
Positive Effects of Lycopene on Growth Performance, Hepatic Antioxidant Capacity, Intestinal Morphology, and Cecal Microflora of Yellow-Feather Broilers
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Feeding, and Experimental Design
2.2. Growth Performance
2.3. Serum Immune Responses
2.4. Hepatic Antioxidant Capacity
2.5. Ileal Morphology
2.6. Hepatic Morphology and Apoptosis-Related Gene Expression
2.7. Cecal Short-Chain Fatty Acids
2.8. Cecal Microflora Structure
2.9. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Hepatic Antioxidant Capacity
3.3. Immune Parameters
3.4. Intestinal Morphology
3.5. Hepatic Morphology and Oxidative Damage to mRNA Expression
3.6. Cecal SCFAs
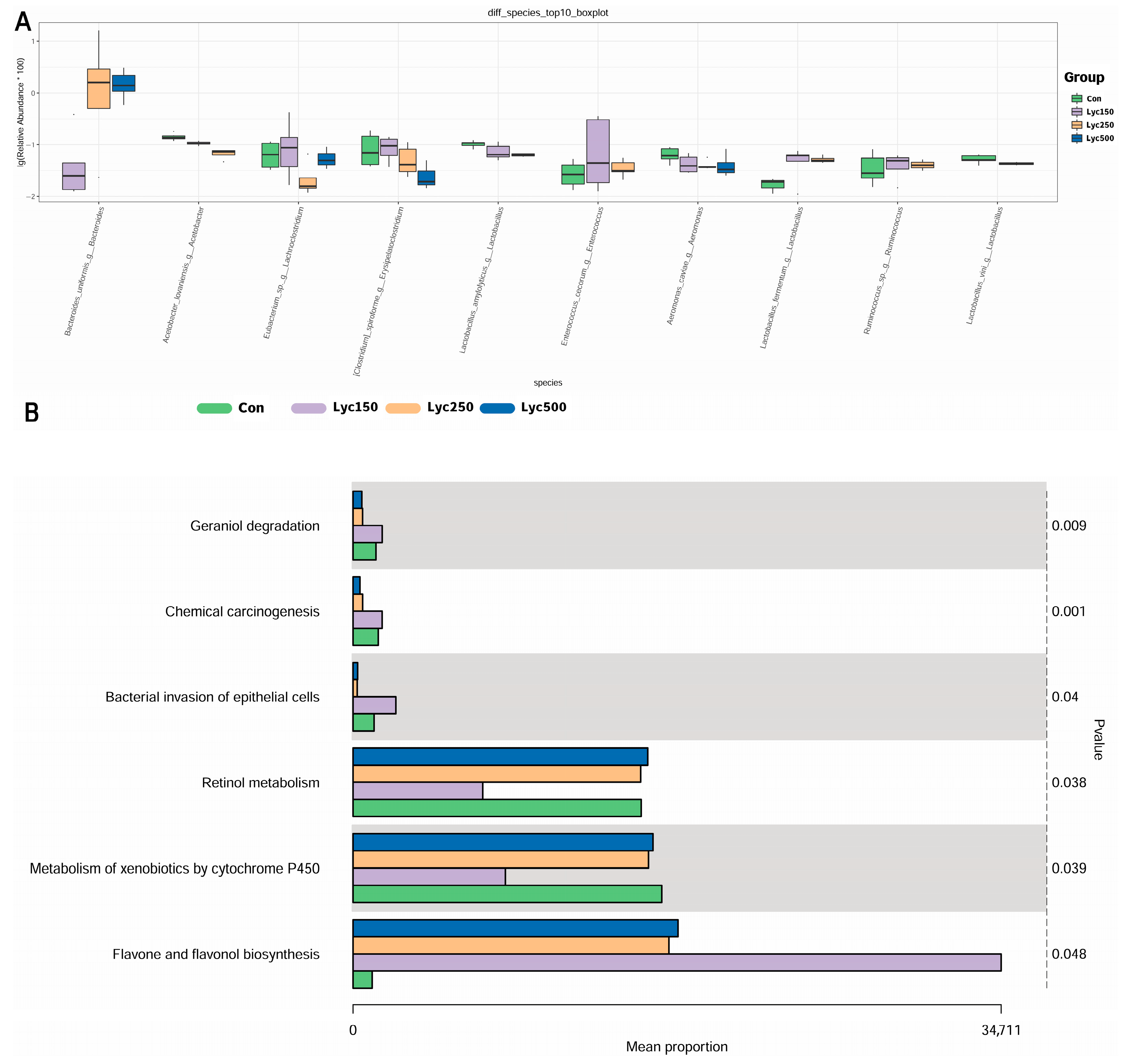
3.7. Cecal Microflora
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Lyc | Lycopene |
| T-AOC | Total antioxidant capacity |
| CAT | Catalase |
| V/C | Villus-to-crypt ratio |
| H&E | Hematoxylin–eosin |
| SEM | Scanning electron microscopy |
| ADG | Average daily gain |
| ADFI | Average daily feed intake |
| F:G | Feed/gain |
| SOD | Superoxide dismutase |
| MDA | Malondialdehyde |
| GSH-Px | Glutathione peroxidase |
| TEM | Transmission electron microscopy |
| SCFAs | Short-chain fatty acids |
| OTUs | Operational taxonomic units |
| PCA | Principal component analysis |
| PCoA | Principal coordinate analysis |
| LEfSe | Linear discriminant analysis effect size |
| ANOVA | Analysis of variance |
References
- Schindler, R.; Mancilla, J.; Endres, S.; Ghorbani, R.; Clark, S.C.; Dinarello, C.A. Correlations and interactions in the production of interleukin-6 (IL-6), IL-1, and tumor necrosis factor (TNF) in human blood mononuclear cells: IL-6 suppresses IL-1 and TNF. Blood 1990, 75, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, S.; Xie, J.; Ji, F.; Peng, W.; Qian, J.; Shen, Q.; Hou, G. Effects of dietary lycopene on the growth performance, antioxidant capacity, meat quality, intestine histomorphology, and cecal microbiota in broiler chickens. Animals 2024, 14, 203. [Google Scholar] [CrossRef] [PubMed]
- Marounek, M.; Pebriansyah, A. Use of carotenoids in feed mixtures for poultry: A review. Agric. Trop. Et Subtrop. 2018, 51, 107–111. [Google Scholar] [CrossRef]
- López-Rull, I.; Hornero-Méndez, D.; Frías, O.; Blanco, G. Age-related relationships between innate immunity and plasma carotenoids in an obligate avian scavenger. PLoS ONE 2015, 10, 1371–1384. [Google Scholar] [CrossRef] [PubMed]
- Duquette, S.C.; Fischer, C.D.; Feener, T.D.; Muench, G.P.; Morck, D.W.; Barreda, D.R.; Nickerson, J.G.; Buret, A.G. Anti-inflammatory effects of retinoids and carotenoid derivatives on caspase-3-dependent apoptosis and efferocytosis of bovine neutrophils. Am. J. Vet. Res. 2014, 75, 1064–1075. [Google Scholar] [CrossRef] [PubMed]
- Kulawik, A.; Cielecka-Piontek, J.; Zalewski, P. The importance of antioxidant activity for the health-promoting effect of lycopene. Nutrients 2023, 15, 3821. [Google Scholar] [CrossRef] [PubMed]
- Jahejo, A.R.; Rajput, N.; Wen-Xia, T.; Naeem, M.; Kalhoro, D.H.; Kaka, A.; Niu, S.; Jia, F. Immunomodulatory and growth promoting wffects of Basil (Ocimum basilicum) and ascorbic acid in heat stressed broiler chickens. Pak. J. Zool. 2019, 51, 801–807. [Google Scholar] [CrossRef]
- Sohail, M.U.; Hume, M.E.; Byrd, J.A.; Nisbet, D.J.; Ijaz, A.; Sohail, A.; Shabbir, M.Z.; Rehman, H. Effect of supplementation of prebiotic mannan-oligosaccharides and probiotic mixture on growth performance of broilers subjected to chronic heat stress. Poult. Sci. 2012, 91, 2235–2240. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.L.; Qin, Q.M.; Li, N.Q. Research progress on the effects of heat stress on immune function in poultry. Chin. Poult. 2021, 43, 99–105. [Google Scholar] [CrossRef]
- Riley, W.W.; Nickerson, J.G.; Burton, G.W. Effect of oxidized β-carotene on the growth and feed efficiency of broilers. Poult. Sci. 2021, 100, 101088. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, H.; Zhu, Y.; Cui, H.; Yang, J.; Lu, M.; Cheng, H.; Gu, L.; Xu, T.; Xu, L. Effect of lycopene on the growth performance, antioxidant enzyme activity, and expression of gene in the Keap1-Nrf2 signaling pathway of Arbor Acres broilers. Front. Vet. Sci. 2022, 9, 833346. [Google Scholar] [CrossRef] [PubMed]
- Hidayat, D.F.; Mahendra, M.Y.N.; Kamaludeen, J.; Pertiwi, H. Lycopene in feed as antioxidant and immuno-modulator improves broiler chicken’s performance under heat-stress conditions. Vet. Med. Int. 2023, 2023, 5418081. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.Y.; Zhu, S.Y.; Chen, J.; Li, M.Z.; Zhao, Y.; Talukder, M.; Li, J.L. Lycopene alleviates di(2-ethylhexyl) phthalate-induced splenic injury by activating P62-Keap1-NRF2 signaling. Food Chem. Toxicol. 2022, 168, 113324. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Wu, J.; Li, J.; Bai, Y.; Luo, Y.; Ji, B.; Xia, B.; Liu, Z.; Tan, X.; Lv, J.; et al. Lycopene alleviates DSS-induced colitis and behavioral disorders via mediating microbes-gut-brain axis balance. J. Agr. Food Chem. 2020, 68, 3963–3975. [Google Scholar] [CrossRef] [PubMed]
- Sarker, M.T.; Wan, X.; Yang, H.; Wang, Z. Dietary lycopene supplementation could alleviate aflatoxin B(1) induced intestinal damage through improving immune function and anti-oxidant capacity in broilers. Animals 2021, 11, 3165. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Nutrient Requirements of Poultry, 9th ed.; The National Academies Press: Washington, DC, USA, 1994; Volume 176. [Google Scholar]
- Cao, G.; Tao, F.; Hu, Y.; Li, Z.; Zhang, Y.; Deng, B.; Zhan, X. Positive effects of a Clostridium butyricum-based compound probiotic on growth performance, immune responses, intestinal morphology, hypothalamic neurotransmitters, and colonic microbiota in weaned piglets. Food Funct. 2019, 10, 2926–2934. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Yu, Y.; Wang, H.; Yang, H.; Tao, F.; Yang, S.; Liu, J.; Li, Z.; Yang, C. Dietary Clostridium butyricum and 25-Hydroxyvitamin D3 modulate bone metabolism of broilers through the gut-brain axis. Poult. Sci. 2024, 103, 103966. [Google Scholar] [CrossRef] [PubMed]
- Lambert, G.P.; Gisolfi, C.V.; Berg, D.J.; Moseley, P.L.; Oberley, L.W.; Kregel, K.C. Selected contribution: Hyperthermia-induced intestinal permeability and the role of oxidative and nitrosative stress. J. Appl. Physiol. 2002, 92, 1750–1761. [Google Scholar] [CrossRef] [PubMed]
- Hosseini-Vashan, S.J.; Golian, A.; Yaghobfar, A. Growth, immune, antioxidant, and bone responses of heat stress-exposed broilers fed diets supplemented with tomato pomace. Int. J. Biometeorol. 2016, 60, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Davidson, J.F.; Schiestl, R.H. Mitochondrial respiratory electron carriers are involved in oxidative stress during heat stress in Saccharomyces cerevisiae. Mol. Cell. Biol. 2001, 21, 8483–8489. [Google Scholar] [CrossRef] [PubMed]
- Slimen, I.; Najar, T.; Ghram, A.; Dabbebi, H.; Mrad, M.B.; Abdrabbah, M. Reactive oxygen species, heat stress and oxidative-induced mitochondrial damage. A review. Int. J. Hyperther 2014, 30, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Chen, C.; Wang, W.; Ma, J.; Xie, Q.; Gao, Y.; Chen, F.; Zhang, X.; Bi, Y. Effects of lycopene supplementation in both maternal and offspring diets on growth performance, antioxidant capacity and biochemical parameters in chicks. J. Anim. Physiol. Anim. Nutr. 2015, 99, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Arain, M.A.; Mei, Z.; Hassan, F.U.; Saeed, M.; Rajput, I.R. Lycopene: A natural antioxidant for prevention of heat-induced oxidative stress in poultry. World Poult. Sci. J. 2018, 74, 89–100. [Google Scholar] [CrossRef]
- Liu, G.; Wang, T.; Wang, T.; Song, J.; Zhou, Z. Effects of apoptosis-related proteins caspase 3, Bax and Bcl-2 on cerebral ischemia rats. Biomed. Rep. 2013, 1, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Fu, G.; Liu, S.; Li, L.; Zhao, X. Effects of oxygen levels and a Lactobacillus plantarum strain on mortality and immune response of chickens at high altitude. Sci. Rep. 2019, 9, 16037. [Google Scholar] [CrossRef] [PubMed]
- Brusca, S.B.; Elinoff, J.M.; Zou, Y.; Jang, M.K.; Kong, H.; Demirkale, C.Y.; Sun, J.; Seifuddin, F.; Pirooznia, M.; Valantine, H.A.; et al. Plasma cell-free DNA predicts survival and maps specific sources of injury in pulmonary arterial hypertension. Circulation 2022, 146, 1033–1045. [Google Scholar] [CrossRef] [PubMed]
- Yonar, M.E. The effect of lycopene on oxytetracycline-induced oxidative stress and immunosuppression in rainbow trout (Oncorhynchus mykiss, W.). Fish Shellfish Immunol. 2012, 32, 994–1001. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Guo, M.H.; Hai, X. Hepatoprotective and antioxidant effects of lycopene on non-alcoholic fatty liver disease in rats. World J. Gastroenterol. 2016, 22, 10180–10188. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Onderci, M.; Sahin, N.; Gursu, M.F.; Khachik, F.; Kuculk, O. Effects of lycopene supplementation on antioxidant status, oxidative stress, performance and carcass characteristics in heat-stressed Japanese quail. J. Therm. Biol. 2005, 31, 307–312. [Google Scholar] [CrossRef]
- Rajput, S.A.; Liang, S.J.; Wang, X.Q.; Yan, H.C. Lycopene protects intestinal epithelium from deoxynivalenol-induced oxidative damage via regulating Keap1/Nrf2 signaling. Antioxidants 2021, 10, 1493. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Jha, R. Strategies to modulate the intestinal microbiota and their effects on nutrient utilization, performance, and health of poultry. J. Anim. Sci. Biotechnol. 2019, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ji, H.; Wang, S.; Liu, H.; Zhang, W.; Zhang, D.; Wang, Y. Probiotic lactobacillus plantarum promotes intestinal barrier function by strengthening the epithelium and modulating gut microbiota. Front. Microbiol. 2018, 9, 1953. [Google Scholar] [CrossRef] [PubMed]
- Gaafar, A.A.; Asker, M.S.; Salama, Z.A. In vitro, antiviral, antimicrobial and antioxidant potential activity of tomato pomace. Int. J. Pharm. Sci. Res. 2015, 32, 262–272. [Google Scholar]
- Kang, K.; Oh, J.Y.; Cha, S.Y.; Kim, W.I.; Cho, H.S.; Jang, H.K. Efficacy of polymers from spontaneous carotenoid oxidation in reducing necrotic enteritis in broilers. Poult. Sci. 2018, 97, 3058–3062. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liu, Y.; Cao, G.; Liu, J.; Xiao, S.; Xiao, P.; Tao, Y.; Gao, H. Effects of lycopene on the growth performance, meat quality, cecal metagenome, and hepatic untargeted metabolome in heat-stressed broilers. Poult. Sci. 2024, 103, 104299. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Rajarammohan, S.; Thakur, R.; Hassan, M. Linear and branched β-Glucans degrading enzymes from versatile Bacteroides uniformis JCM 13288T and their roles in cooperation with gut bacteria. Gut Microbes 2020, 12, 1826761. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.W.; Chen, H.C.; Chen, C.Y.; Yen, C.Y.; Lin, C.J.; Prajnamitra, R.P.; Chen, L.; Ruan, S.; Lin, J.; Lin, P.; et al. Loss of gut microbiota alters immune system composition and cripples postinfarction cardiac repair. Circulation 2019, 139, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Jiao, S.; Guo, Q.; Zhou, M.; Ren, W.; Zhao, Y.; Li, D.; Yang, W.; Dai, S.; Pei, Y.; et al. Lycopene regulates intestinal dysmotility and behavioral disorders via regulating enteric neurons and cholinergic system. Food Funct. 2025, 16, 5363–5379. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Stage (d) | |
|---|---|---|
| 1–28 | 29–56 | |
| composition (%) | ||
| corn | 52 | 60.3 |
| soybean meal | 24.7 | 14 |
| extruded soybean | 6 | 5 |
| corn lees | 4 | 8 |
| corn protein powder | 2 | 4 |
| corn bran | 0.6 | |
| soybean oil | 3.5 | 2.8 |
| limestone | 1.3 | 1.3 |
| fermented soybean meal | 2.5 | 2.5 |
| vitamin and mineral 1 | 4 | 4 |
| total | 100.00 | 100.00 |
| nutrition level 2 | ||
| AME (Kcal/kg) | 3050 | 3125 |
| CP (%) | 21 | 18.2 |
| L-lysine HCl (%) | 1.25 | 0.97 |
| DL-methionine (%) | 0.55 | 0.44 |
| cysteine (%) | 0.89 | 0.76 |
| threonine (%) | 0.92 | 0.74 |
| tryptophan (%) | 0.23 | 0.19 |
| Ca (%) | 0.86 | 0.76 |
| available p (%) | 0.59 | 0.48 |
| Gene | Position | Prime Sequence (5′-3′) | Length |
|---|---|---|---|
| GADPH | Forward | AGAACATCATCCCAGCGTCC | 20 |
| Reverse | CGGCAGGTCAGGTCAACAAC | 20 | |
| Caspase-3 | Forward | GGCTCCTGGTTTATTCAGTCTC | 22 |
| Reverse | ATTCTGCCACTCTGCGATTT | 20 | |
| Caspase-9 | Forward | ACAGTGGCAGGGTCCTCAAACAGA | 24 |
| Reverse | GTCACGCAGGGCAAAGAAACTCAG | 24 | |
| Bcl-2 | Forward | TCGCGCCGCTACCAGAGGGACTTC | 24 |
| Reverse | CCGGTTGACGCTCTCGACGCACAT | 24 | |
| Bax | Forward | ATCGTCGCCTTCTTCGAGTT | 20 |
| Reverse | ATCCCATCCTCCGTTGTCCT |
| Item | Con | Lyc150 | Lyc250 | Lyc500 | SEM | p-Value |
|---|---|---|---|---|---|---|
| 1–28 d | ||||||
| ADG/g | 24.73 | 25.18 | 25.22 | 24.98 | 0.344 | 0.493 |
| ADFI/g | 42.59 b | 44.31 ab | 45.32 a | 43.78 ab | 0.738 | 0.021 |
| F/G | 1.80 | 1.73 | 1.77 | 1.72 | 0.028 | 0.053 |
| 29–56 d | ||||||
| ADG/g | 41.24 b | 47.07 a | 47.23 a | 47.34 a | 1.660 | 0.007 |
| ADFI/g | 111 | 112.5 | 119.1 | 117.3 | 4.450 | 0.270 |
| F/G | 2.80 | 2.55 | 2.71 | 2.61 | 0.084 | 0.057 |
| 1–56 d | ||||||
| ADG/g | 33.53 | 36.1 | 36.82 | 36.90 | 1.845 | 0.070 |
| ADFI/g | 67.14 | 72.23 | 71.18 | 71.42 | 0.148 | 0.328 |
| F/G | 2.52 | 2.25 | 2.38 | 2.46 | 1.412 | 0.108 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, G.; Liu, X.; Wang, H.; Kang, J.; Wang, F.; Li, M.; Sun, W.; Lv, X. Positive Effects of Lycopene on Growth Performance, Hepatic Antioxidant Capacity, Intestinal Morphology, and Cecal Microflora of Yellow-Feather Broilers. Animals 2025, 15, 2108. https://doi.org/10.3390/ani15142108
Cao G, Liu X, Wang H, Kang J, Wang F, Li M, Sun W, Lv X. Positive Effects of Lycopene on Growth Performance, Hepatic Antioxidant Capacity, Intestinal Morphology, and Cecal Microflora of Yellow-Feather Broilers. Animals. 2025; 15(14):2108. https://doi.org/10.3390/ani15142108
Chicago/Turabian StyleCao, Guangtian, Xiyue Liu, Huixian Wang, Jia Kang, Feiyang Wang, Molin Li, Wenqi Sun, and Xiaosi Lv. 2025. "Positive Effects of Lycopene on Growth Performance, Hepatic Antioxidant Capacity, Intestinal Morphology, and Cecal Microflora of Yellow-Feather Broilers" Animals 15, no. 14: 2108. https://doi.org/10.3390/ani15142108
APA StyleCao, G., Liu, X., Wang, H., Kang, J., Wang, F., Li, M., Sun, W., & Lv, X. (2025). Positive Effects of Lycopene on Growth Performance, Hepatic Antioxidant Capacity, Intestinal Morphology, and Cecal Microflora of Yellow-Feather Broilers. Animals, 15(14), 2108. https://doi.org/10.3390/ani15142108