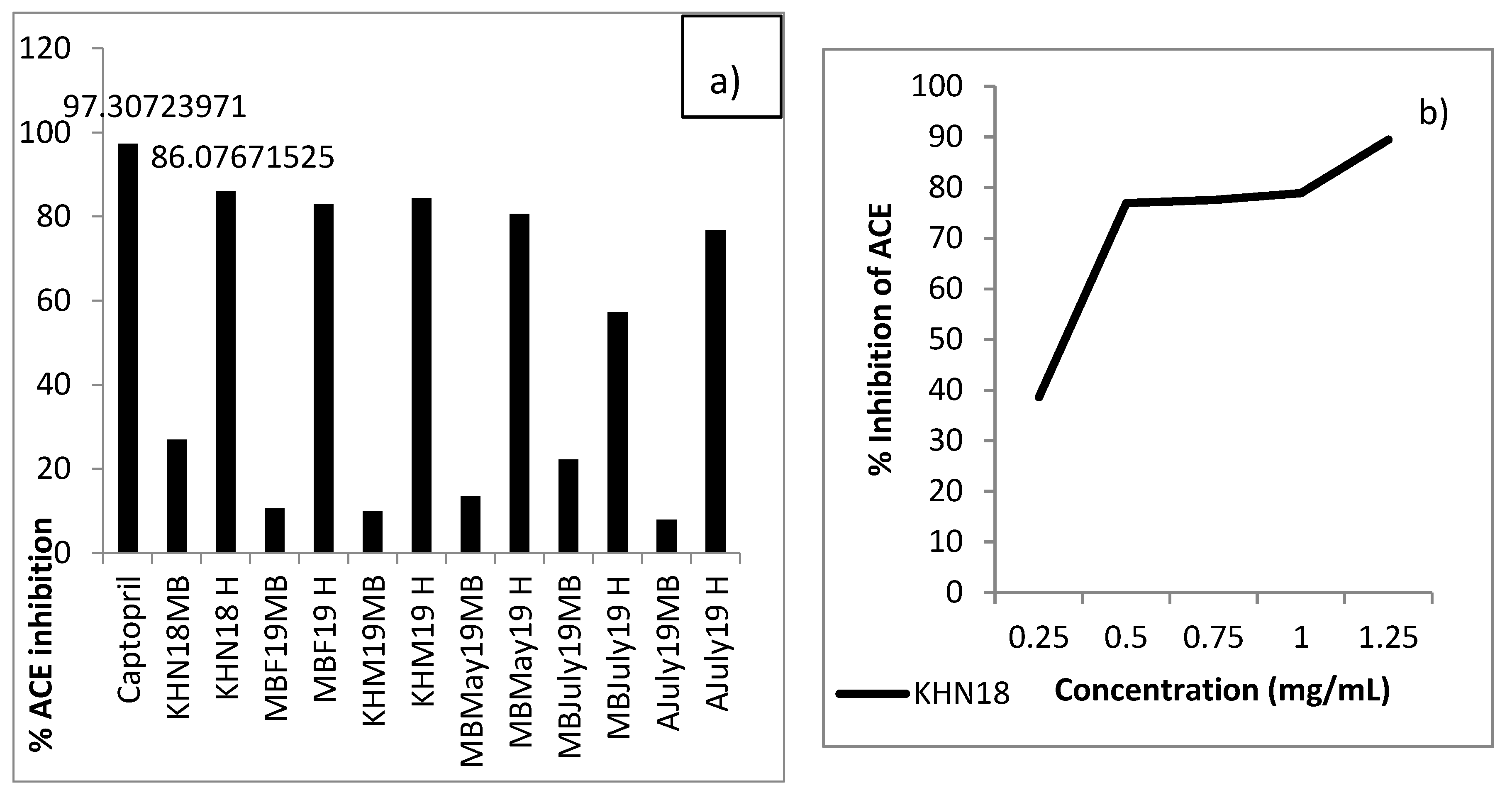
- ACE-I inhibition: All the samples showed appreciable ACE-I inhibition as compared to un-hydrolysed mussel broth, as seen in Figure 9. The percentage of ACE-I inhibition was seen to vary between 22.23% ± 1.79% and 86.08% ± 1.59% for hydrolysates as compared to 97.31% ± 0.255% by positive control captopril. The KHN18 sample inhibited ACE-I by 86.08% ± 1.59% when assayed at a concentration of 1 mg/mL, compared to captopril which was assayed at a concentration of 0.5 mg/mL. Further, the IC50 value for KHN18 was 0.2944 mg/mL, which is lower than reported values of other marine hydrolysates (1.50–2.54 mg/mL) [48], highlighting the superior ACE-I inhibitor potential of the mussel hydrolysates. An interesting study conducted on Mytilus edulis-based fermented sauce also established the formation of ACE-I inhibitory peptides from blue mussel sources [14]. The findings from the in vitro assay also confirm the results obtained from bioinformatics mining of the sequenced peptides, as 92% of the peptides identified in the 3-kDa permeate fraction and assessed using BIOPEP had ACE-I inhibitory activity.
4. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Fishery and Aquaculture Statistics; FAO: Rome, Italy, 2016. [Google Scholar]
- Naik, A.S.; Hayes, M. Bioprocessing of mussel by-products for value added ingredients. Trends Food Sci. Technol. 2019, 92, 111–121. [Google Scholar] [CrossRef]
- Laura, E.; Petes, B.A.M.; Alyssa, L. Harris Intertidal mussels exhibit energetic trade-offs between reproduction and stress resistance. Ecol. Monogr. 2008, 78, 387–402. [Google Scholar]
- Fernandez, A.; Grienke, U.; Soler-Vila, A.; Guiheneuf, F.; Stengel, D.B.; Tasdemir, D. Seasonal and geographical variations in the biochemical composition of the blue mussel (Mytilus edulis L.) from Ireland. Food Chem. 2015, 177, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Bongiorno, T.; Iacumin, L.; Tubaro, F.; Marcuzzo, E.; Sensidoni, A.; Tulli, F. Seasonal changes in technological and nutritional quality of Mytilus galloprovincialis from suspended culture in the Gulf of Trieste (North Adriatic Sea). Food Chem. 2015, 173, 355–362. [Google Scholar] [CrossRef]
- Dare, P.J.; Edwards, D.B. Seasonal changes in flesh weight and biochemical composition of mussels (Mytilus Edulis, L.) in the Conwy Estuary, North Wales. J. Exp. Mar. Biol. Ecol. 1975, 18, 89–97. [Google Scholar] [CrossRef]
- Carboni, S.; Kaur, G.; Pryce, A.; McKee, K.; Desbois, A.P. Mussel Consumption as a “Food First” Approach to Improve Omega-Status. Nutrients 2019, 11, 1381. [Google Scholar] [CrossRef]
- Doggrell, S.A. Lyprinol—Is it a useful anti-inflammatory agent? Evid. Based Complementary Altern. Med. Ecam 2011, 2011, 307121. [Google Scholar] [CrossRef]
- Wang, B.; Li, L.; Chi, C.F.; Ma, J.H.; Luo, H.Y.; Xu, Y.F. Purification and characterisation of a novel antioxidant peptide derived from blue mussel (Mytilus edulis) protein hydrolysate. Food Chem. 2013, 138, 1713–1719. [Google Scholar] [CrossRef]
- Ilva, V.M.; Park, K.J.; Hubinger, M.D. Optimization of the enzymatic hydrolysis of mussel meat. J. Food Sci. 2010, 75, C36–C42. [Google Scholar]
- Dai, Z.-Y.; Zhang, Y.-P.; Zhang, H.; Lu, Y.-B. Preparation and Characterization of Mussel (Mytilus Edulis) Protein Hydrolysates with Angiotensin-I-Converting Enzyme (Ace) Inhibitory Activity by Enzymatic Hydrolysis. J. Food Biochem. 2012, 36, 66–74. [Google Scholar] [CrossRef]
- Nguyen HT, M.; Sylla KS, B.; Randriamahatody, Z.; Donnay-Moreno, C.; Moreau, J.; Tran, L.T.; Bergé, J. Enzymatic Hydrolysis of Yellowfin Tuna (Thunnus albacares)By-Products Using Protamex Protease. Food Technol. Biotechnol. 2011, 49, 48–55. [Google Scholar]
- Park, S.Y.; Ahn, C.-B.; Je, J.-Y. Antioxidant and Anti-Inflammatory Activities of Protein Hydrolysates fromMytilus Edulisand Ultrafiltration Membrane Fractions. J. Food Biochem. 2014, 38, 460–468. [Google Scholar] [CrossRef]
- Rajapakse, N.; Mendis, E.; Jung, W.-K.; Je, J.-Y.; Kim, S.-K. Purification of a radical scavenging peptide from fermented mussel sauce and its antioxidant properties. Food Res. Int. 2005, 38, 175–182. [Google Scholar] [CrossRef]
- Beaulieu, L.; Thibodeau, J.; Bonnet, C.; Bryl, P.; Carbonneau, M.E. Evidence of anti-proliferative activities in blue mussel (Mytilus edulis) by-products. Mar. Drugs 2013, 11, 975–990. [Google Scholar] [CrossRef]
- Oh, R.; Lee, M.J.; Kim, Y.O.; Nam, B.H.; Kong, H.J.; Kim, J.W.; Park, J.Y.; Seo, J.K.; Kim, D.G. Purification and characterization of an antimicrobial peptide mytichitin-chitin binding domain from the hard-shelled mussel, Mytilus coruscus. Fish Shellfish Immunol. 2018, 83, 425–435. [Google Scholar] [CrossRef]
- Jung, W.-K.; Kim, S.-K. Isolation and characterisation of an anticoagulant oligopeptide from blue mussel, Mytilus edulis. Food Chem. 2009, 117, 687–692. [Google Scholar] [CrossRef]
- Elavarasan, K.; Shamasundar, B.A.; Badii, F.; Howell, N. Angiotensin I-converting enzyme (ACE) inhibitory activity and structural properties of oven- and freeze-dried protein hydrolysate from fresh water fish (Cirrhinus mrigala). Food Chem. 2016, 206, 210–216. [Google Scholar] [CrossRef]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Karami, Z.; Akbari-Adergani, B. Bioactive food derived peptides: A review on correlation between structure of bioactive peptides and their functional properties. J. Food Sci. Technol. 2019, 56, 535–547. [Google Scholar] [CrossRef]
- Ovando, C.A.; Carvalho, J.C.D.; Vinícius de Melo Pereira, G.; Jacques, P.; Soccol, V.T.; Soccol, C.R. Functional properties and health benefits of bioactive peptides derived from Spirulina: A review. Food Rev. Int. 2018, 34, 34–51. [Google Scholar] [CrossRef]
- Turck, D.; Bresson, J.L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Safety of shrimp peptide concentrate as a novel food pursuant to Regulation (EU) 2015. EFSA J. 2018, 16. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Enzymic Hydrolysis of Food Proteins; Elsevier Applied Science Publishers: Barking, UK, 1986; p. xxiv. 427p. [Google Scholar]
- Association of Official Analytical Chemists. Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 1998. [Google Scholar]
- Fountoulakis, M.; Lahm, H.W. Hydrolysis and amino acid composition of proteins. J. Chromatogr. A 1998, 826, 109–134. [Google Scholar] [CrossRef]
- The UniProt Consortium UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [CrossRef] [PubMed]
- Perkins, D.N.; Pappin, D.J.; Creasy, D.M.; Cottrell, J.S. Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophoresis 1999, 20, 3551–3567. [Google Scholar] [CrossRef]
- Keska, P.; Rohn, S.; Halagarda, M.; Wojciak, K.M. Different carcass elements of organic and conventional pork –potential sources of antioxidant activities. Antioxidants 2020, 9, 835. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef]
- Araujo, P.; Nguyen, T.T.; Frøyland, L.; Wang, J.; Kang, J.X. Evaluation of a rapid method for the quantitative analysis of fatty acids in various matrices. J. Chromatogr. 2008, 1212, 106–113. [Google Scholar] [CrossRef]
- Silvestre, M.P.C.; Morais, H.A.; Silva, V.D.M.; Silva, M.R. Degree of hydrolysis and peptide profile of whey proteins using pancreatin. Nutrire 2013, 38, 278–290. [Google Scholar] [CrossRef]
- Chiang, J.H.; Loveday, S.M.; Hardacre, A.K.; Parker, M.E. Effects of enzymatic hydrolysis treatments on the physicochemical properties of beef bone extract using endo- and exoproteases. Int. J. Food Sci. Technol. 2019, 54, 111–120. [Google Scholar] [CrossRef]
- Novikov, V.Y.; Derkach, S.R.; Kuchina, Y.A.; Shironina, A.Y.; Mukhin, V.A. Kinetics of enzymatic reactions in the production of fish protein hydrolysates. J. Dispers. Sci. Technol. 2018, 39, 1454–1461. [Google Scholar] [CrossRef]
- Duinker, A.; Håland, L.; Hovgaard, P.; Mortensen, S. Gonad development and spawning in one and two year old mussels (Mytilus edulis) from Western Norway. J. Mar. Biol. Assoc. UK 2008, 88, 1465–1473. [Google Scholar] [CrossRef]
- Naqash, S.Y.; Nazeer, R.A. Antioxidant and functional properties of protein hydrolysates from pink perch (Nemipterus japonicus) muscle. J. Food Sci. Technol. 2013, 50, 972–978. [Google Scholar] [CrossRef] [PubMed]
- Segura-Campos, M.R.; Espinosa-García, L.; chel-Guerrero, L.A.; Betanar-Ancona, D.A. Effect of enzymatic hydrolysis on solubility, hydrophobicity and in vivo digestibility in cowpea (Vigna unguiculata). Int. J. Food Prop. 2012, 15, 770–780. [Google Scholar] [CrossRef]
- Yu, D.; Sun, Y.; Wang, W.; O’Keefe, S.F.; Neilson, A.P.; Feng, H.; Wang, Z.; Huang, H. Recovery of protein hydrolysates from brewer’s spent grain using enzyme and ultrasonication. Int. J. Food Sci. Technol. 2020, 55, 357–368. [Google Scholar] [CrossRef]
- Ben Slama-Ben Salem, R.; Bkhairia, I.; Abdelhedi, O.; Nasri, M. Octopus vulgaris protein hydrolysates: Characterization, antioxidant and functional properties. J. Food Sci. Technol. 2017, 54, 1442–1454. [Google Scholar] [CrossRef]
- Gorissen, S.H.M.; Crombag, J.J.R.; Senden, J.M.G. Protein content and amino acid composition of commercially available plant-based protein isolates. Amino Acids 2018, 50, 1685–1695. [Google Scholar] [CrossRef]
- Bak, K.H.; Petersen, M.A.; Lametsch, R.; Hansen, E.T.; Ruiz-Carrascal, J. Development of Volatile Compounds during Hydrolysis of Porcine Hemoglobin with Papain. Molecules 2018, 23, 357. [Google Scholar] [CrossRef]
- Breternitz, N.R.; Bolini, H.M.A.; Hubinger, M.D. Sensory acceptance evaluation of a new food flavoring produced by microencapsulation of a mussel (Perna perna) protein hydrolysate. LWT Food Sci. Technol. 2017, 83, 141–149. [Google Scholar] [CrossRef]
- Bachmanov, A.A.; Bosak, N.P.; Glendinning, J.I.; Inoue, M.; Li, X.; Manita, S.; McCaughey, S.A.; Murata, Y.; Reed, D.R.; Tordoff, M.G.; et al. Genetics of Amino Acid Taste and Appetite. Adv. Nutr. 2016, 7, 806S–822S. [Google Scholar] [CrossRef]
- Xu, Y.J.; Arneja, A.S.; Tappia, P.S.; Dhalla, N.S. The potential health benefits of taurine in cardiovascular disease. Exp. Clin. Cardiol. 2008, 13, 57–65. [Google Scholar]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Wolfenden, R.; Andersson, L.; Cullis, P.; Southgate, C. Affinities of Amino Acid Side Chains for Solvent Water. Biochemistry 1981, 20, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Mora, L.; Gallego, M.; Aristoy, M.C.; Reig, M.; Toldrá, F. Bioactive Peptides, chapter 12. In Innovative Thermal and Non-Thermal Processing, Bioaccessibility and Bioavailability of Nutrients and Bioactive Compounds; Woodhead Publishing: Sawston, UK, 2019; pp. 333–345. [Google Scholar]
- Han, J.-K.; Sung, S.-C.; Jo, M.-J.; Lee, S.-C. Antioxidant, ACE inhibitory, and acetylcholinesterase inhibitory activities of subcritical water extract of blue mussel. Food Sci. Biotechnol. 2018, 27, 847–851. [Google Scholar] [CrossRef] [PubMed]
- Ghanbari, R.; Zarei, M.; Ebrahimpour, A.; Abdul-Hamid, A.; Ismail, A.; Saari, N. Angiotensin-I Converting Enzyme (ACE) Inhibitory and Anti-Oxidant Activities of Sea Cucumber (Actinopyga lecanora) Hydrolysates. Int. J. Mol. Sci. 2015, 16, 28870–28885. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Average aw | Standard Deviation (±) |
|---|---|---|
| KHN18 | 0.35 | 0.0028 |
| MBF19 | 0.44 | 0.0035 |
| KHM19 | 0.33 | 0.004 |
| MBMay19 | 0.33 | 0.0005 |
| KHJ19 | 0.35 | 0.0028 |
| MBJuly19 | 0.37 | 0.008 |
| AJuly19 | 0.30 | 0.0005 |
| KHS19 | 0.25 | 0.0047 |
| KHO19 | 0.29 | 0.0115 |
| Amino Acids | KHN18pro | MBF19 H | KHM19 H | MBMay19H | Soy Protein | Whey Protein | Icelandic Scallop Hydrolysate | Atlantic Salmon Hydrolysate | |
|---|---|---|---|---|---|---|---|---|---|
| Essential amino acids | Methionine | 0.6607 | 0.1135 | 0.1799 | 0.3687 | 0.3 | 1.8 | 0.457 | 1.3 |
| Threonine | 1.2815 | 0.2494 | 0.2855 | 0.2355 | 2.3 | 5.4 | 0.856 | 2.9 | |
| Lysine | 1.9237 | 0.2726 | 0.7896 | 0.4005 | 3.4 | 7.1 | 3.778 | 0 | |
| Valine | 1.1967 | 0.0063 | 0.4275 | 0.7775 | 2.2 | 3.5 | 1.681 | 2.5 | |
| Leucine | 1.6869 | 0.2679 | 0.2968 | 0.8958 | 5 | 8.6 | 0 | 2.4 | |
| Isoleucine | 1.1182 | 0.1956 | 0.2297 | 0.6471 | 1.9 | 3.8 | 0 | 1.2 | |
| tryptophan | 0 | 0 | 0 | 0.0989 | 0 | 0 | 0.812 | 0 | |
| phenylalanine | 1.0326 | 0.2275 | 0.347 | 0.565 | 3.2 | 2.5 | 2.811 | 3.1 | |
| histidine | 1.4382 | 0.3174 | 0.5225 | 0.5066 | 1.5 | 1.4 | 0.229 | 16.4 | |
| arginine | 1.5961 | 0.1854 | 0.2495 | 0.1888 | 4.8 | 1.7 | 4.611 | 1.3 | |
| Non-essential amino acids | glutamate/glutamine | 3.6814 | 0.5535 | 1.5894 | 1.9325 | 12.4 | 15.5 | 2.678 | 3.1 |
| proline | 0.9729 | 0.1508 | 0.2229 | 0.5169 | 3.3 | 4.8 | 0.741 | 0 | |
| serine | 0.8765 | 0.2015 | 0.2646 | 0.2063 | 3.4 | 4 | 0.941 | 4.8 | |
| glycine | 2.0381 | 0.2704 | 0.5315 | 1.5489 | 2.7 | 1.5 | 1.6 | 13.7 | |
| cysteine | 0.8134 | 0.0385 | 0.643 | 0.5137 | 0.2 | 0.8 | 0 | 0 | |
| aspartate/asparagine | 2.4696 | 0.6104 | 0.7903 | 0.66 | 0 | 0 | 4.116 | 0.4 | |
| alanine | 1.5091 | 0.2281 | 0.7916 | 0.8975 | 2.8 | 4.2 | 2.03 | 24.7 | |
| tyrosine | 0 | 0.0124 | 0.0078 | 0.021 | 2.2 | 2.4 | 1.909 | 1.9 | |
| GABA | 0.0065 | 0.0243 | 0.0507 | 0 | 0 | 0 | 0 | 0 | |
| Taurine | 1.8639 | 2.3182 | 2.5595 | 2.3964 | 0 | 0 | 0 | 0 | |
| Total | 26.166 | 6.2437 | 10.7793 | 13.3776 | 0 | 0 | 0 | 0 | |
| TEAA | 11.9346 | 1.8356 | 3.328 | 4.6844 | 24.6 | 35.8 | 15.235 | 31.1 | |
| TNEAA | 12.361 | 2.0656 | 4.8411 | 6.2968 | 27 | 33.2 | 14.015 | 48.6 | |
| Ratio of E:NE | 0.9655044 | 0.8886522 | 0.68744707 | 0.743933427 | 0.911111 | 1.078313 | 1.08704959 | 0.639917695 |
| mg Fatty Acid/g Lipid | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Name (Methyl Ester) | KHN18 | MBF19 | KHM19 | MBMay19 | KHJ19 | MBJuly19 | AJuly19 | KHS19 | KHO19 | |
| Butyric acid | C4:0 | 15.06 | 14.02 | 14.83 | 14.13 | 15.33 | 39.19 | 12.378 | 14.065 | 20.340 |
| Caproic acid | C6:0 | 16.26 | 15.10 | 15.98 | 15.20 | 15.45 | 18.05 | 13.369 | 15.130 | 21.986 |
| Caprylic acid | C8:0 | 30.58 | 39.55 | 34.01 | 30.57 | 28.85 | 35.12 | 19.442 | 48.533 | 61.002 |
| Capric acid | C10:0 | 22.59 | 31.65 | 27.53 | 24.54 | 22.93 | 25.86 | 15.243 | 38.067 | 44.464 |
| Undecanoic acid | C11:0 | 6.98 | 6.49 | 6.85 | 6.53 | 6.63 | 7.71 | 5.731 | 6.514 | 9.458 |
| Lauric acid | C12:0 | 9.26 | 8.16 | 8.66 | 8.42 | 8.86 | 8.91 | 7.886 | 8.605 | 12.316 |
| Tridecanoic acid | C13:0 | 7.13 | 6.54 | 6.93 | 6.64 | 6.75 | 7.85 | 5.886 | 6.613 | 9.576 |
| Myristic acid | C14:0 | 179.19 | 9.01 | 12.39 | 25.42 | 31.37 | 19.00 | 41.669 | 27.279 | 25.037 |
| Myristoleic acid | C14:1 | 5.58 | 3.00 | 3.17 | 3.03 | 3.25 | 3.73 | 2.914 | 3.066 | 4.441 |
| Pentadecanoic acid | C15:0 | 10.66 | 7.18 | 7.99 | 7.82 | 8.95 | 8.63 | 7.825 | 8.662 | 11.53 |
| cis-10-Pentadecenoic acid | C15:1 | 3.15 | 3.10 | 3.28 | 3.07 | 3.16 | 3.62 | 2.676 | 2.932 | 4.249 |
| Palmitic acid | C16:0 | 395.73 | 32.17 | 66.60 | 115.13 | 175.60 | 99.48 | 173.312 | 132.21 | 114.37 |
| Palmitoleic acid | C16:1 | 210.43 | 3.61 | 16.28 | 56.53 | 70.06 | 53.58 | 92.557 | 55.937 | 43.761 |
| cis-10-Heptadecenoic acid | C17:1 | 2.11 | 0.56 | 1.40 | 2.10 | 2.50 | 1.30 | 1.892 | 1.878 | 1.521 |
| Stearic acid | C18:0 | 34.27 | 20.56 | 22.26 | 23.38 | 39.42 | 28.38 | 30.469 | 29.365 | 34.148 |
| Elaidic acid | C18:1 t | 45.37 | 0.92 | 2.64 | 4.55 | 3.73 | 2.36 | 5.749 | 2.964 | 2.751 |
| Oleic acid | C18:1 c | 90.50 | 1.81 | 5.25 | 9.07 | 7.42 | 4.69 | 11.455 | 5.896 | 5.468 |
| Linolelaidic acid | C18:2 t | 4.74 | 3.50 | 3.95 | 3.95 | 2.64 | 4.11 | 4.353 | 3.489 | 5.418 |
| Linoleic acid | C18:2 c | 95.81 | 5.56 | 10.18 | 12.73 | 14.12 | 8.66 | 16.439 | 13.031 | 13.276 |
| γ-Linolenic acid | C18:3 n6 | 5.24 | 3.61 | 3.96 | 4.22 | 4.13 | 4.64 | 4.714 | 4.199 | 5.533 |
| α-Linolenic acid | C18:3 n3 | 43.98 | 5.99 | 12.26 | 19.32 | 23.31 | 9.61 | 22.633 | 19.865 | 20.183 |
| Arachidic acid | C20:0 | 16.47 | 14.80 | 15.62 | 14.94 | 15.24 | 17.83 | 13.191 | 14.866 | 21.402 |
| cis-11-Eicosenoic acid | C20:1 | 18.18 | 10.10 | 9.75 | 6.08 | 15.54 | 6.77 | 11.152 | 7.211 | 8.213 |
| cis-11,14-Eicosadienoic acid | C20:2 | 9.22 | 1.42 | 4.03 | 3.43 | 5.01 | 1.74 | 4.835 | 4.634 | 4.964 |
| Henicosanoic acid | C21:0 | 9.26 | 8.08 | 8.54 | 7.98 | 8.18 | 9.44 | 7.213 | 8.026 | 11.619 |
| cis-8,11,14-Eicosatrienoic acid | C20:3 n6 | 1.24 | 1.13 | 1.19 | 1.58 | 1.42 | 1.19 | 2.133 | 1.400 | 2.117 |
| Arachidonic acid | C20:4 n6 | 20.77 | 20.56 | 14.82 | 10.03 | 21.76 | 16.83 | 22.099 | 17.688 | 19.010 |
| cis-11,14,17-Eicosatrienoic acid | C20:3 n3 | 3.82 | 2.92 | 3.56 | 3.56 | 3.96 | 3.66 | 3.342 | 4.251 | 4.687 |
| cis-5,8,11,14,17-Eicosapentaenoic | C20:5 n3 (EPA) | 141.49 | 47.02 | 54.62 | 128.90 | 212.28 | 73.04 | 313.410 | 193.973 | 163.444 |
| Tricosanoic acid | C23:0 | 8.26 | 7.73 | 8.17 | 7.77 | 7.90 | 9.15 | 6.808 | 7.693 | 11.178 |
| Lignoceric acid | C24:0 | 15.80 | 14.75 | 15.62 | 14.76 | 15.12 | 17.38 | 12.969 | 14.710 | 21.319 |
| cis-4,7,10,13,16,19-Docosahexaenoic acid | C22:6 (DHA) | 538.28 | 81.33 | 92.31 | 82.71 | 158.53 | 53.75 | 173.005 | 150.713 | 131.364 |
| omega 6 | 137.01 | 35.78 | 38.13 | 35.94 | 49.09 | 40.27 | 54.58 | 44.44 | 50.32 | |
| omega 3 | 727.57 | 137.26 | 162.75 | 234.49 | 398.08 | 140.05 | 512.39 | 368.80 | 319.68 | |
| omega 6:omega 3 ratio | 0.188 | 0.260 | 0.234 | 0.153 | 0.123 | 0.287 | 0.106 | 0.120 | 0.157 | |
| Peptide Sequence | mol wt | No of AA RESIDUES | Hydrophobicity | GRAVY | Nature | pI | Potential Bioactivities (BIOPEP) |
|---|---|---|---|---|---|---|---|
| VDDHHDDHD | 1240.44 | 10 | 0.57 | −2.61 | very high hydrophilicity | 4.4 | DPP-IV inhibition and antioxidant |
| PVDDHHDDHD | 1337.50 | 11 | 2.2 | −2.52 | very high hydrophilicity | 4.4 | DPP4-IV inhibition and antioxidant |
| NPVDDHHDDHD | 1451.53 | 12 | 3.57 | −2.6 | very high hydrophilicity | 4.4 | DPP4-IV inhibition and antioxidant |
| KPEAPKVP | 864.54 | 8 | 8.9 | −1.26 | high hydrophilicity | 9.9 | ACE-I inhibitor, DPP-IV andDPP-III inhibition and antioxidant |
| NLGAIGH | 680.36 | 7 | 13.74 | 0.37 | moderate hydrophobicity | 7.8 | ACE-I inhibitor, DPP-IV inhibition |
| KGLVDGAPAN | 940.47 | 10 | 16.48 | −0.17 | moderate hydrophobicity | 6.8 | ACE-I inhibitor, DPP-IV inhibition and glucose uptake stimulating |
| FNAEKGFGF | 1015.47 | 9 | 26.99 | −0.17 | moderate hydrophobicity | 6.9 | ACE-I inhibitor, DPP-IV and DPP-III inhibition |
| DHPLPGTD | 850.38 | 8 | 14.49 | −1.34 | moderate hydrophobicity | 3.9 | ACE-I inhibitor, DPP-IV and DPP-III inhibition, anti-amnestic, antithrombotic, regulates stomach mucosal activity |
| SSDVPGV | 659.32 | 7 | 11.81 | 0.19 | moderate hydrophobicity | 3.1 | ACE-I inhibitor, DPP-IV inhibition, anti-amnestic, antithrombotic, regulates stomach mucosal activity |
| GPPGEPGEPGSS | 1066.45 | 12 | 6.37 | −1.38 | very high hydrophilicity | 3.1 | ACE-I inhibitor, antioxidative, DPP-IV and DPPIII inhibition, anti-amnestic, antithrombotic, regulates stomach mucosal activity |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naik, A.S.; Mora, L.; Hayes, M. Characterisation of Seasonal Mytilus edulis By-Products and Generation of Bioactive Hydrolysates. Appl. Sci. 2020, 10, 6892. https://doi.org/10.3390/app10196892
Naik AS, Mora L, Hayes M. Characterisation of Seasonal Mytilus edulis By-Products and Generation of Bioactive Hydrolysates. Applied Sciences. 2020; 10(19):6892. https://doi.org/10.3390/app10196892
Chicago/Turabian StyleNaik, Azza Silotry, Leticia Mora, and Maria Hayes. 2020. "Characterisation of Seasonal Mytilus edulis By-Products and Generation of Bioactive Hydrolysates" Applied Sciences 10, no. 19: 6892. https://doi.org/10.3390/app10196892
APA StyleNaik, A. S., Mora, L., & Hayes, M. (2020). Characterisation of Seasonal Mytilus edulis By-Products and Generation of Bioactive Hydrolysates. Applied Sciences, 10(19), 6892. https://doi.org/10.3390/app10196892