
RNA-Seq Revealed the Effects of Cold Stress on Different Brain Regions of Leiocassis longirostris

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Fish
2.2. Tissue Sampling
2.3. RNA Extraction, Library Construction, and Sequencing
2.4. Transcriptomics (RNA-Seq) Analysis
2.4.1. Sequence Read Processing and Alignment
2.4.2. Functional Annotation and Classification
2.4.3. Analysis of Differentially Expressed Genes (DEGs) and Functional Enrichment
2.5. Weighted Gene Co-Expression Network Analysis (WGCNA)
3. Results
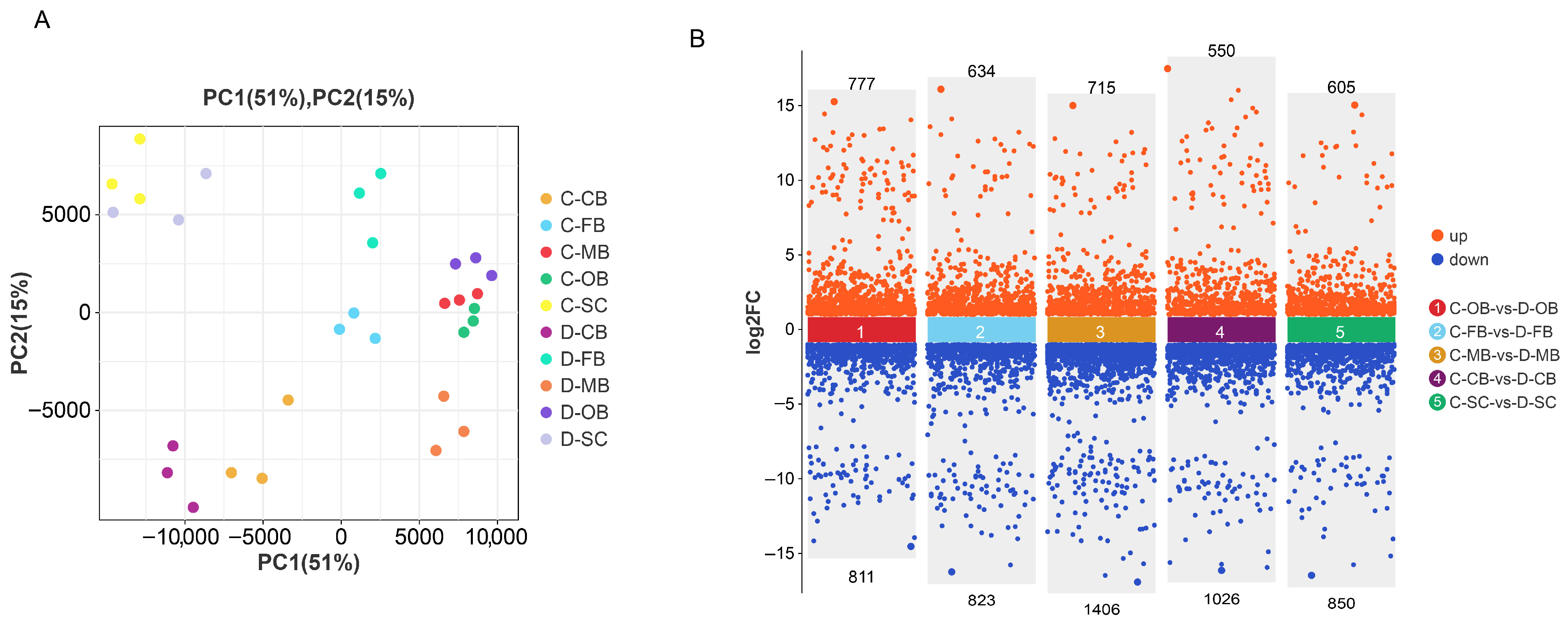
3.1. Transcriptome Sequencing Data Quality Assessment
3.2. Differential Gene Expression Analysis in Brain Tissue
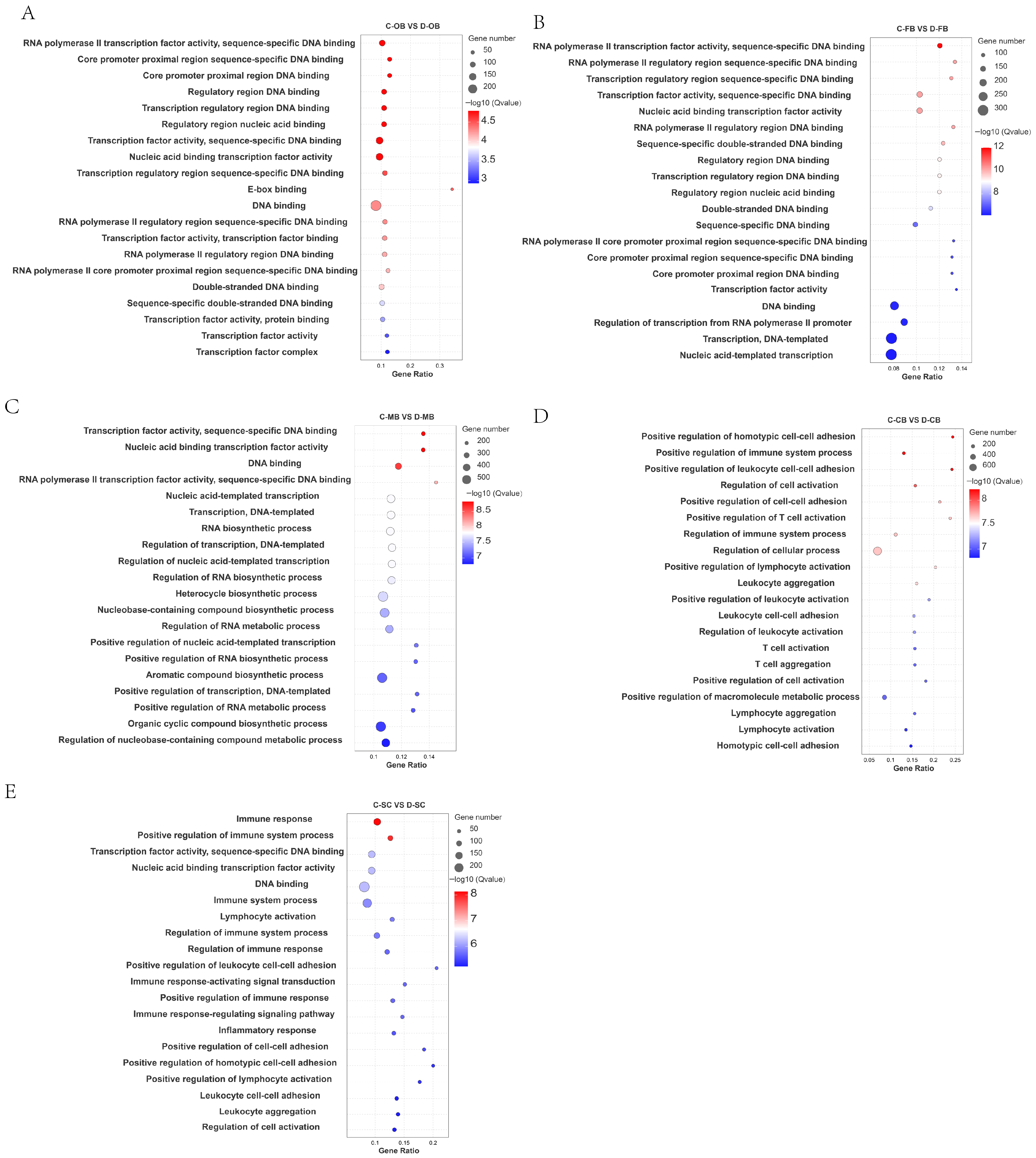
3.3. GO Enrichment of DEGs in Brain Tissue
3.4. KEGG Enrichment of DEGs in Brain Tissue
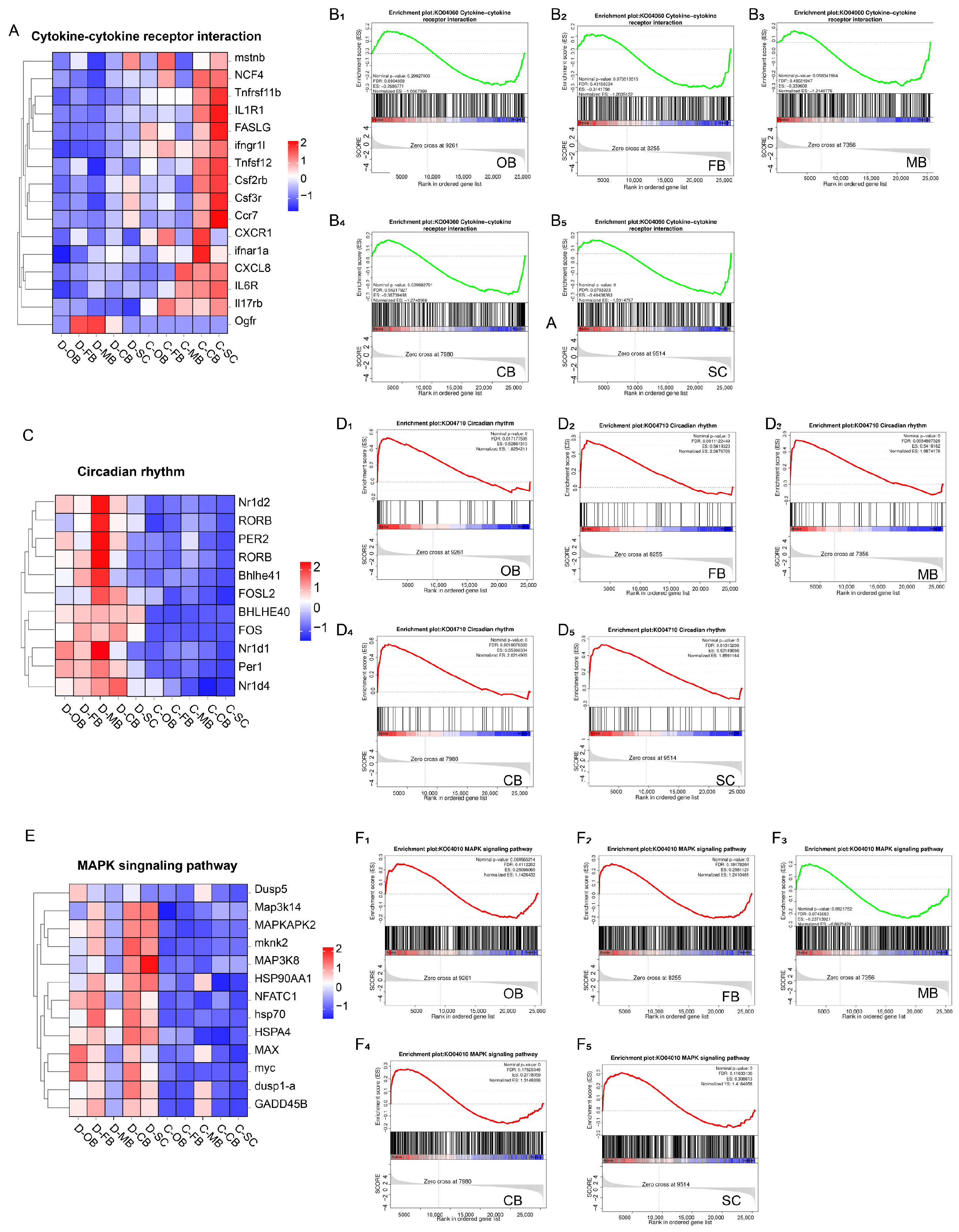
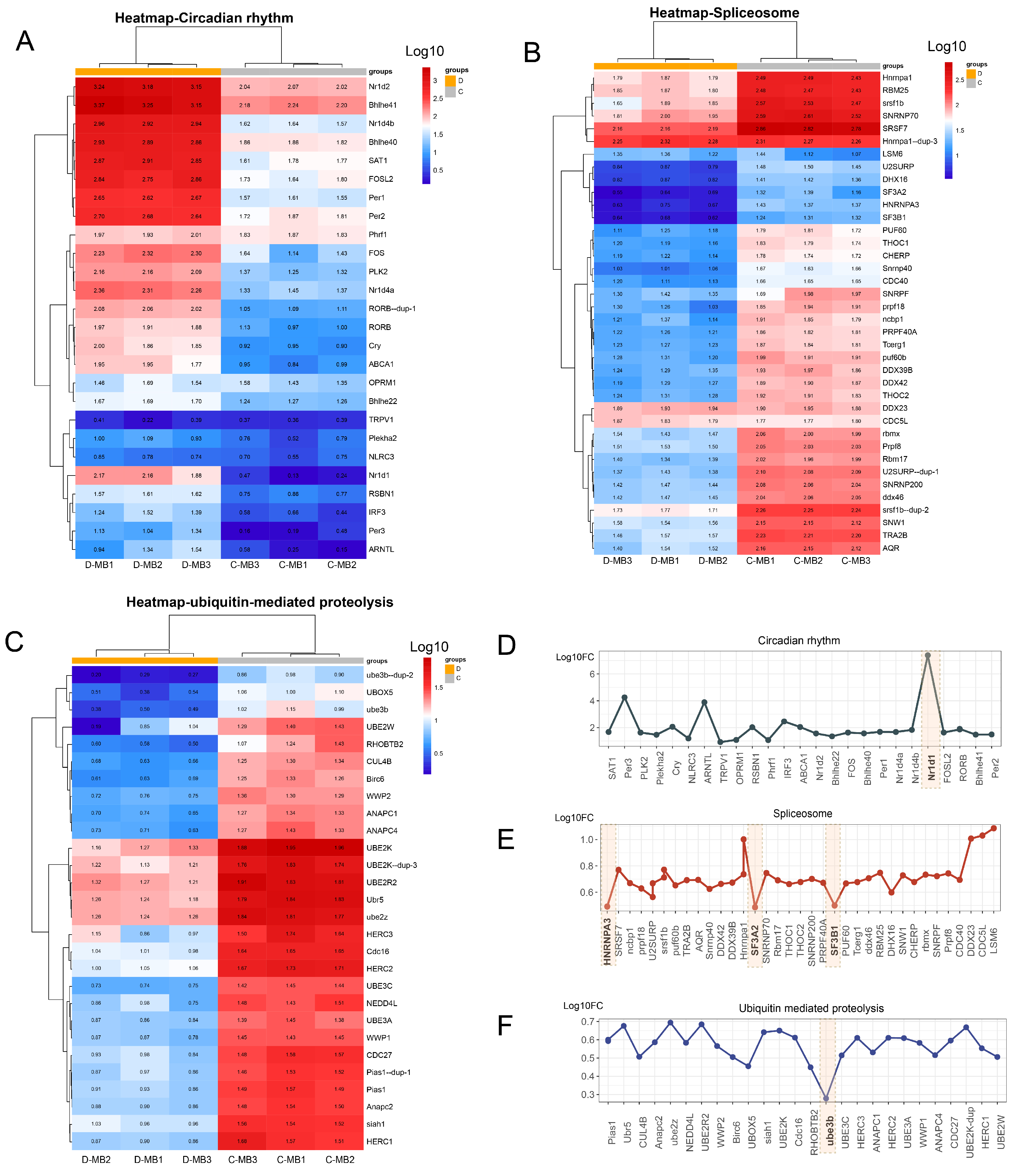
3.5. Effects of Cold Stress on Brain Immunity and Rhythm
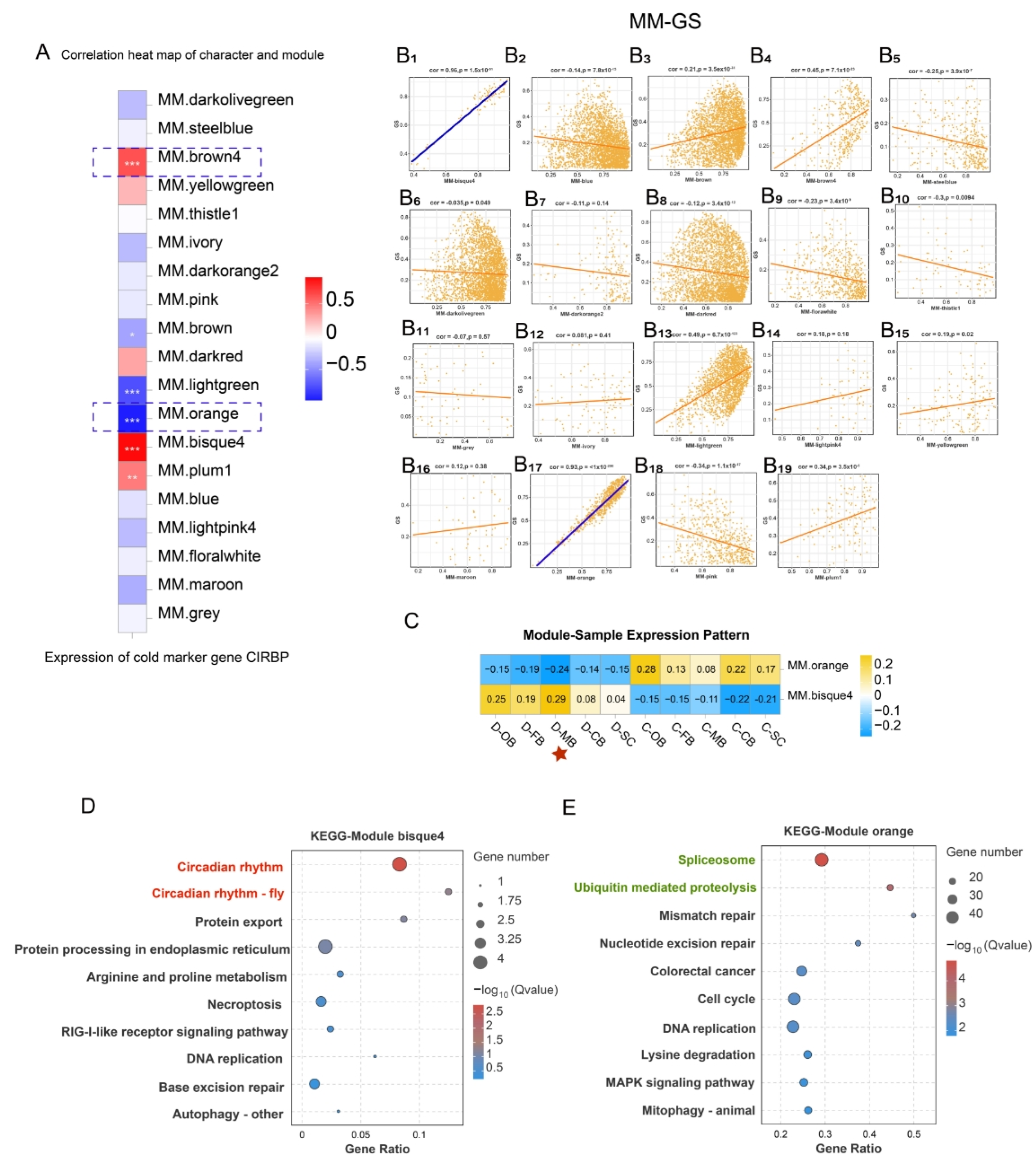
3.6. WGCNA Identifies Cold-Responsive Gene Modules Associated with CIRBP
4. Discussion
4.1. The Mesencephalon Plays an Important Role in the Life Activities of Fish
4.2. Cold Stress Affects the Biological Clock
4.3. Cold Stress Affects Immune Function and Inflammatory Response
4.4. Cold Stress Affects the Gene Expression Process
4.5. Effect of Cold Stress on Ubiquitination
4.6. Effect of Cold Stress on Nutrition Metabolism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feugere, L.; Scott, V.F.; Rodriguez-Barucg, Q.; Beltran-Alvarez, P.; Wollenberg Valero, K.C. Thermal stress induces a positive phenotypic and molecular feedback loop in zebrafish embryos. J. Therm. Biol. 2021, 102, 103114. [Google Scholar] [CrossRef]
- Beers, J.M.; Jayasundara, N. Antarctic notothenioid fish: What are the future consequences of losses and gains acquired during long-term evolution at cold and stable temperatures? J. Exp. Biol. 2015, 218, 1834–1845. [Google Scholar] [CrossRef]
- Francispillai, E.K.; Dietsch, S.M.; Chapman, L.J. Effects of temperature on fish aggression and the combined impact of temperature and turbidity on thermal tolerance. J. Therm. Biol. 2024, 125, 103987. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Peng, C.; Yang, M.; Huang, F.; Duan, X.; Wang, S.; Cheng, H.; Yang, H.; Zhao, H.; Qin, Q. Single-cell RNA-seq landscape midbrain cell responses to red spotted grouper nervous necrosis virus infection. PLoS Pathog. 2021, 17, e1009665. [Google Scholar] [CrossRef] [PubMed]
- Topal, A.; Özdemir, S.; Arslan, H.; Çomaklı, S. How does elevated water temperature affect fish brain? (A neurophysiological and experimental study: Assessment of brain derived neurotrophic factor, cFOS, apoptotic genes, heat shock genes, ER-stress genes and oxidative stress genes). Fish Shellfish. Immunol. 2021, 115, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Ge, G.; Long, Y.; Song, G.; Li, Q.; Cui, Z.; Yan, H. Transcriptomic Profiling Revealed Signaling Pathways Associated with the Spawning of Female Zebrafish under Cold Stress. Int. J. Mol. Sci. 2022, 23, 7494. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, R.; Wang, X.; Zhu, H.; Tian, Z. Transcriptome analysis reveals molecular mechanisms responsive to acute cold stress in the tropical stenothermal fish tiger barb (Puntius tetrazona). BMC Genom. 2020, 21, 737–742. [Google Scholar] [CrossRef]
- Long, Y.; Li, X.; Li, F.; Ge, G.; Liu, R.; Song, G.; Li, Q.; Qiao, Z.; Cui, Z. Transcriptional Programs Underlying Cold Acclimation of Common Carp (Cyprinus carpio L.). Front. Genet. 2020, 11, 556418. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, J.; Ye, Y.; Wei, Q.; Wu, Q. Genetic structure and low-genetic diversity suggesting the necessity for conservation of the Chinese Longsnout catfish, Leiocassis longirostris (Pisces : Bagriidae). Environ. Biol. Fishes 2006, 75, 455–463. [Google Scholar] [CrossRef]
- Zhu, X.; Xie, S.; Zou, Z.; Lei, W.; Cui, Y.; Yang, Y.; Wootton, R.J. Compensatory growth and food consumption in gibel carp, Carassius auratus gibelio, and Chinese longsnout catfish, Leiocassis longirostris, experiencing cycles of feed deprivation and re-feeding. Aquaculture 2004, 241, 235–247. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhao, H.; Wang, X.; Zhang, L.; Mou, C.; Huang, Z.; Ke, H.; Duan, Y.; Zhou, J.; Li, Q. Effects of different temperatures on Leiocassis longirostris gill structure and intestinal microbial composition. Sci. Rep. 2024, 14, 7150. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, Q.; Deng, Y.; Wang, Z.; Feng, Y.; Zhao, H.; Zhao, Z.; Zhang, L.; Duan, Y.; Huang, Z.; et al. RNA-seq revealed the effects of heat stress on different brain regions of Leiocassis longirostris. Front. Physiol. 2025, 16, 1579499. [Google Scholar] [CrossRef] [PubMed]
- He, W.P.; Zhou, J.; Li, Z.; Jing, T.S.; Li, C.H.; Yang, Y.J.; Xiang, M.B.; Zhou, C.W.; Lv, G.J.; Xu, H.Y.; et al. Chromosome-level genome assembly of the Chinese longsnout catfish Leiocassis longirostris. Zool. Res. 2021, 42, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Wang, Y.H.; Ai, C.X.; Zhang, H.; Huang, Y.C.; Zou, W.G. Different cold tolerances among three strains of large yellow croaker: Related to antioxidant defense and energy metabolism. Fish. Physiol. Biochem. 2023, 49, 471–486. [Google Scholar] [CrossRef]
- Madar, V.; Batista, S. FastLSU: A more practical approach for the Benjamini-Hochberg FDR controlling procedure for huge-scale testing problems. Bioinformatics 2016, 32, 1716–1723. [Google Scholar] [CrossRef]
- Morf, J.; Rey, G.; Schneider, K.; Stratmann, M.; Fujita, J.; Naef, F.; Schibler, U. Cold-inducible RNA-binding protein modulates circadian gene expression posttranscriptionally. Science 2012, 338, 379–383. [Google Scholar] [CrossRef]
- Zhong, P.; Huang, H. Recent progress in the research of cold-inducible RNA-binding protein. Future Sci. OA 2017, 3, FSO246. [Google Scholar] [CrossRef]
- Reyes, A.S.; Bittar, A.; Ávila, L.C.; Botia, C.; Esmeral, N.P.; Bloch, N.I. Divergence in brain size and brain region volumes across wild guppy populations. Proc. R. Soc. B Biol. Sci. 2022, 289, 20212784. [Google Scholar] [CrossRef]
- Carbo-Tano, M.; Lapoix, M.; Jia, X.; Thouvenin, O.; Pascucci, M.; Auclair, F.; Quan, F.B.; Albadri, S.; Aguda, V.; Farouj, Y.; et al. The mesencephalic locomotor region recruits V2a reticulospinal neurons to drive forward locomotion in larval zebrafish. Nat. Neurosci. 2023, 26, 1775–1790. [Google Scholar] [CrossRef]
- Bass, J.; Takahashi, J.S. Circadian integration of metabolism and energetics. Science 2010, 330, 1349–1354. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, Y.; Zhang, H.; Xu, J.; Gao, X.; Zhang, T.; Liu, X.; Guo, L.; Zhao, D. Environmental F actors coordinate circadian clock function and rhythm to regulate plant development. Plant Signal Behav. 2023, 18, 2231202. [Google Scholar] [CrossRef] [PubMed]
- Reppert, S.M.; Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002, 418, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yoo, S.H.; Takahashi, J.S. Development and Therapeutic Potential of Small-Molecule Modulators of Circadian Systems. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 231–252. [Google Scholar] [CrossRef] [PubMed]
- López-Olmeda, J.F.; Sánchez-Vázquez, F.J. Zebrafish temperature selection and synchronization of locomotor activity circadian rhythm to ahemeral cycles of light and temperature. Chronobiol. Int. 2009, 26, 200–218. [Google Scholar] [CrossRef]
- Hu, P.; Liu, M.; Liu, Y.; Wang, J.; Zhang, D.; Niu, H.; Jiang, S.; Wang, J.; Zhang, D.; Han, B.; et al. Transcriptome comparison reveals a genetic network regulating the lower temperature limit in fish. Sci. Rep. 2016, 6, 28952. [Google Scholar] [CrossRef]
- Yin, L.; Wu, N.; Curtin, J.C.; Qatanani, M.; Szwergold, N.R.; Reid, R.A.; Waitt, G.M.; Parks, D.J.; Pearce, K.H.; Wisely, G.B.; et al. Rev-erbα, a heme sensor that coordinates metabolic and circadian pathways. Science 2007, 318, 1786–1789. [Google Scholar] [CrossRef]
- Rizzi, A.; Saccia, M.; Benagiano, V. Is the Cerebellum Involved in the Nervous Control of the Immune System Function? Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 546–557. [Google Scholar] [CrossRef]
- Schomberg, D.; Olson, J.K. Immune responses of microglia in the spinal cord: Contribution to pain states. Exp. Neurol. 2012, 234, 262–270. [Google Scholar] [CrossRef]
- Liang, O.D.; Korff, T.; Eckhardt, J.; Rifaat, J.; Baal, N.; Herr, F.; Preissner, K.T.; Zygmunt, M. Oncodevelopmental α-fetoprotein acts as a selective proangiogenic factor on endothelial cell from the fetomaternal unit. J. Clin. Endocrinol. Metab. 2004, 89, 1415–1422. [Google Scholar] [CrossRef]
- Ren, Q.; Guo, F.; Tao, S.; Huang, R.; Ma, L.; Fu, P. Flavonoid fisetin alleviates kidney inflammation and apoptosis via inhibiting Src-mediated NF-κB p65 and MAPK signaling pathways in septic AKI mice. Biomed. Pharmacother. 2020, 122, 109772. [Google Scholar] [CrossRef]
- Müller, F.S.; Aherrahrou, Z.; Grasshoff, H.; Heidorn, M.W.; Humrich, J.Y.; Johanson, L.; Aherrahrou, R.; Reinberger, T.; Schulz, A.; Ten Cate, V.; et al. Autoantibodies against the chemokine receptor 3 predict cardiovascular risk. Eur. Heart J. 2023, 44, 4935–4949. [Google Scholar] [CrossRef]
- Rino, J.; Carmo-Fonseca, M. The spliceosome: A self-organized macromolecular machine in the nucleus? Trends Cell Biol. 2009, 19, 375–384. [Google Scholar] [CrossRef]
- Will, C.L.; Lührmann, R. Spliceosome structure and function. Cold Spring Harb. Perspect. Biol. 2011, 3, a003707. [Google Scholar] [CrossRef] [PubMed]
- Gracey, A.Y.; Fraser, E.J.; Li, W.; Fang, Y.; Taylor, R.R.; Rogers, J.; Brass, A.; Cossins, A.R. Coping with cold: An integrative, multitissue analysis of the transcriptome of a poikilothermic vertebrate. Proc. Natl. Acad. Sci. USA 2004, 101, 16970. [Google Scholar] [CrossRef] [PubMed]
- Keren, H.; Lev-Maor, G.; Ast, G. Alternative splicing and evolution: Diversification, exon definition and function. Nat. Rev. Genet. 2010, 11, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, M.; Sanchez, G.; Barbier, J.; Corcos, L.; Auboeuf, D. The emerging role of pre-messenger RNA splicing in stress responses: Sending alternative messages and silent messengers. RNA Biol. 2011, 8, 740–747. [Google Scholar] [CrossRef]
- Hoskins, A.A.; Moore, M.J. The spliceosome: A flexible, reversible macromolecular machine. Trends Biochem. Sci. 2012, 37, 179–188. [Google Scholar] [CrossRef]
- Ciechanover, A. Proteolysis: From the lysosome to ubiquitin and the proteasome. Nat. Rev. Mol. Cell Biol. 2005, 6, 79–87. [Google Scholar] [CrossRef]
- Ravid, T.; Hochstrasser, M. Diversity of degradation signals in the ubiquitin-proteasome system. Nat. Rev. Mol. Cell Biol. 2008, 9, 679–689. [Google Scholar] [CrossRef]
- Bornstein, M.R.; Neinast, M.D.; Zeng, X.; Chu, Q.; Axsom, J.; Thorsheim, C.; Li, K.; Blair, M.C.; Rabinowitz, J.D.; Arany, Z. Comprehensive quantification of metabolic flux during acute cold stress in mice. Cell Metab. 2023, 35, 2077–2092.e6. [Google Scholar] [CrossRef]
- Hsieh, S.L.; Chen, Y.N.; Kuo, C.M. Physiological responses, desaturase activity, and fatty acid composition in milkfish (Chanos chanos) under cold acclimation. Aquaculture 2003, 220, 903–918. [Google Scholar] [CrossRef]
- Lidén, M.; Eriksson, U. Understanding retinol metabolism: Structure and function of retinol dehydrogenases. J. Biol. Chem. 2006, 281, 13001–13004. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Monier, M.N. Stimulatory effect of dietary taurine on growth performance, digestive enzymes activity, antioxidant capacity, and tolerance of common carp, Cyprinus carpio L., fry to salinity stress. Fish. Physiol. Biochem. 2018, 44, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Hanna, V.S.; Hafez, E.A.A. Synopsis of arachidonic acid metabolism: A review. J. Adv. Res. 2018, 11, 23–32. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Li, Q.; Deng, Y.; Wang, Z.; Feng, Y.; Zhao, Z.; Zhao, H.; Zhang, L.; Duan, Y.; Huang, Z.; et al. RNA-Seq Revealed the Effects of Cold Stress on Different Brain Regions of Leiocassis longirostris. Animals 2025, 15, 2107. https://doi.org/10.3390/ani15142107
Liu S, Li Q, Deng Y, Wang Z, Feng Y, Zhao Z, Zhao H, Zhang L, Duan Y, Huang Z, et al. RNA-Seq Revealed the Effects of Cold Stress on Different Brain Regions of Leiocassis longirostris. Animals. 2025; 15(14):2107. https://doi.org/10.3390/ani15142107
Chicago/Turabian StyleLiu, Senyue, Qiang Li, Yongqiang Deng, Zhongwei Wang, Yang Feng, Zhongmeng Zhao, Han Zhao, Lu Zhang, Yuanliang Duan, Zhipeng Huang, and et al. 2025. "RNA-Seq Revealed the Effects of Cold Stress on Different Brain Regions of Leiocassis longirostris" Animals 15, no. 14: 2107. https://doi.org/10.3390/ani15142107
APA StyleLiu, S., Li, Q., Deng, Y., Wang, Z., Feng, Y., Zhao, Z., Zhao, H., Zhang, L., Duan, Y., Huang, Z., Zhou, J., & Mou, C. (2025). RNA-Seq Revealed the Effects of Cold Stress on Different Brain Regions of Leiocassis longirostris. Animals, 15(14), 2107. https://doi.org/10.3390/ani15142107