1. Introduction
The Demodecidae (Acariformes: Prostigmata) fauna of the soricomorphs (Soricomorpha) has been poorly studied. Six species have been described in this group, and only 11 original publications are known worldwide [
1]. The majority (eight) of these records concern the Mediterranean water shrew
Neomys anomalus Cabrera, 1907, and the common shrew
Sorex araneus Linnaeus, 1758; for each of these hosts, two Demodecidae species were described [
2,
3,
4,
5,
6]. Thus far, only one species,
Demodex foveolator Bukva, 1984, has been known from the lesser white-toothed shrew
Crocidura suaveolens (Pallas, 1811), found in this host only in the tail region. It was described from the Czechia and recently recorded from Poland [
7,
8]. In contrast,
Demodex talpae Hirst, 1921, collected from the European mole
Talpa europaea Linnaeus, 1758, was described based on several specimens obtained from a single mole specimen from the United Kingdom [
9] and was mentioned in a brief report from Poland covering the parasite fauna of the mole [
10].
However, it appears that these data do not reflect the actual distribution of the Demodecidae in the Soricomorpha, a group comprising approx. 500 mammal species, several of which are widely distributed. The common shrew, Mediterranean water shrew, lesser white-toothed shrew, and the European mole mentioned above, exhibit distributions covering the entirety of Europe and the Palearctic [
11,
12]. The absence of data regarding the presence of Demodecidae in these host species likely stems from the lack of appropriate studies, as this group is rarely mentioned in comprehensive analyses of parasitofauna communities. However, the generic diversity of these mites in the soricomorphs is interesting because apart from the most commonly noted and the most species-rich
Demodex, the genera
Apodemodex and
Soricidex have been described only from Soricomorpha [
1,
2,
5,
13].
Recently, a new
Demodex species has been found in the hairy skin of the body of the lesser white-toothed shrew. In terms of morphological traits, it was found to be similar to
D. talpae, which was previously known from a highly laconic description by Hirst [
9], supplemented with an illustration. Therefore, in order to verify the status of the presently discovered demodecid from the lesser white-toothed shrew, a redescription
D. talpae was necessary.
2. Materials and Methods
Eight specimens of dead C. suaveolens from Poland (Wielkopolska Voivodeship, Słomowo, 52°21′11″ N, 17°32′41″ E) and ten T. europaea (Wielkopolska Voivodeship, Słomowo, 52°21′11″ N, 17°32′41″ E, six moles; Pomeranian Voivodeship, Pszczółki, 54°10′21″ N, 18°41′54″ E, three moles; Lublewko 54°45′28″ N, 17°55′37″ E, one mole), collected from October 2016–May 2019, were examined for demodecid mites.
Demodecidae were isolated using the digestion method developed for the detection of mammalian skin mites [
14], with modifications to suit the examined host. Skin fragments of 1 cm
2 were excised with a scalpel from several body regions, including the head (around eyes, ear pinnae, nose, area of vibrissae, lips, chin, cheeks, vertex), neck, abdomen, back, limbs, tail, and the genital–anal area. Skin samples were preserved in 70% ethanol and subjected to digestion in 10% potassium hydroxide solution until it was completely digested. The obtained samples were decanted (examination of 1 cm
2 of the skin equal to the analysis of approximately 100 wet preparations), mounted and examined using phase–contrast microscopy (Nikon Eclipse 50 i). The mites were placed in polyvinyl-lactophenol solution and measured (measurements in micrometers) as follows: total body length = length of gnathosoma, podosoma and opisthosoma; gnathosomal width = width at base; podosomal and opisthosomal width = maximum width.
The specimen depositories are cited using the following abbreviation: UGDIZP, University of Gdańsk, Department of Invertebrate Zoology and Parasitology, Gdańsk, Poland. The description of the species adopted the nomenclature commonly used for the family Demodecidae [
15] and was completed with the nomenclature proposed by Bochkov [
16] for the superfamily Cheyletoidea (Acariformes: Prostigmata) and by Izdebska and Rolbiecki [
17]. The scientific and common names of the hosts follow Wilson and Reeder [
11] and the Taxonomic Information System [
18].
To define the level of host infection, the following main parasitological parameters were measured: prevalence (percentage of hosts infected), mean intensity (mean number of parasites in infected hosts), and intensity range (minimum and maximum number of parasite individuals per host) [
19].
3. Results
3.1. Demodex crocidurae Izdebska, Cierocka, Rolbiecki, 2021
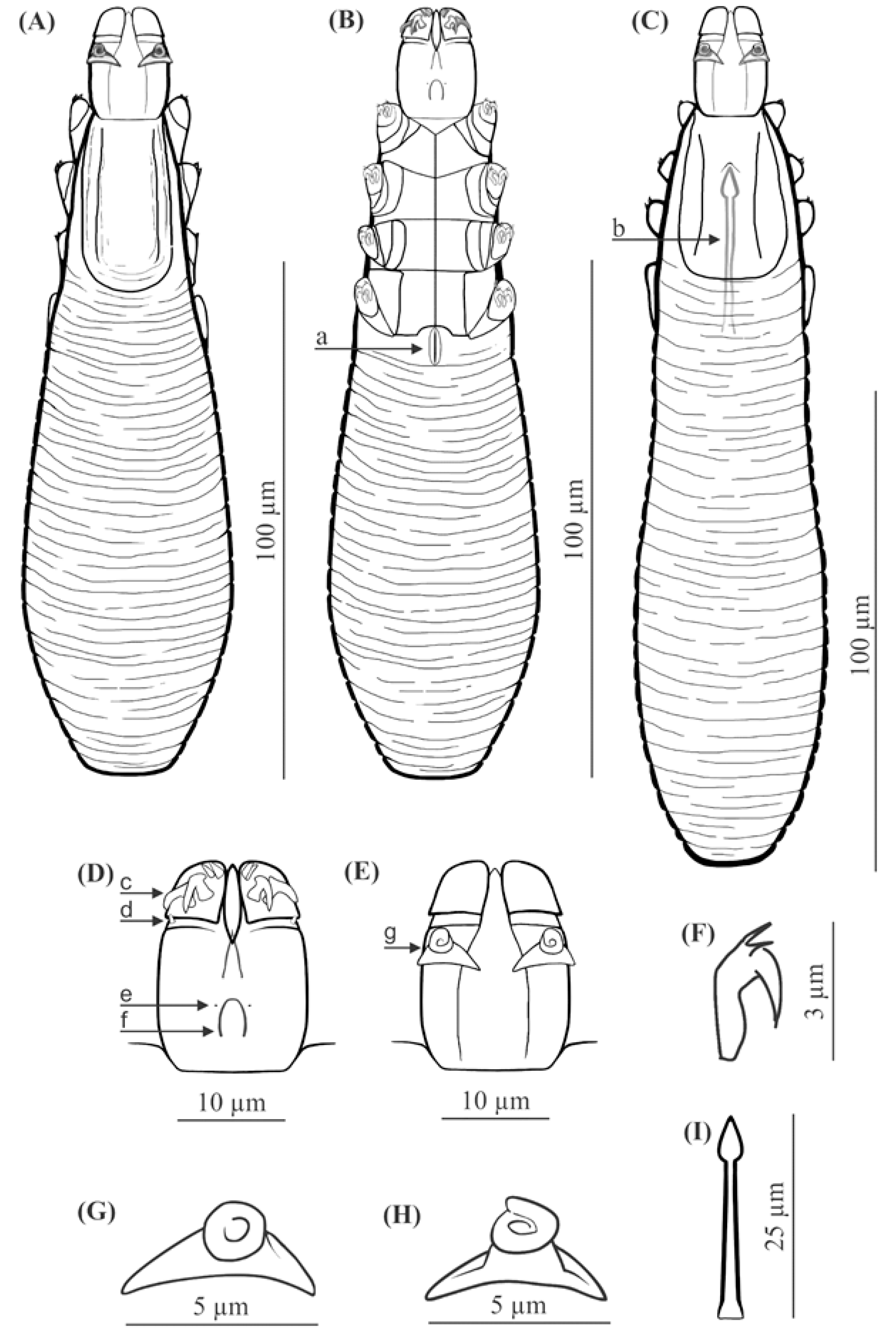
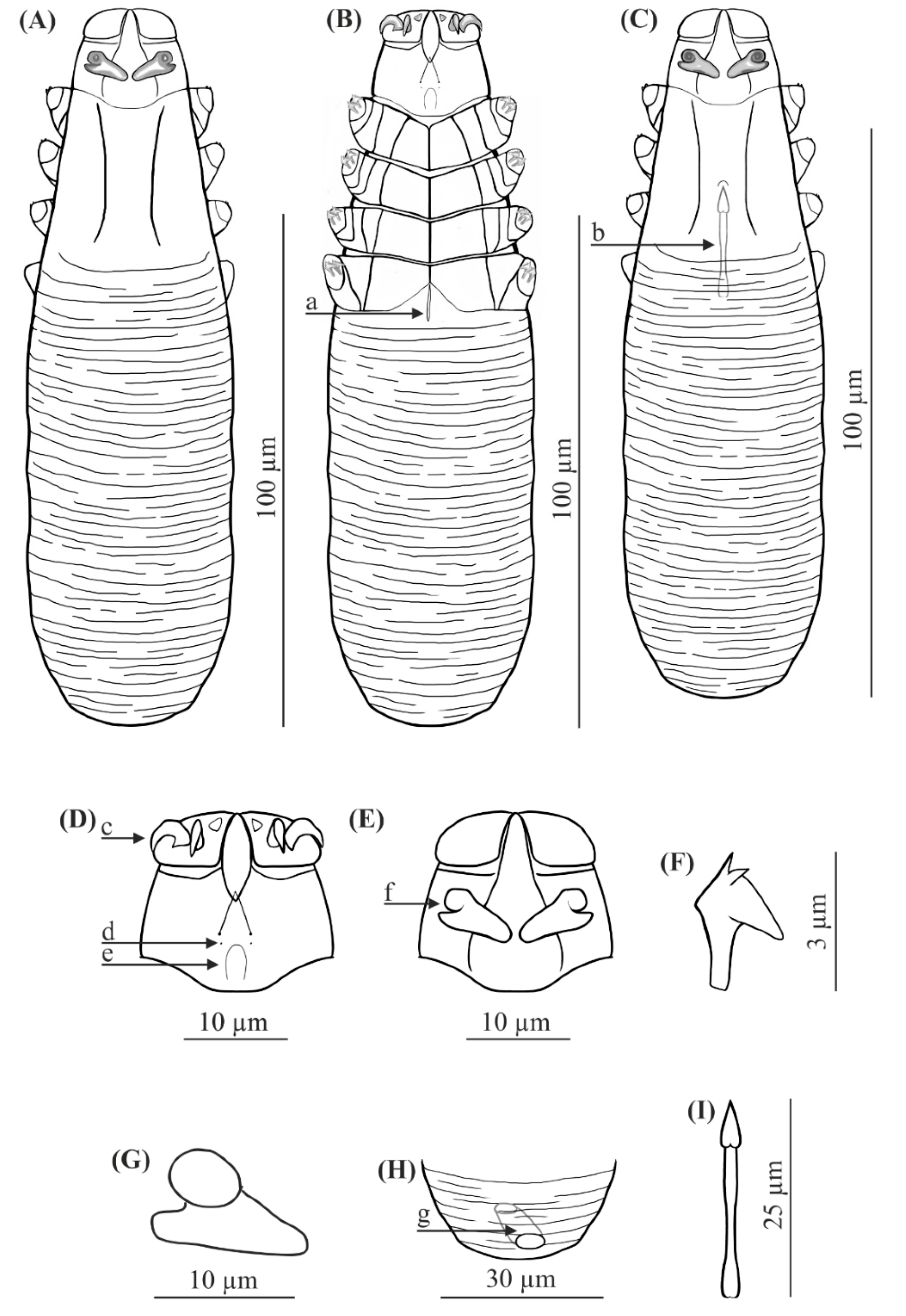
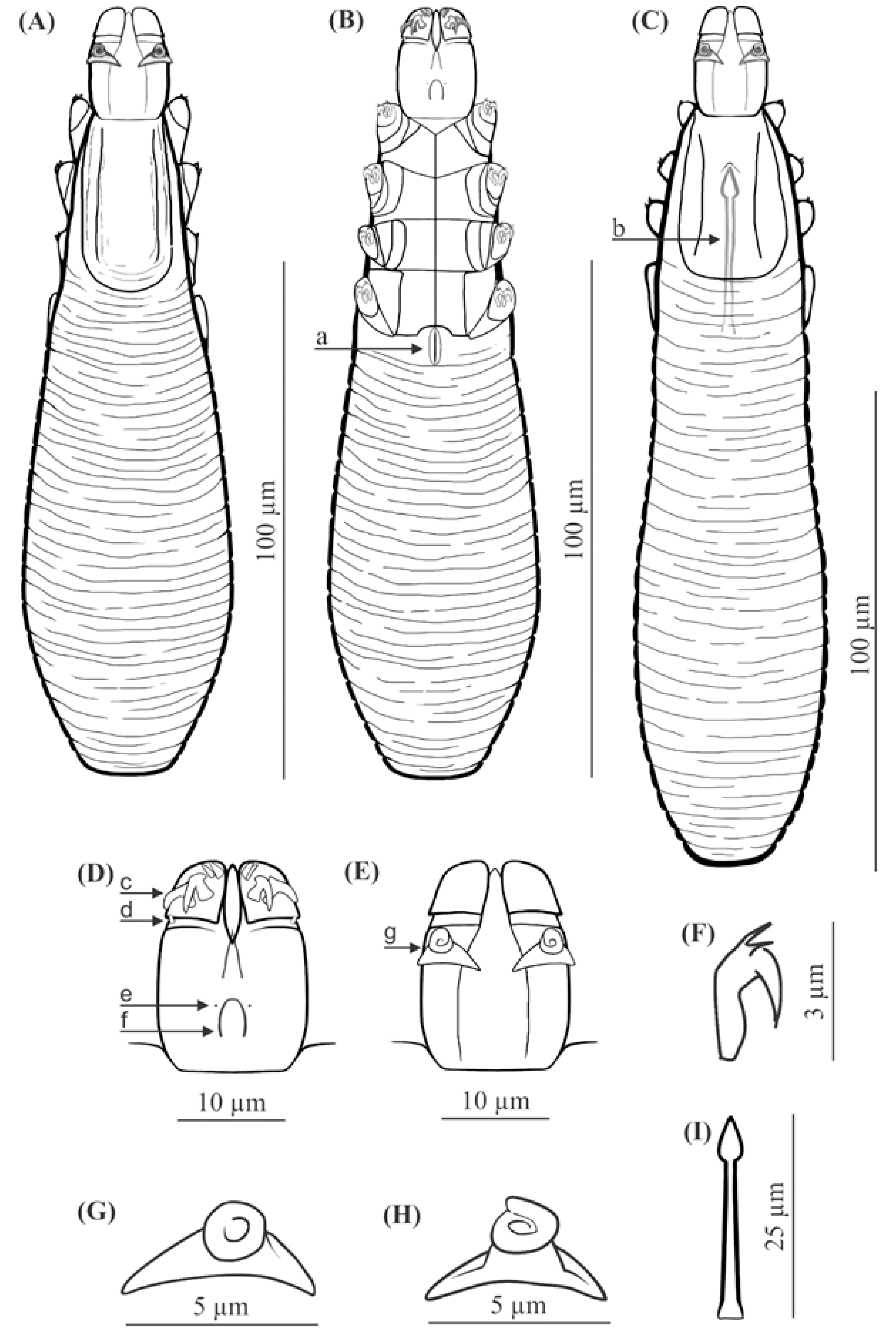
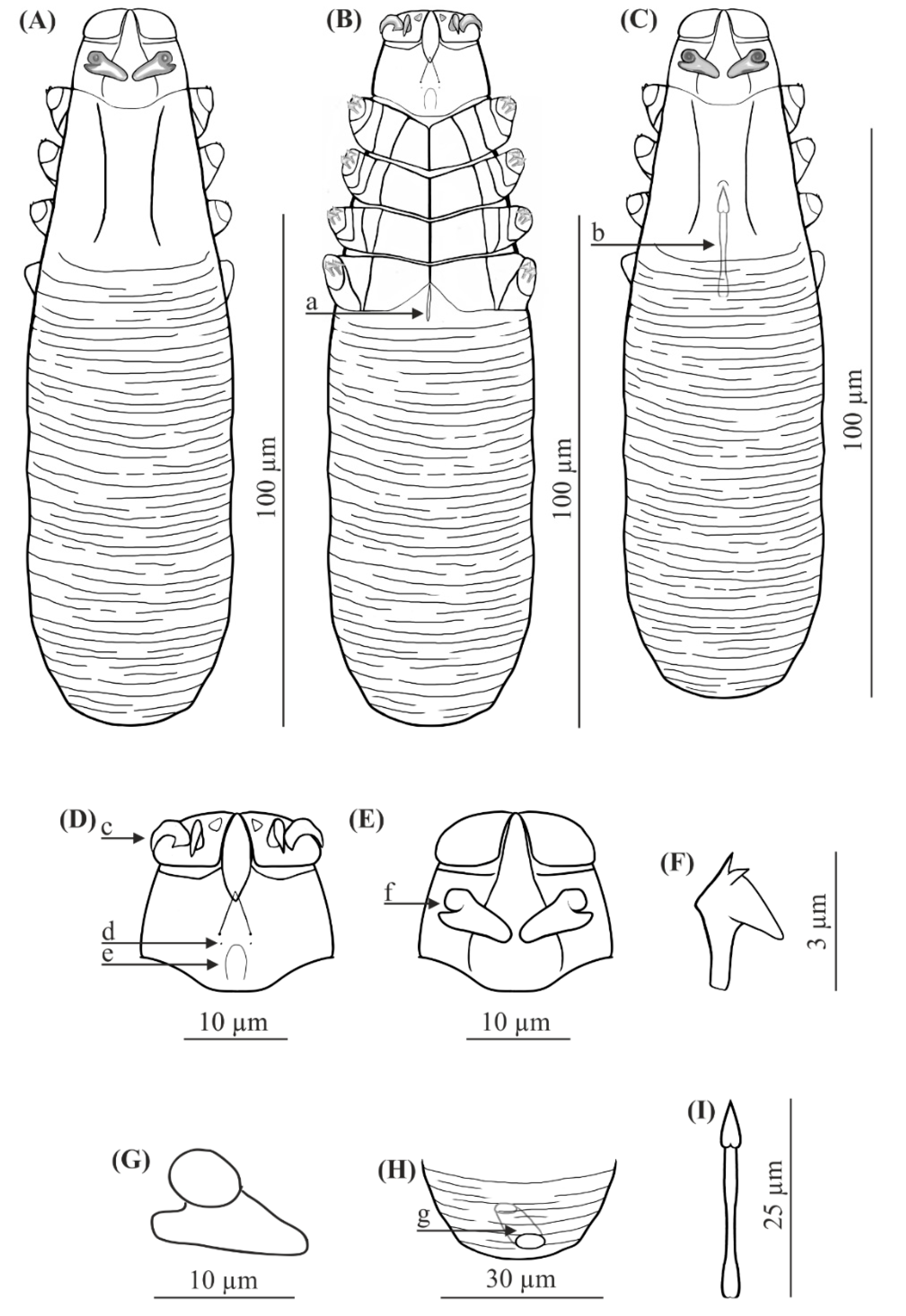
FEMALE (holotype and 60 paratypes): Body 153 (138–176) long and 29 (23–35) wide (holotype, 150 × 27). Distinctly separated gnathosoma, rectangular, longer than wide; on dorsal surface in anterior part of basal (coxal) segment, parallel to base, pair of massive boomerang-shaped supracoxal spines (setae elc.p) present, ca. 5.0 long (holotype, 5.0). Palps 3-segmented, terminating in two claw-like, large spines on tibio-tarsus, ca. 2.0 long (holotype, 2.0) and two small, conical spines; conical setae v″F near external edge of middle segment (trochanter-femur-genu) present. On ventral surface, horseshoe-shaped pharyngeal bulb with pair of conical subgnathosomal setae (setae n) situated on either side of and slightly posterior to anterior limit. Podosoma trapezoidal, widening posterior end; four pairs of short legs, with coxae integrated into ventral idiosomal wall and five free, overlapping segments (trochanter–tarsus); two forked claws, ca. 3.0 long (holotype, 3.0) with large, pointed subterminal spur, and one solenidion (ω) on each tarsus. Epimeral plates (coxal fields) distinctly sclerotized; pairs I-II trapezoidal, III-IV pairs rectangular; posterior edges of pair IV indented archwise; anterior end of vulva between incision. Podosomal shield reaching level of legs III; posterior edge of this shield is convex. Opisthosoma constitutes 57% (53−63%) of body length (holotype, 57%); widens to 3/4 of opisthosoma length and then becomes narrower. Whole opisthosoma distinctly annulated; annulation reaches level of legs III dorsally; annuli relatively wide, ca. 1.5−2.0 µm. Opisthosomal organ absent. Vulva 6 (5−10) long (holotype, 8.0), located between and behind incision of IV epimeral plate.
MALE (32 paratypes): Slightly longer and slender than female, 158 (139−182) long, 30 (24−35) wide. Distinctly separated gnathosoma, rectangular, longer than wide. Pharyngeal bulb and morphological details of gnathosoma similar to those in females. In addition, podosoma and legs shaped similar to those in females; however, posterior edge of epimeral plate IV lacks incision. Opisthosoma constitutes 61% (56−66%) of body length; opisthosoma, similar to females, distinctly annulated; annuli relatively wide, ca. 1.5−2.0 µm. Opisthosomal organ absent. Aedeagus 25 (22−30) long, on dorsal surface, located between epimeral plates II and IV. Genital opening located on dorsal surface, at level of anterior margin of epimeral plate II.
Type material: Female holotype (reg. no. UGDIZPSCSDDc13f) from Crocidura suaveolens (reg. no. MSSCs01/2017), Słomowo, Wielkopolska Voivodeship, Poland, August 2017, parasites coll. K. Cierocka, J.N. Izdebska, host coll. J.N. Izdebska, L. Rolbiecki; 60 female paratypes (reg. nos. UGDIZPSCSDDc01-12f, UGDIZPSCSDDc14-61f) and 32 male paratypes (reg. nos. UGDIZPSCSDDc01-32m) from Crocidura suaveolens (reg. nos. MSSCs01/2017, MSSCs02/2018, MSSCs03-05/2017, MSSCs06-08/2018), Słomowo, Wielkopolska Voivodeship, Poland, August 2017, August 2018, same collectors.
Type material deposition: Whole type material (mounted microscope slides with the demodecid mites) is deposited in scientific collections within the framework of the Collection of Extant Invertebrates in Department of Invertebrate Zoology and Parasitology, University of Gdańsk, Poland.
Infection and location in the host. Demodex crocidurae sp. nov. was found in all examined lesser white-toothed shrews (100%), with a mean intensity of 11.7 and intensity range of 3–26 individuals per host: 93 individuals in total (32 males, 61 females). The demodecid mites were found on the hairy skin of the body (head—31 individuals, abdomen—30, back—28, and genital-anal area—4). The observed mites did not cause any lesions in examined shrews.
Etymology. The specific epithet crocidurae refers to the specific name of the host.
3.2. Demodex talpae Hirst, 1921
FEMALE (n = 8): Body 143 (134–154) long and 38 (35–43) wide. Distinctly separated gnathosoma, trapezoidal, base width longer than length; on dorsal surface in anterior part of basal (coxal) segment, directed posteromedially, pair of massive, wedge-shaped supracoxal spines (setae elc.p) present, ca. 5.5 long. Palps 3-segmented, terminating in two claw-like, large spines on tibio-tarsus, ca. 3.0 long and one small, conical spine. On ventral surface, horseshoe-shaped pharyngeal bulb with pair of conical subgnathosomal setae (setae n) situated exactly at level of anterior margin on both sides. Podosoma trapezoidal, widening posterior end; four pairs of short legs, with coxae integrated into ventral idiosomal wall and five free, overlapping segments (trochanter–tarsus); two forked claws, ca. 3.0 long with large, pointed subterminal spur. Epimeral plates (coxal fields) distinctly sclerotized; I-IV pairs trapezoidal; posterior edges of pair IV weakly sclerotized, form triangular incision, which almost surrounding vulva. Podosomal shield reaching level of legs III; posterior edge of this shield is concave. Opisthosoma constitutes 57% (53−60%) of body length; widens towards end; widest at end. Opisthosoma distinctly and densely annulated; annulation also reaches dorsal podosoma side (level of legs III); annuli relatively wide at ca. 1.0–1.5 µm. Opisthosomal organ present, tubular; opisthosomal pore oval, 1.5–2.0 in diameter. Vulva 9 (8−10) long, located between and behind incision of IV epimeral plate.
MALE (n = 6): Shorter than female, 128 (118−150) long, 34 (33−40) wide. Distinctly separated gnathosoma, trapezoidal, base width longer than length. Pharyngeal bulb and morphological details of gnathosoma similar to those in female. Shape of podosoma and legs similar to those in female; however, only posterior edge of epimeral plate IV without incision. Opisthosoma constitutes 58% (55−60%) of body length; widens towards posterior end. Opisthosoma, similar to females, distinctly annulated; annuli relatively wide, ca. 1.0−1.5 µm. Opisthosomal organ present, similar to that in female. Aedeagus 19 (18−22) long, on dorsal surface, located between epimeral plates III and IV. Genital opening located on dorsal surface, on border between epimeral plates II and III.
Material deposition: Mounted microscope slides with specimens were stored in scientific collections within the framework of the Collection of Extant Invertebrates in Department of Invertebrate Zoology and Parasitology, University of Gdańsk, Poland. Eight females (reg. no. UGDIZPTTeDDt01-08f) and six males (reg. no. UGDIZPTTeDDt01-06m) from Talpa europaea (reg. no. MSTTe01/2016, MSTTe02/2019, MSTTe03/2018), Lublewko, Pomeranian Voivodeship, Słomowo, Wielkopolska Voivodeship, Pszczółki, Pomeranian Voivodeship, Poland, October 2016, August 2018, May 2019, parasites coll. K. Cierocka, J.N. Izdebska, host coll. J.N. Izdebska, L. Rolbiecki.
Infection and location in the host. Demodex talpae was noted in 30.0% of the ten examined European moles, with a mean intensity of 4.7 and intensity range of 2.0–8.0 individuals per host: 14 individuals in total (six males, eight females). Mites were found in the hairy skin of the body (back—six individuals; abdomen—eight). The observed mites did not cause any lesions in the examined European moles. Infestations were not associated with skin lesions or other symptoms.
3.3. Differential Diagnosis
Regarding the known Demodecidae,
D. crocidurae sp. nov. appears to be closely related to
D. talpae described from the European mole, this being a representative of another family within the order Soricomorpha. However,
D. talpae is smaller and has different body proportions; it is relatively wider, with shorter opisthosoma; in addition,
D. talpae males and females have similar body size and proportions, whereas
D. crocidurae males are typically slightly longer (with longer opisthosoma) than the females (
Table 1,
Table 2 and
Table 3,
Figure 1,
Figure 2 and
Figure 3).
The gnathosoma in D. crocidurae is rectangular, narrow, and longer than wide; in D. talpae, it is trapezoidal, shorter than wide at the base. The supracoxal spines in both species are large and massive (ca. 5.0 um in length); however, they are boomerang shaped in D. crocidurae and wedge-shaped in D. talpae. Additionally, they are directed parallel to the base of the gnathosoma in D. crocidurae and are directed posteromedially in D. talpae. In addition, there is a process on the supracoxal spines; this is located in the center of the spine in D. crocidurae and at the widest part in D. talpae. The terminal segments of the palpi are equipped with two large and two small spines in D. cocidurae, while there are two large and one small spines in D. talpae. The subgnathosomal setae are located on either side of and slightly posterior to the anterior margin of the pharyngeal bulb in D. crocidurae, and exactly at the level of anterior margin of the pharyngeal bulb in D. talpae. The leg tarsi are equipped with forked claws differing in shape between both demodecid species. Differences also exist between the epimeral plates: in D. crocidurae, I–II pairs are trapezoidal and III–IV are rectangular, while in D. talpae, all epimeral plates are trapezoidal. Moreover, the posterior edge of epimeral plates IV has an arched shape in D. crocidurae females but a triangular shape in D. talpae females. In addition, the posterior edge of the podosomal shield is convex in D. crocidurae and concave in D. talpae. The opisthosoma in D. crocidurae is narrower and longer, widens to 3/4 of its length and then narrows, while in D. talpae it is wider and shorter, widening posteriorly, with the widest part at the end of the body. In addition, the opisthosomal organ is absent in D. crocidurae, but it is present in both sexes in D. talpae. Furthermore, in D. crocidurae males, the aedeagus is longer (22–30 μm in length) and located at epimeral plates II–IV with the genital opening at the level of anterior margin of epimeral plates II; in D. talpae males, the aedeagus is shorter (18–22 μm in length) and located at the border between epimeral plates III and IV, with the genital opening on the border between epimeral plates II and III.
4. Discussion
Demodex crocidurae sp. nov. clearly differs from all known Demodecidae species. However, it resembles
D. talpae in general body shape, and certain characteristics, namely, the shape of the supracoxal spines and leg claws, which are also observed in several demodecid mites of the Muridae, e.g.,
D. musculi Oudemans, 1897,
D. apodemi Hirst, 1918,
D. corniculatus Izdebska, 2012,
D. bandicotae Izdebska, Rolbiecki, Morand & Ribas, 2017 [
20,
21,
22,
23]. Although both the two host species, under discussion, belongs to the order Soricomorpha, they are representatives of two separate families, Soricidae and Talpidae, respectively with a doubtful level of affinity [
11]. In addition, the lesser white-toothed shrew and the European mole have different environmental preferences and different biologies.
Crocidura suaveolens is a small mammal (weight in the 3–7 g range), found in Central Europe, Israel, Saudi Arabia, Middle East, Caucasus, Kyrgyzstan, north-east China and Korea, where it primarily inhabits bushes, forest communities, as well as parks and gardens and buildings [
24]. It forms a nest among grass or in abandoned rodent burrows. It is active throughout the year and day and night and has short activity and rest cycles. Conversely,
T. europaea is considerably larger (up to 120 g) and has a considerably wider distribution range, including the Northern Hemisphere, being found in Europe, Asia and North America. It is also active throughout the year during both day and night, but it has longer activity and rest cycles. The mole is also a subterranean mammal, constructing underground nests and tunnels [
12,
25]. Thus, it is difficult to expect that the same Demodecidae species will be found in both host species.
Demodex crocidurae and
D. talpae, although morphologically similar (
Table 1,
Table 2 and
Table 3,
Figure 1,
Figure 2 and
Figure 3) and found in comparable microhabitats on their hosts (hairy skin), were found to demonstrate different infestation levels.
Demodex crocidurae was common in all examined shrews, despite originating from different populations and seasons. In contrast,
D. talpae, demonstrates low infestation parameters; only 14 demodecid mites were recorded in only three mole specimens in the present study (30.0% of the examined).
Previously,
D. talpae was only known from a single record from the United Kingdom, in which Hirst [
9] described the species based on several specimens obtained from a single mole in 1919. However,
D. talpae was recorded from several hosts in recent studies on the mole parasitofauna conducted in Poland [
10]; but, due to the different study methodology (determined by parasite analyses from different groups and locations), and poor preservation state of the specimens, it was not possible to use all specimens for the redescription.
Knowledge of the distribution of Demodex species is still fragmentary the distribution of both species may be wider than presently known. In the present study, D. crocidurae was found in lesser white-toothed shrews from an area located near the northwestern boundary of their distribution in Europe; the shrew is found relatively rarely in this area, and considering its solitary behavior (apart from the breeding season), this likely does not facilitate the spread of this parasite. Despite this, D. crocidurae was well represented on all individuals of all the host populations. The considerably lower D. talpae infestation level observed in European mole is probably linked to its subterranean and extremely solitary behavior. However, its records from distant localities (the United Kingdom and Poland) over the period of 100 years clearly suggests that it may be present in more areas of its host distribution.







{kind=link}
{kind=link}
{kind=link}