Phenotypic Characterization of Milk Yield and Quality Traits in a Large Population of Water Buffaloes

 ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data
2.2. Statistical Analyses
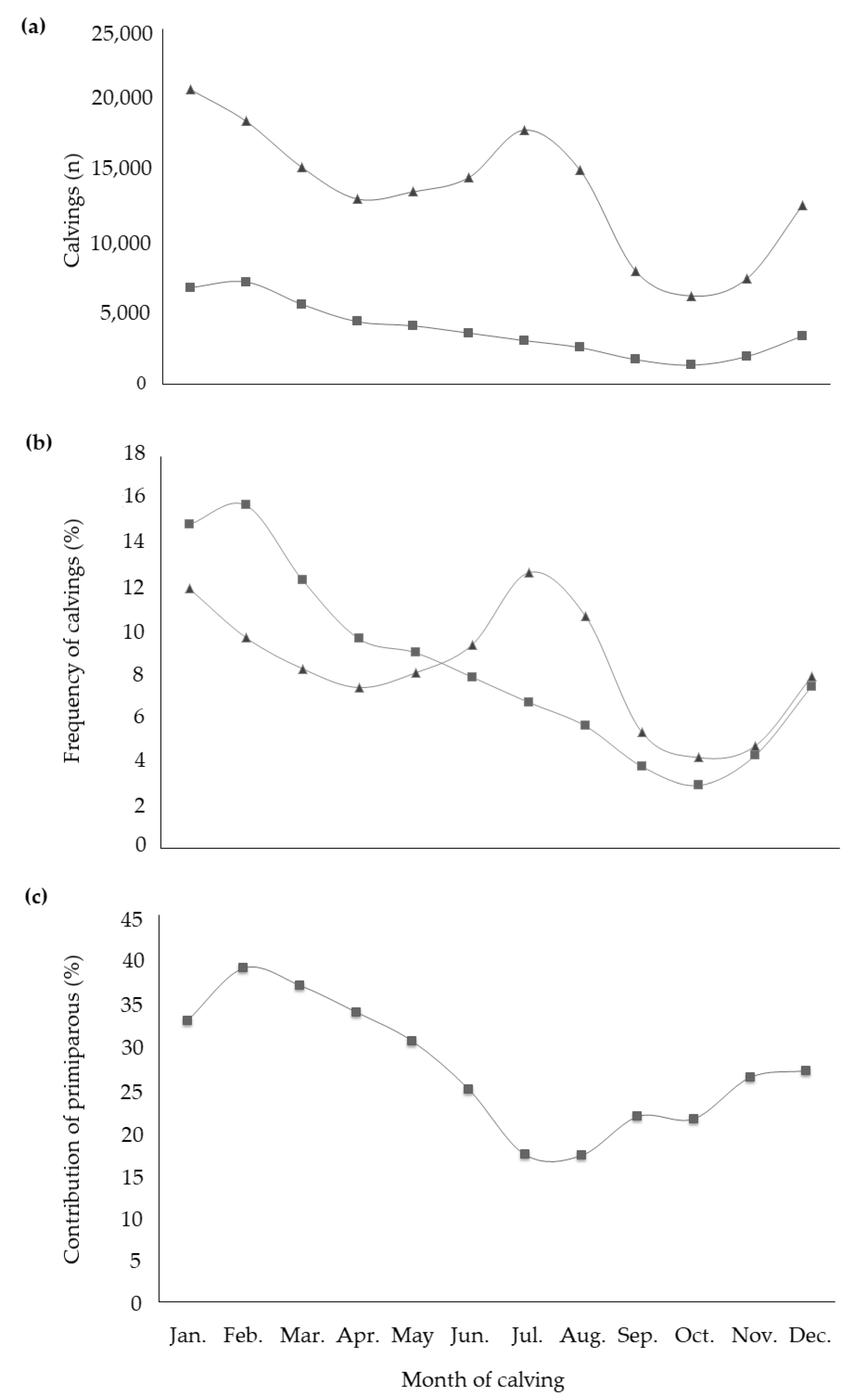
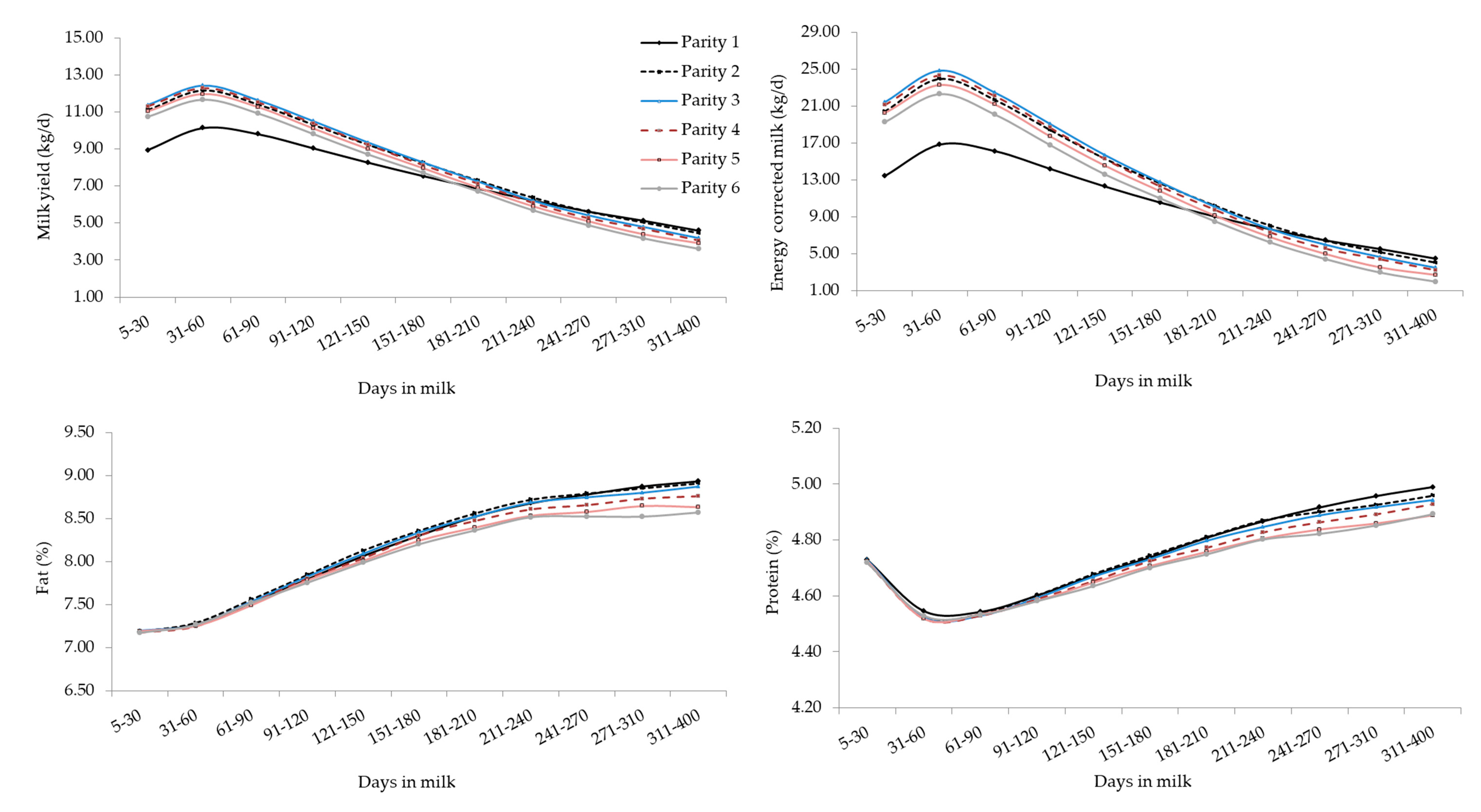
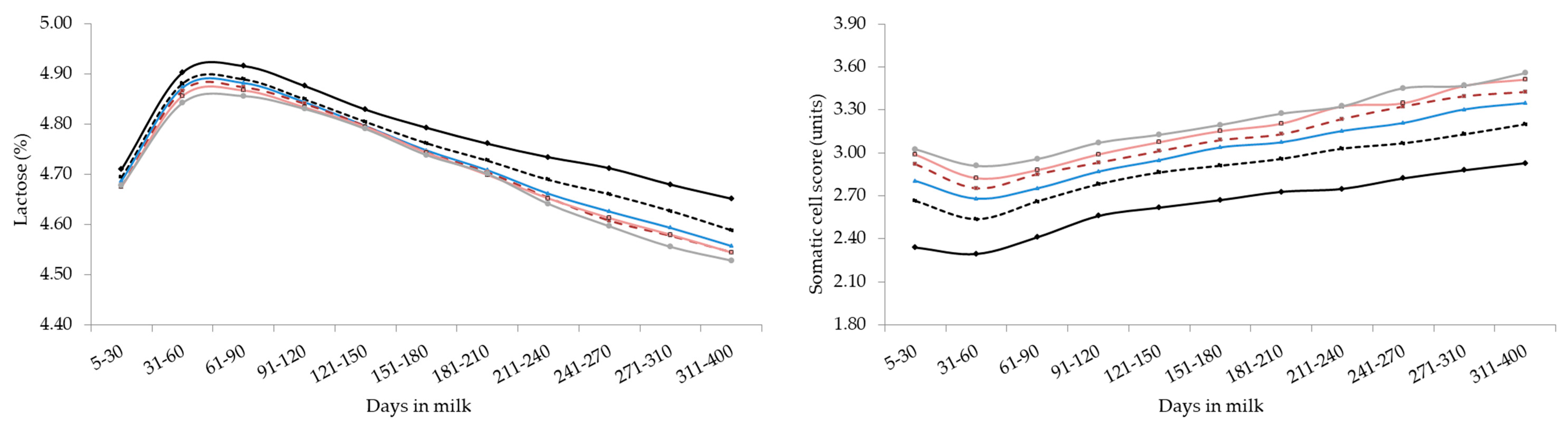
3. Results and Discussion
3.1. Descriptive Statistics
3.2. Correlations
3.3. Analysis of Variance
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Borghese, A.; Moioli, B. Husbandry of Dairy Animals. Buffalo: Mediterranean Region. In Encyclopedia of Dairy Science; Elsevier Science Ltd.: London, UK, 2011; p. 784. [Google Scholar] [CrossRef]
- Fox, P.F.; Uniacke-Lowe, T.; McSweeney, P.L.H.; O’Mahony, J.A. Dairy Chemistry and Biochemistry; Springer International Publishing: Basel, Switzerland, 2015. [Google Scholar]
- Associazione Nazionale Allevatori Specie Bufalina (ANASB). Available online: http://www.anasb.it/home1.htm.2018 (accessed on 20 June 2018).
- CLAL. Production of Mozzarella di Bufala Campana PDO, Italy. 2019. Available online: https://www.clal.it/en/?section=mozzarella_bufala_campana (accessed on 23 June 2019).
- Italian Breeders Association (Associazione Italiana Allevatori, AIA). Bollettino online. Statistiche ufficiali. 2018. Available online: http://bollettino.aia.it/Contenuti.aspx?CD_GruppoStampe=RS&CD_Specie=C4 (accessed on 30 September 2019).
- Di Palo, R. Produzione lattea nella bufala con diete tradizionali e con l’impiego di acidi grassi. Ph.D. Thesis, University of Naples, Naples, Italy, 1992. [Google Scholar]
- Gilmour, A.R.; Gogel, B.J.; Cullis, B.R.; Welham, S.J.; Thompson, R. ASReml user guide. Release 4.1. VSN International Ltd.: Hemel Hempstead, UK, 2015. Available online: https://asreml.kb.vsni.co.uk/wp-content/uploads/sites/3/2018/02/ASReml-4.1-Functional-Specification.pdf (accessed on 5 March 2019).
- El-Salam, M.H.A.; El-Shibiny, S. A comprehensive review on the composition and properties of buffalo milk. Dairy Sci. Technol. 2011, 91, 663–699. [Google Scholar] [CrossRef]
- Manuelian, C.L.; Visentin, G.; Boselli, C.; Giangolini, G.; Cassandro, M.; De Marchi, M. Prediction of milk coagulation and acidity traits in Mediterranean buffalo milk using Fourier-transform mid-infrared spectroscopy. J. Dairy Sci. 2017, 100, 7083–7087. [Google Scholar] [CrossRef] [PubMed]
- Tripaldi, C.; Palocci, G.; Miarelli, M.; Catta, M.; Orlandini, S.; Amatiste, S.; Di Bernardini, R.; Catillo, G. Effects of mastitis on buffalo milk quality. Asian-Aust. J. Anim. Sci. 2010, 23, 1319–1324. [Google Scholar] [CrossRef]
- Sharif, A.; Ahmad, T.; Bilal, M.Q.; Yousaf, A.; Muhammad, G. Effect of severity of sub-clinical mastitis on somatic cell count and lactose contents of buffalo milk. Pakistan Vet. J. 2007, 27, 142–144. Available online: http://www.pvj.com.pk/pdf-files/27_3/page%20142-144.pdf (accessed on 10 September 2019).
- Moroni, P.; Sgoifo Rossi, C.; Pisoni, G.; Bronzo, V.; Castiglioni, B.; Boettcher, P.J. Relationships between somatic cell count and intramammary infection in buffaloes. J. Dairy Sci. 2006, 89, 998–1003. [Google Scholar] [CrossRef]
- Niero, G.; Currò, S.; Costa, A.; Penasa, M.; Cassandro, M.; Boselli, C.; Giangolini, G.; De Marchi, M. Short communication: Phenotypic characterization of total antioxidant activity of buffalo, goat, and sheep milk. J. Dairy Sci. 2018, 101, 4864–4868. [Google Scholar] [CrossRef]
- Weaver, B.; Wuensch, K.L. SPSS and SAS programs for comparing Pearson correlations and OLS regression coefficients. Behav. Res. 2013, 45, 880–895. [Google Scholar] [CrossRef]
- Aspilcueta-Borquis, R.R.; Araujo Neto, F.R.; Baldi, F.; Bignardi, A.B.; Albuquerque, L.G.; Tonhati, H. Genetic parameters for buffalo milk yield and milk quality traits using Bayesian inference. J. Dairy Sci. 2010, 93, 2195–2201. [Google Scholar] [CrossRef]
- Costa, A.; Lopez-Villalobos, N.; Visentin, G.; De Marchi, M.; Cassandro, M.; Penasa, M. Heritability and repeatability of milk lactose and its relationships with traditional milk traits, somatic cell score and freezing point in Holstein cows. Animal 2019, 13, 909–916. [Google Scholar] [CrossRef]
- Costa, A.; Egger-Danner, C.; Mészáros, G.; Fuerst, C.; Penasa, M.; Sölkner, J.; Fuerst-Waltl, B. Genetic associations of lactose and its ratios to other milk solids with health traits in Austrian Fleckvieh cows. J. Dairy Sci. 2019, 102, 4238–4248. [Google Scholar] [CrossRef]
- Cerón-Muñoz, M.; Tonhati, H.; Duarte, J.; Oliveira, J.; Muñoz-Berrocal, M.; Jurado-Gámez, H. Factors affecting somatic cell counts and their relations with milk and milk constituent yield in buffaloes. J. Dairy Sci. 2002, 85, 2885–2889. [Google Scholar] [CrossRef]
- Costa, A.; Lopez-Villalobos, N.; Sneddon, N.W.; Shalloo, L.; Franzoi, M.; De Marchi, M.; Penasa, M. Invited review: Milk lactose—Current status and future challenges in dairy cattle. J. Dairy Sci. 2019, 102, 5883–5898. [Google Scholar] [CrossRef] [PubMed]
- Miglior, F.; Sewalem, A.; Jamrozik, J.; Lefebvre, D.M.; Moore, R.K. Analysis of milk urea nitrogen and lactose and their effect on longevity in Canadian dairy cattle. J. Dairy Sci. 2006, 89, 4886–4894. [Google Scholar] [CrossRef]
- Haile-Mariam, M.; Pryce, J.E. Genetic parameters for lactose and its correlation with other milk production traits and fitness traits in pasture-based production systems. J. Dairy Sci. 2017, 100, 3754–3766. [Google Scholar] [CrossRef] [PubMed]
- Macciotta, N.P.P.; Cecchinato, A.; Mele, M.; Bittante, G. Use of multivariate factor analysis to define new indicator variables for milk composition and coagulation properties in Brown Swiss cows. J. Dairy Sci. 2012, 95, 7346–7354. [Google Scholar] [CrossRef]
- Campanile, G.; Neglia, G.; D’Occhio, M.J. Embryonic and fetal mortality in river buffalo (Bubalus bubalis). Theriogenology 2016, 86, 207–213. [Google Scholar] [CrossRef] [PubMed]
- D’Occhio, M.J.; Ghuman, S.S.; Neglia, G.; Della Valle, G.; Baruselli, P.S.; Zicarelli, L.; Visintin, J.A.; Sarkar, M.; Campanile, G. Exogenous and endogenous factors in seasonality of reproduction in buffaloes: A review. Theriogenology 2020, in press. [Google Scholar] [CrossRef]
- Zicarelli, L. Enhancing reproductive performance in domestic dairy water buffalo (Bubalus bubalis). Soc. Reprod. Fertil. 2010, 67, 443–455. [Google Scholar] [CrossRef]
- Campanile, G.; Baruselli, P.S.; Neglia, G.; Vecchio, D.; Gasparrini, B.; Gimenes, L.U.; Zicarelli, L.; D’Occhio, M.J. Ovarian function in the buffalo and implications for embryo development and assisted reproduction. Anim. Reprod. Sci. 2010, 121, 1–11. [Google Scholar] [CrossRef]
- Neglia, G.; De Nicola, D.; Esposito, L.; D’Occhio, M.J.; Fatone, G. Reproductive management in buffalo by artificial insemination. Theriogenology 2020, in press. [Google Scholar] [CrossRef]
- Presicce, G.A. Reproduction in Water Buffalo. Reprod. Dom. Anim. 2007, 42, 24–32. [Google Scholar] [CrossRef]
- Araújo, K.B.S.; Rangel, A.H.N.; Fonseca, F.C.E.; Aguiar, E.M.; Simplício, A.A.; Novaes, L.P.; Lima Júnior, D.M. Influence of the year and calving season on production, composition and mozzarella cheese yield of water buffalo in the State of Rio Grande Do Norte, Brazil. Ital. J. Anim. Sci. 2012, 11, 87–91. [Google Scholar] [CrossRef]
- Nagasaku, E.M.; Toledo, L.M.; Baruselli, P.S.; Carvalho, N.A.T. Effects of the season and number of calving on buffaloes milk production. Ital. J. Anim. Sci. 2007, 6, 1287–1289. [Google Scholar] [CrossRef]
- Pasquini, M.; Osimani, A.; Tavoletti, S.; Moreno, I.; Clementi, F.; Trombetta, M.F. Trends in the quality and hygiene parameters of bulk Italian Mediterranean buffalo (Bubalus bubalis) milk: A three year study. Animal Sci. J. 2018, 89, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Bartocci, S.; Tripaldi, C.; Terramoccia, S. Characteristics of foodstuffs and diets, and the quanti-qualitative milk parameters of Mediterranean buffaloes bred in Italy using the intensive system: An estimate of the nutritional requirements of buffalo herds lactating or dry. Livest. Prod. Sci. 2002, 77, 45–48. [Google Scholar] [CrossRef]
- Zotos, A.; Bampidis, V. Short Communication: Milk fat quality of Greek buffalo (Bubalus bubalis). J. Food Compos. Anal. 2014, 33, 181–186. [Google Scholar] [CrossRef]
- Catillo, G.; Macciotta, N.P.P.; Carretta, A.; Cappio-Borlino, A. Effects of age and calving season on lactation curves of milk production traits in Italian water buffaloes. J. Dairy Sci. 2002, 85, 1298–1306. [Google Scholar] [CrossRef]
- Cole, J.B.; Null, D.J.; De Vries, A. Short Communication: Best prediction of 305-day lactation yields with regional and seasonal effects. J. Dairy Sci. 2011, 94, 1601–1604. [Google Scholar] [CrossRef]
- Herve, L.; Lollivier, V.; Quesnel, H.; Boutinaud, M. Oxytocin induces mammary epithelium disruption and could stimulate epithelial cell exfoliation. J. Mammary Gland Biol. Neoplasia. 2018, 23, 139–147. [Google Scholar] [CrossRef]
- Dürr, J.W.; Cue, R.I.; Monardes, H.G.; Moro-Méndez, J.; Wade, K.M. Milk losses associated with somatic cell counts per breed, parity and stage of lactation in Canadian dairy cattle. Livest Sci. 2008, 117, 225–232. [Google Scholar] [CrossRef]
- Herve, L.; Quesnel, H.; Veron, M.; Portanguen, J.; Gross, J.J.; Bruckmaier, R.M.; Boutinaud, M. Milk yield loss in response to feed restriction is associated with mammary epithelial cell exfoliation in dairy cows. J. Dairy Sci. 2019, 102, 2670–2685. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Trait | Mean | CV | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2013 | 2014 | 2015 | 2016 | 2017 | 2013 | 2014 | 2015 | 2016 | 2017 | |
| Milk yield (kg/d) | 8.81 | 8.78 | 8.97 | 9.24 | 9.20 | 38.85 | 39.58 | 39.07 | 38.53 | 39.41 |
| ECM (kg/d) | 14.48 | 14.36 | 14.83 | 15.75 | 15.73 | 67.83 | 70.30 | 69.07 | 67.09 | 67.06 |
| Fat (%) | 8.18 | 8.00 | 8.01 | 8.14 | 8.26 | 20.64 | 20.87 | 20.18 | 19.85 | 20.03 |
| Protein (%) | 4.69 | 4.74 | 4.67 | 4.69 | 4.73 | 9.42 | 9.60 | 9.61 | 9.14 | 8.44 |
| Lactose (%) | 4.79 | 4.78 | 4.78 | 4.75 | 4.72 | 6.54 | 6.93 | 6.82 | 6.56 | 6.29 |
| SCS (units) | 2.71 | 3.02 | 2.92 | 2.94 | 3.11 | 72.59 | 59.44 | 56.70 | 56.83 | 55.04 |
| Table | Milk Yield | ECM | Fat | Protein | Lactose | SCS |
|---|---|---|---|---|---|---|
| Milk yield | - | 0.96 | −0.27 | −0.26 | 0.23 | −0.12 |
| ECM | 0.96 | - | −0.09 | −0.15 | 0.14 | −0.09 |
| Fat | −0.26 | −0.08 | - | 0.43 | −0.36 | 0.12 |
| Protein | −0.24 | −0.14 | 0.39 | - | −0.41 | 0.06 |
| Lactose | 0.25 | 0.16 | −0.35 | −0.40 | - | −0.28 |
| SCS | −0.14 | −0.12 | 0.10 | 0.02 | −0.30 | - |
| Month of Calving | Milk Yield | SE | ECM | SE | Fat | SE | Protein | SE | Lactose | SE | SCS | SE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| January | 8.116 | 0.017 | 12.817 | 0.050 | 8.155 | 0.010 | 4.728 | 0.003 | 4.731 | 0.002 | 3.004 | 0.011 |
| February | 8.159 | 0.017 | 12.898 | 0.052 | 8.152 | 0.010 | 4.728 | 0.003 | 4.734 | 0.002 | 2.992 | 0.012 |
| March | 8.190 | 0.017 | 12.928 | 0.053 | 8.114 | 0.010 | 4.726 | 0.003 | 4.735 | 0.002 | 2.997 | 0.012 |
| April | 8.124 | 0.018 | 12.744 | 0.055 | 8.118 | 0.011 | 4.731 | 0.003 | 4.733 | 0.002 | 2.998 | 0.012 |
| May | 8.009 | 0.018 | 12.405 | 0.054 | 8.125 | 0.011 | 4.737 | 0.003 | 4.733 | 0.002 | 3.012 | 0.012 |
| June | 7.904 | 0.018 | 12.100 | 0.053 | 8.114 | 0.010 | 4.740 | 0.003 | 4.728 | 0.002 | 3.013 | 0.012 |
| July | 7.805 | 0.017 | 11.817 | 0.051 | 8.118 | 0.010 | 4.745 | 0.003 | 4.725 | 0.002 | 3.013 | 0.012 |
| August | 7.838 | 0.017 | 11.966 | 0.053 | 8.108 | 0.010 | 4.735 | 0.003 | 4.736 | 0.002 | 2.997 | 0.012 |
| September | 7.806 | 0.020 | 11.843 | 0.063 | 8.120 | 0.012 | 4.739 | 0.003 | 4.730 | 0.002 | 3.007 | 0.014 |
| October | 7.849 | 0.022 | 12.027 | 0.067 | 8.130 | 0.013 | 4.735 | 0.004 | 4.729 | 0.003 | 3.003 | 0.014 |
| November | 7.943 | 0.021 | 12.288 | 0.065 | 8.130 | 0.012 | 4.735 | 0.003 | 4.721 | 0.002 | 2.988 | 0.014 |
| December | 8.084 | 0.018 | 12.760 | 0.056 | 8.146 | 0.011 | 4.732 | 0.003 | 4.726 | 0.002 | 3.000 | 0.012 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, A.; Negrini, R.; De Marchi, M.; Campanile, G.; Neglia, G. Phenotypic Characterization of Milk Yield and Quality Traits in a Large Population of Water Buffaloes. Animals 2020, 10, 327. https://doi.org/10.3390/ani10020327
Costa A, Negrini R, De Marchi M, Campanile G, Neglia G. Phenotypic Characterization of Milk Yield and Quality Traits in a Large Population of Water Buffaloes. Animals. 2020; 10(2):327. https://doi.org/10.3390/ani10020327
Chicago/Turabian StyleCosta, Angela, Riccardo Negrini, Massimo De Marchi, Giuseppe Campanile, and Gianluca Neglia. 2020. "Phenotypic Characterization of Milk Yield and Quality Traits in a Large Population of Water Buffaloes" Animals 10, no. 2: 327. https://doi.org/10.3390/ani10020327
APA StyleCosta, A., Negrini, R., De Marchi, M., Campanile, G., & Neglia, G. (2020). Phenotypic Characterization of Milk Yield and Quality Traits in a Large Population of Water Buffaloes. Animals, 10(2), 327. https://doi.org/10.3390/ani10020327