
Dietary Supplementation of Novel Aflatoxin Oxidase CotA Alleviates Aflatoxin B1-Induced Oxidative Stress, Lipid Metabolism Disorder, and Apoptosis in the Liver of Japanese Quails
 ,
,  ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Animal Trial Design
2.2. Growth Performance and Sample Collection
2.3. H&E Staining
2.4. Oil Red O Staining
2.5. TUNEL Staining
2.6. Serum Biochemical Analysis
2.7. Liver Antioxidant and Oxidative Biomarkers
2.8. Quantitative Real-Time PCR
2.9. Determination of AFB1 Residues and AFB1-DNA Adduct Levels
2.10. Statistical Analysis
3. Results
3.1. Aflatoxin Oxidase CotA Improved the Growth Performance of Japanese Quails Fed with AFB1-Contaminated Diet
3.2. Aflatoxin Oxidase CotA Protected Japanese Quails from AFB1-Induced Liver Injury
3.3. Aflatoxin Oxidase CotA Alleviated AFB1-Induced Oxidative Stress in Liver
3.4. Aflatoxin Oxidase CotA Ameliorated AFB1-Induced Lipid Metabolism Disorder in Liver
3.5. Aflatoxin Oxidase CotA Mitigated AFB1-Induced Liver Apoptosis
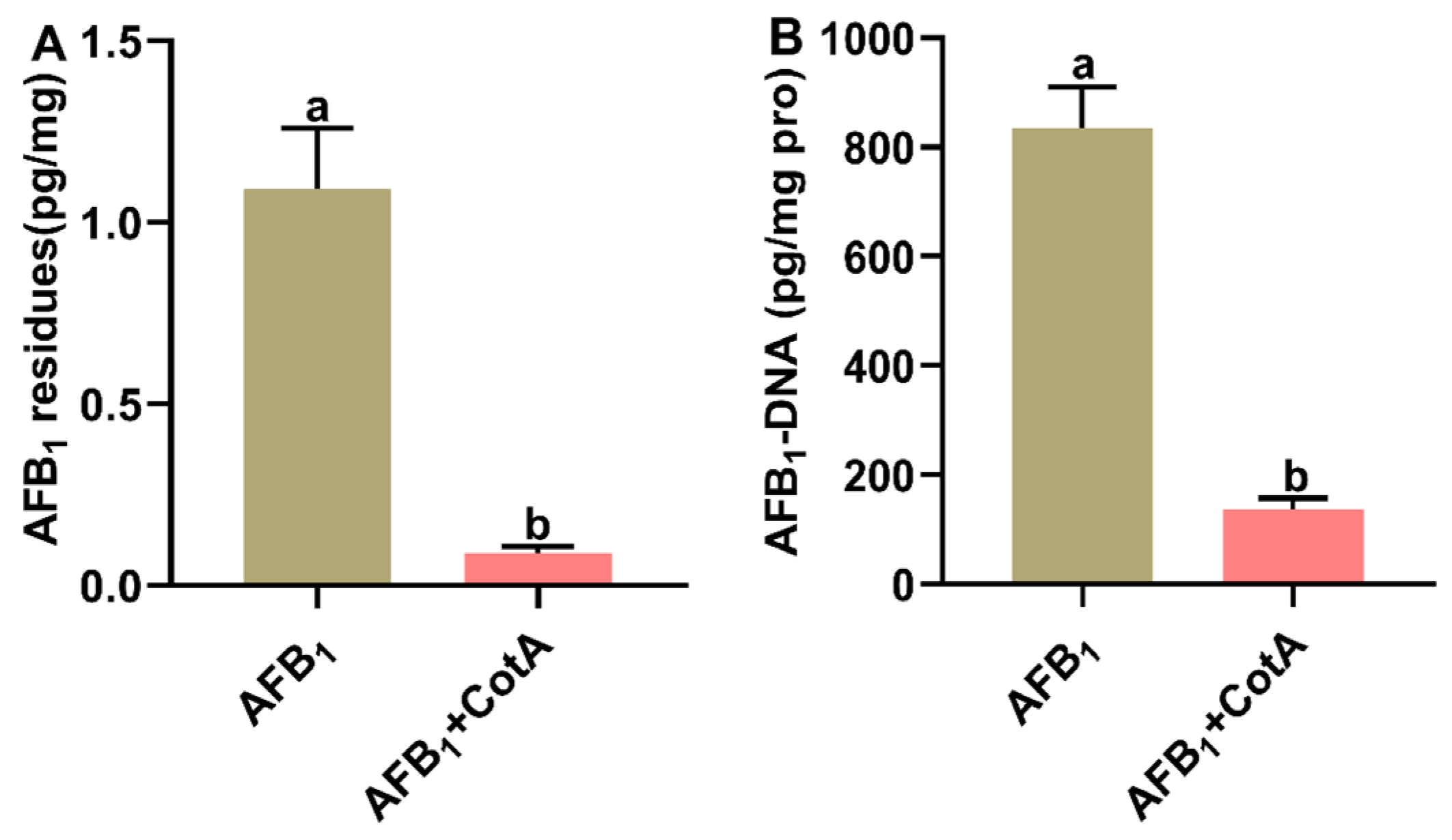
3.6. Aflatoxin Oxidase CotA Reduced AFB1 Residues and AFB1-DNA Adduct Content in Liver
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van der Zijden, A.S.M.; Koelensmid, W.; Boldingh, J.; Barrett, C.B.; Ord, W.O.; Philip, J. Aspergillus flavus and Turkey X disease: Isolation in crystalline form of a toxin responsible for Turkey X-disease. Nature 1962, 195, 1060–1062. [Google Scholar] [CrossRef]
- Blount, W.P. Turkey “X” disease. J. Br. Turkey 1961, 9, 55–58. [Google Scholar]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens-the IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, L.; Ma, Q.; Ji, C. Novel strategies for degradation of aflatoxins in food and feed: A review. Food Res. Int. 2021, 140, 109878. [Google Scholar] [CrossRef]
- Benkerroum, N. Retrospective and prospective look at aflatoxin research and development from a practical standpoint. Int. J. Environ. Res. Public Health 2019, 16, 3633. [Google Scholar] [CrossRef]
- Fouad, A.M.; Ruan, D.; El-Senousey, H.K.; Chen, W.; Jiang, S.; Zheng, C. Harmful effects and control strategies of aflatoxin B1 produced by Aspergillus flavus and Aspergillus parasiticus strains on poultry: Review. Toxins 2019, 11, 176. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Li, Q. Aflatoxin B1 in poultry liver: Toxic mechanism. Toxicon 2023, 233, 107262. [Google Scholar] [CrossRef]
- Luo, Y.; Liu, X.; Yuan, L.; Li, J. Complicated interactions between bio-adsorbents and mycotoxins during mycotoxin adsorption: Current research and future prospects. Trends Food Sci. Technol. 2020, 96, 127–134. [Google Scholar] [CrossRef]
- Hu, S.; Xu, C.; Lu, P.; Wu, M.; Chen, A.; Zhang, M.; Xie, Y.; Han, G. Widespread distribution of the DyP-carrying bacteria involved in the aflatoxin B1 biotransformation in Proteobacteria and Actinobacteria. J. Hazard. Mater. 2024, 478, 135493. [Google Scholar] [CrossRef]
- Adegoke, T.V.; Yang, B.; Tian, X.; Yang, S.; Gao, Y.; Ma, J.; Wang, G.; Si, P.; Li, R.; Xing, F. Simultaneous degradation of aflatoxin B1 and zearalenone by porin and peroxiredoxin enzymes cloned from Acinetobacter nosocomialis Y1. J. Hazard. Mater. 2023, 459, 132105. [Google Scholar] [CrossRef]
- Loi, M.; Fanelli, F.; Zucca, P.; Liuzzi, V.C.; Quintieri, L.; Cimmarusti, M.T.; Monaci, L.; Haidukowski, M.; Logrieco, A.F.; Sanjust, E.; et al. Aflatoxin B1 and M1 degradation by Lac2 from Pleurotus pulmonarius and redox mediators. Toxins 2016, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Qin, X.; Tang, Y.; Ma, Q.; Zhang, J.; Zhao, L. CotA laccase, a novel aflatoxin oxidase from Bacillus licheniformis, transforms aflatoxin B1 to aflatoxin Q1 and epi-aflatoxin Q1. Food Chem. 2020, 325, 126877. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Wu, T.; Zhang, H.; Sun, Z.; Mwabulili, F.; Xie, Y.; Sun, S.; Ma, W.; Li, Q.; Yang, Y.; et al. Mining lactonase gene from aflatoxin B1-degrading strain Bacillus megaterium and degrading properties of the recombinant enzyme. J. Agric. Food Chem. 2023, 71, 20762–20771. [Google Scholar] [CrossRef]
- Graham, D.E. A new role for coenzyme F420 in aflatoxin reduction by soil mycobacteria. Mol. Microbiol. 2010, 78, 533–536. [Google Scholar] [CrossRef]
- Liu, M.; Zhao, L.; Gong, G.; Zhang, L.; Shi, L.; Dai, J.; Han, Y.; Wu, Y.; Khalil, M.M.; Sun, L. Invited review: Remediation strategies for mycotoxin control in feed. J. Anim. Sci. Biotechnol. 2022, 13, 19. [Google Scholar] [CrossRef]
- Ma, M.; Wang, Q.; Liu, Y.; Li, G.; Liu, L.; Wang, G.; Guo, Y.; Huang, S.; Ma, Q.; Ji, C.; et al. Bacillus CotA laccase improved the intestinal health, amino acid metabolism and hepatic metabolic capacity of Pekin ducks fed naturally contaminated AFB1 diet. J. Anim. Sci. Biotechnol. 2024, 15, 138. [Google Scholar] [CrossRef]
- Sakamoto, M.I.; Murakami, A.E.; Fernandes, A.M.; Ospina-Rojas, I.C.; Nunes, K.C.; Hirata, A.K. Performance and serum biochemical profile of Japanese quail supplemented with silymarin and contaminated with aflatoxin B1. Poult. Sci. 2018, 97, 159–166. [Google Scholar] [CrossRef]
- Bagherzadeh Kasmani, F.; Karimi Torshizi, M.A.; Allameh, A.; Shariatmadari, F. A novel aflatoxin-binding Bacillus probiotic: Performance, serum biochemistry, and immunological parameters in Japanese quail. Poult. Sci. 2012, 91, 1846–1853. [Google Scholar] [CrossRef]
- Nazar, F.N.; Magnoli, A.P.; Dalcero, A.M.; Marin, R.H. Effect of feed contamination with aflatoxin B1 and administration of exogenous corticosterone on Japanese quail biochemical and immunological parameters. Poult. Sci. 2012, 91, 47–54. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, L.; Huang, Z.; Guo, Y.; Tang, Y.; Wang, Y.; Ma, Q.; Zhao, L. Combined strategies for improving aflatoxin B1 degradation ability and yield of a Bacillus licheniformis CotA laccase. Int. J. Mol. Sci. 2024, 25, 6455. [Google Scholar] [CrossRef]
- Rawal, S.; Kim, J.E.; Coulombe, R. Aflatoxin B1 in poultry: Toxicology, metabolism and prevention. Res. Vet. Sci. 2010, 89, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Ding, K.; Wang, J.; Deng, Q.; Gu, K.; Wang, J. Effects of lactic acid bacteria and smectite after aflatoxin B1 challenge on the growth performance, nutrient digestibility and blood parameters of broilers. J. Anim. Physiol. Anim. Nutr. 2018, 102, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Habashy, W.S.; Milfort, M.C.; Rekaya, R.; Aggrey, S.E. Cellular antioxidant enzyme activity and biomarkers for oxidative stress are affected by heat stress. Int. J. Biometeorol. 2019, 63, 1569–1584. [Google Scholar] [CrossRef]
- Lennicke, C.; Rahn, J.; Lichtenfels, R.; Wessjohann, L.A.; Seliger, B. Hydrogen peroxide—Production, fate and role in redox signaling of tumor cells. Cell Commun. Signal. 2015, 13, 39. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Xu, F.; Li, Y.; Cao, Z.; Zhang, J.; Huang, W. AFB1-induced mice liver injury involves mitochondrial dysfunction mediated by mitochondrial biogenesis inhibition. Ecotoxicol. Environ. Saf. 2021, 216, 112213. [Google Scholar] [CrossRef]
- Guo, J.; Yan, W.; Tang, J.; Jin, X.; Xue, H.; Wang, T.; Zhang, L.; Sun, Q.; Liang, Z. Dietary phillygenin supplementation ameliorates aflatoxin B1-induced oxidative stress, inflammation, and apoptosis in chicken liver. Ecotoxicol. Environ. Saf. 2022, 236, 113481. [Google Scholar] [CrossRef]
- Wu, G.; San, J.; Pang, H.; Du, Y.; Li, W.; Zhou, X.; Yang, X.; Hu, J.; Yang, J. Taurine attenuates AFB1-induced liver injury by alleviating oxidative stress and regulating mitochondria-mediated apoptosis. Toxicon 2022, 215, 17–27. [Google Scholar] [CrossRef]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar]
- Rotimi, O.; Rotimi, S.; Duru, C.; Ebebeinwe, O.; Abiodun, A.; Oyeniyi, B.; Faduyile, F. Acute aflatoxin B1-induced hepatotoxicity alters gene expression and disrupts lipid and lipoprotein metabolism in rats. Toxicol. Rep. 2017, 4, 408–414. [Google Scholar] [CrossRef]
- Ren, X.L.; Han, P.; Meng, Y. Aflatoxin B1-induced COX-2 expression promotes mitophagy and contributes to lipid accumulation in hepatocytes in vitro and in vivo. Int. J. Toxicol. 2020, 39, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, J.; Kang, W.; Liu, S.; Liu, J.; Shi, M.; Wang, Y.; Liu, X.; Chen, X.; Huang, K. Aflatoxin B1 induces liver injury by disturbing gut microbiota-bile acid-FXR axis in mice. Food Chem. Toxicol. 2023, 176, 113751. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yang, X.; Liu, F.; Wang, X.; Zhang, X.; He, K.; Wang, H. Comprehensive metabolomic analysis reveals dynamic metabolic reprogramming in Hep3B cells with aflatoxin B1 exposure. Toxins 2021, 13, 384. [Google Scholar] [CrossRef] [PubMed]
- Kiraz, Y.; Adan, A.; Kartal Yandim, M.; Baran, Y. Major apoptotic mechanisms and genes involved in apoptosis. Tumor Biol. 2016, 37, 8471–8486. [Google Scholar] [CrossRef]
- Nabi, F.; Tao, W.; Ye, R.; Li, Z.; Lu, Q.; Shang, Y.; Hu, Y.; Fang, J.; Bhutto, Z.; Liu, J. Penthorum Chinense Pursh extract alleviates aflatoxin B1-induced liver injury and oxidative stress through mitochondrial pathways in broilers. Front. Vet. Sci. 2022, 9, 822259. [Google Scholar] [CrossRef]
- Bedard, L.L.; Massey, T.E. Aflatoxin B1-induced DNA damage and its repair. Cancer Lett. 2006, 241, 174–183. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items 1 | Treatment 2 | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | AFB1 | AFB1+CotA | |||
| Initial BW (g) | 69.43 | 69.52 | 69.84 | 0.10 | 0.188 |
| Final BW (g) | 156.63 a | 149.40 b | 158.57 a | 1.43 | 0.009 |
| ADG (g) | 4.15 a | 3.80 b | 4.23 a | 0.07 | 0.009 |
| ADFI (g) | 17.38 | 17.09 | 18.25 | 0.27 | 0.203 |
| FCR | 4.19 | 4.51 | 4.32 | 0.09 | 0.381 |
| Item 1 | Treatment 2 | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | AFB1 | AFB1+CotA | |||
| T-AOC (mmol/g prot) | 1.71 a | 1.26 b | 1.94 a | 0.10 | 0.004 |
| T-SOD (U/mg prot) | 17.99 | 14.58 | 18.43 | 0.97 | 0.216 |
| GST (U/mg prot) | 117.77 a | 78.45 b | 91.80 ab | 5.78 | 0.006 |
| GSH-Px (U/mg prot) | 95.50 a | 84.04 b | 90.77 a | 1.48 | 0.001 |
| POD (U/mg prot) | 7.48 a | 4.54 b | 8.39 a | 0.55 | 0.002 |
| CAT (U/mg prot) | 14.55 a | 10.05 b | 15.40 a | 0.88 | 0.014 |
| H2O2 (mmol/g prot) | 3.77 a | 8.81 b | 4.73 a | 0.74 | 0.003 |
| MDA (nmol/mg prot) | 12.63 b | 18.58 a | 12.43 b | 1.06 | 0.012 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, H.; Rao, Z.; Li, Y.; Zhang, W.; Zhao, L.; Wang, Z.; Guo, Y. Dietary Supplementation of Novel Aflatoxin Oxidase CotA Alleviates Aflatoxin B1-Induced Oxidative Stress, Lipid Metabolism Disorder, and Apoptosis in the Liver of Japanese Quails. Animals 2025, 15, 1555. https://doi.org/10.3390/ani15111555
Lv H, Rao Z, Li Y, Zhang W, Zhao L, Wang Z, Guo Y. Dietary Supplementation of Novel Aflatoxin Oxidase CotA Alleviates Aflatoxin B1-Induced Oxidative Stress, Lipid Metabolism Disorder, and Apoptosis in the Liver of Japanese Quails. Animals. 2025; 15(11):1555. https://doi.org/10.3390/ani15111555
Chicago/Turabian StyleLv, Hao, Zhiyong Rao, Yuting Li, Wei Zhang, Lihong Zhao, Zhixiang Wang, and Yongpeng Guo. 2025. "Dietary Supplementation of Novel Aflatoxin Oxidase CotA Alleviates Aflatoxin B1-Induced Oxidative Stress, Lipid Metabolism Disorder, and Apoptosis in the Liver of Japanese Quails" Animals 15, no. 11: 1555. https://doi.org/10.3390/ani15111555
APA StyleLv, H., Rao, Z., Li, Y., Zhang, W., Zhao, L., Wang, Z., & Guo, Y. (2025). Dietary Supplementation of Novel Aflatoxin Oxidase CotA Alleviates Aflatoxin B1-Induced Oxidative Stress, Lipid Metabolism Disorder, and Apoptosis in the Liver of Japanese Quails. Animals, 15(11), 1555. https://doi.org/10.3390/ani15111555