Combined Use of Lachancea thermotolerans and Schizosaccharomyces pombe in Winemaking: A Review


Abstract

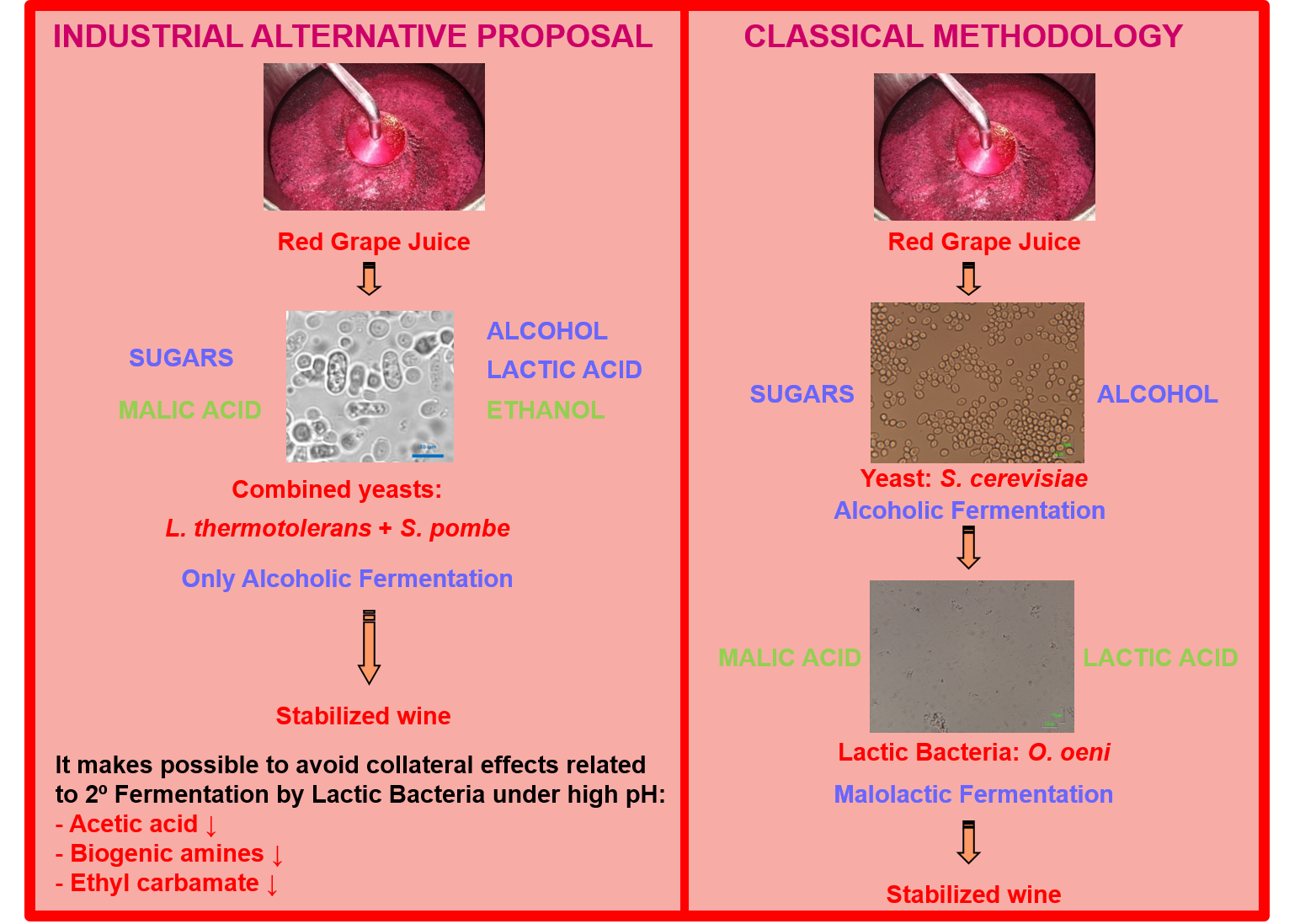
1. Introduction
2. The Influence of L. thermotolerans and S. pombe on the Main Wine Quality Parameters
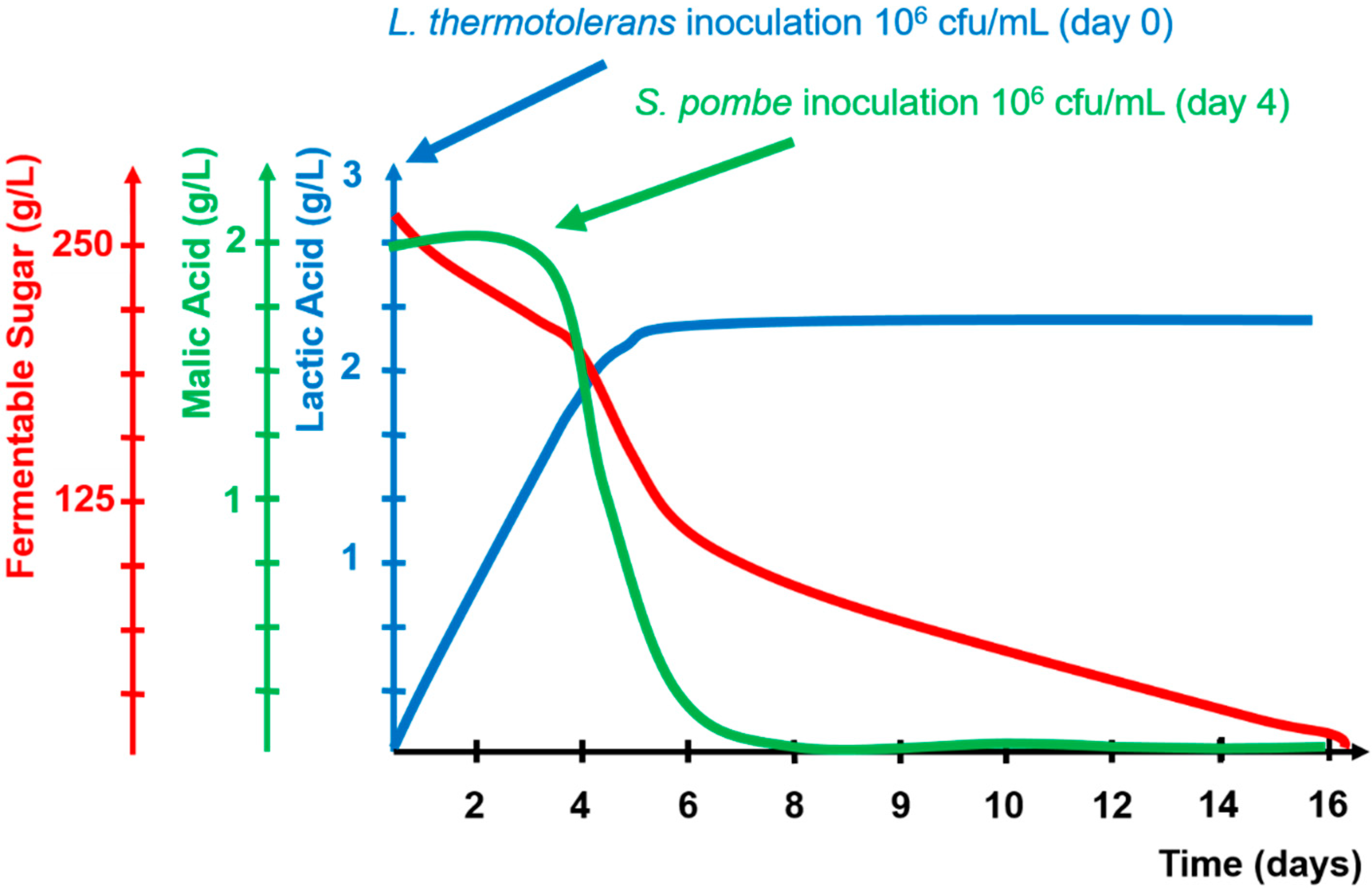
2.1. Fermentation Kinetics
2.2. Ethanol
2.3. Glycerol
2.4. L-Malic Acid
2.5. L-Lactic Acid
2.6. Citric Acid
2.7. Acetic Acid
2.8. pH and Total Acidity
2.9. Acetaldehyde
2.10. Amino Acids
2.11. Polysaccharides
2.12. Biogenic Amines
2.13. Ethyl Carbamate Precursors
2.14. Aroma Compounds
2.15. Color Intensity and Anthocyanins
2.16. Sensory Evaluations
3. Conclusions
Funding
Conflicts of Interest
Abbreviations
| LT | Lachancea thermotolerans |
| SP | Schizosaccharomyces pombe |
| LT+SP | L. thermotolerans and S. pombe combined alcoholic fermentation |
| SC | Saccharomyces cerevisiae |
| MLF | Malolactic fermentation |
| AF | Alcoholic fermentation |
| SC+MLF | Alcoholic fermentation by S. cerevisiae followed by malolactic fermentation by O. oeni. |
| LB | Lactic bacteria |
References
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2013, 14, 215–237. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Benito, S. The Influence of Non-Saccharomyces Species on Wine Fermentation Quality Parameters. Fermentation 2019, 5, 54. [Google Scholar] [CrossRef]
- Benito, S. The impact of Torulaspora delbrueckii yeast in winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 3081–3094. [Google Scholar] [CrossRef] [PubMed]
- Vilela, A. Lachancea thermotolerans, the Non-Saccharomyces Yeast that Reduces the Volatile Acidity of Wines. Fermentation 2018, 4, 56. [Google Scholar] [CrossRef]
- Porter, T.J.; Divol, B.; Setati, M.E. Lachancea yeast species: Origin, biochemical characteristics and oenological significance. Food Res. Int. 2019, 119, 378–389. [Google Scholar] [CrossRef]
- Ruiz, J.; Belda, I.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Rauhut, D.; Santos, A.; Benito, S. Analytical impact of Metschnikowia pulcherrima in the volatile profile of Verdejo white wines. Appl. Microbiol. Biotechnol. 2018, 102, 8501–8509. [Google Scholar] [CrossRef]
- Benito, S. The impacts of Schizosaccharomyces on winemaking. Appl. Microbiol. Biotechnol. 2019, 103, 4291–4312. [Google Scholar] [CrossRef]
- Dutraive, O.; Benito, S.; Fritsch, S.; Beisert, B.; Patz, C.-D.; Rauhut, D. Effect of Sequential Inoculation with Non-Saccharomyces and Saccharomyces Yeasts on Riesling Wine Chemical Composition. Fermentation 2019, 5, 79. [Google Scholar] [CrossRef]
- Du Plessis, H.; Du Toit, M.; Nieuwoudt, H.; Van der Rijst, M.; Hoff, J.; Jolly, N. Modulation of wine flavor using Hanseniaspora uvarum in combination with different Saccharomyces cerevisiae, lactic acid bacteria strains and malolactic fermentation strategies. Fermentation 2019, 5, 64. [Google Scholar] [CrossRef]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces Commercial Starter Cultures: Scientific Trends, Recent Patents and Innovation in the Wine Sector. Recent Pat. Food Nutr. Agric. 2019, 10, 1. [Google Scholar] [CrossRef]
- Berbegal, C.; Fragasso, M.; Russo, P.; Bimbo, F.; Grieco, F.; Spano, G.; Capozzi, V. Climate changes and food quality: The potential of microbial activities as mitigating strategies in the wine sector. Fermentation 2019, 5, 85. [Google Scholar] [CrossRef]
- Kapsopoulou, K.; Kapaklis, A.; Spyropoulos, H. Growth and fermentation characteristics of a strain of the wine yeast Kluyveromyces thermotolerans isolated in Greece. World J. Microbiol. Biotechnol. 2005, 21, 1599–1602. [Google Scholar] [CrossRef]
- Kapsopoulou, K.; Mourtzini, A.; Anthoulas, M.; Nerantzis, E. Biological acidification during grape must fermentation using mixed cultures of Kluyveromyces thermotolerans and Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2006, 23, 735–739. [Google Scholar] [CrossRef]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Benito, Á.; Calderón, F.; Palomero, F.; Benito, S. Quality and composition of airén wines fermented by sequential inoculation of Lachancea thermotolerans and Saccharomyces cerevisiae. Food Technol. Biotechnol. 2016, 54, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Balikci, E.K.; Tanguler, H.; Jolly, N.P.; Erten, H. Influence of Lachancea thermotolerans on cv. Emir wine fermentation. Yeast 2016, 33, 313–321. [Google Scholar] [CrossRef]
- Vilela, A. Use of nonconventional yeasts for modulating wine acidity. Fermentation 2019, 5, 27. [Google Scholar] [CrossRef]
- Nally, M.C.; Ponsone, M.L.; Pesce, V.M.; Toro, M.E.; Vazquez, F.; Chulze, S. Evaluation of behaviour of Lachancea thermotolerans biocontrol agents on grape fermentations. Lett. Appl. Microbiol. 2018, 67, 89–96. [Google Scholar] [CrossRef]
- Benito, S. The impacts of Lachancea thermotolerans yeast strains on winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 6775–6790. [Google Scholar] [CrossRef]
- Hranilovic, A.; Li, S.; Boss, P.K.; Bindon, K.; Ristic, R.; Grbin, P.R.; Van der Westhuizen, T.; Jiranek, V. Chemical and sensory profiling of Shiraz wines co-fermented with commercial non-Saccharomyces inocula. Aust. J. Grape Wine Res. 2017, 24, 166–180. [Google Scholar] [CrossRef]
- Benito, S.; Palomero, F.; Calderón, F.; Palmero, D.; Suárez-Lepe, J.A. Selection of appropriate Schizosaccharomyces strains for winemaking. Food Microbiol. 2014, 42, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Benito, A.; Jeffares, D.; Palomero, F.; Calderón, F.; Bai, F.-Y.; Bähler, J.; Benito, S. Selected Schizosaccharomyces pombe strains have characteristics that are beneficial for winemaking. PLoS ONE 2016, 11, e0151102. [Google Scholar] [CrossRef]
- Domizio, P.; Liu, Y.; Bisson, L.F.; Barile, D. Cell wall polysaccharides released during the alcoholic fermentation by Schizosaccharomyces pombe and S. japonicus: Quantification and characterization. Food Microbiol. 2016, 61, 136–149. [Google Scholar] [CrossRef]
- Domizio, P.; Lencioni, L.; Calamai, L.; Portaro, L.; Bisson, L.F. Evaluation of the yeast Schizosaccharomyces japonicus for use in wine production. Am. J. Enol. Vitic. 2018, 69, 266–277. [Google Scholar] [CrossRef]
- Liu, S.; Laaksonen, O.; Yang, W.; Zhang, B.; Yang, B. Pyranoanthocyanins in bilberry (Vaccinium myrtillus L.) wines fermented with Schizosaccharomyces pombe and their evolution during aging. Food Chem. 2020, 305, 125438. [Google Scholar] [CrossRef] [PubMed]
- Miljić, U.; Puškaš, V.; Vučurović, V.; Muzalevski, A. Fermentation Characteristics and Aromatic Profile of Plum Wines Produced with Indigenous Microbiota and Pure Cultures of Selected Yeast. J. Food Sci. 2017, 47, 1139–1450. [Google Scholar] [CrossRef] [PubMed]
- Minnaar, P.P.; Jolly, N.P.; Paulsen, V.; Du Plessis, H.W.; Van Der Rijst, M. Schizosaccharomyces pombe and Saccharomyces cerevisiae yeasts in sequential fermentations: Effect on phenolic acids of fermented Kei-apple (Dovyalis caffra L.) juice. Int. J. Food Microbiol. 2017, 257, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Roca-Domènech, G.; Cordero-Otero, R.; Rozès, N.; Cléroux, M.; Pernet, A.; Mira de Orduña, R. Metabolism of Schizosaccharomyces pombe under reduced osmotic stress conditions afforded by fed-batch alcoholic fermentation of white grape must. Food Res. Int. 2018, 113, 401–406. [Google Scholar] [CrossRef]
- Scomparin, A.; Bureik, M. A convenient new method for reproducible fed-batch fermentation of fission yeast Schizosaccharomyces pombe. Biotechnol. Lett. 2020, 1–7. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Palomero, F.; Benito, S. Combine use of selected Schizosaccharomyces pombe and Lachancea thermotolerans yeast strains as an alternative to the traditional malolactic fermentation in red wine production. Molecules 2015, 20, 9510–9523. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Benito, S. Schizosaccharomyces pombe and Lachancea thermotolerans: Joint Use as an Alternative to the Traditional Fermentations by Saccharomyces cerevisiae and Oenococcus oeni in Oenology. In Alcoholic Beverages; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Benito, A.; Calderón, F.; Benito, S. Combined use of S. pombe and L. thermotolerans in winemaking. Beneficial effects determined through the study of wines’ analytical characteristics. Molecules 2016, 21, 1744. [Google Scholar] [CrossRef] [PubMed]
- Benito, Á.; Calderón, F.; Benito, S. The combined use of Schizosaccharomyces pombe and Lachancea thermotolerans—Effect on the anthocyanin wine composition. Molecules 2017, 22, 739. [Google Scholar] [CrossRef] [PubMed]
- Escott, C.; Morata, A.; Ricardo-Da-Silva, J.M.; Callejo, M.J.; Del Carmen González, M.; Suarez-Lepe, J.A. Effect of Lachancea thermotolerans on the formation of polymeric pigments during sequential fermentation with schizosaccharosmyces pombe and Saccharomyces cerevisiae. Molecules 2018, 23, 2353. [Google Scholar] [CrossRef] [PubMed]
- Escott, C.; Del Fresno, J.M.; Loira, I.; Morata, A.; Tesfaye, W.; del Carmen González, C.; Suárez-Lepe, J.A. Formation of polymeric pigments in red wines through sequential fermentation of flavanol-enriched musts with non-Saccharomyces yeasts. Food Chem. 2018, 239, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Benito, Á.; Calderón, F.; Benito, S. Mixed alcoholic fermentation of Schizosaccharomyces pombe and Lachancea thermotolerans and its influence on mannose-containing polysaccharides wine Composition. AMB Express 2019, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Del Fresno, J.M.; Morata, A.; Loira, I.; Bañuelos, M.A.; Escott, C.; Benito, S.; González Chamorro, C.; Suárez-Lepe, J.A. Use of non-Saccharomyces in single-culture, mixed and sequential fermentation to improve red wine quality. Eur. Food Res. Technol. 2017, 243, 2175–2185. [Google Scholar] [CrossRef]
- Chen, K.; Escott, C.; Loira, I.; del Fresno, J.M.; Morata, A.; Tesfaye, W.; Calderon, F.; Suárez-Lepe, J.A.; Han, S.; Benito, S. Use of non-Saccharomyces yeasts and oenological tannin in red winemaking: Influence on colour, aroma and sensorial properties of young wines. Food Microbiol. 2018, 69, 51–63. [Google Scholar] [CrossRef]
- Wang, Y.; Sheng, W.; Li, M.; Mi, L.; Jiang, Y. Effect of Sequential Fermentation with Lachancea thermotolerans and Schizosaccharomyces pombe on the Quality of Merlot Dry Red Wine. Food Sci. 2019, 40, 102–111. [Google Scholar]
- Hu, C.K.; Bai, F.W.; An, L.J. Enhancing ethanol tolerance of a self-flocculating fusant of Schizosaccharomyces pombe and Saccharomyces cerevisiae by Mg2+ via reduction in plasma membrane permeability. Biotechnol. Lett. 2003, 25, 1191–1194. [Google Scholar] [CrossRef]
- du Plessis, H.W.; du Toit, M.; Hoff, J.W.; Hart, R.S.; Ndimba, B.K.; Jolly, N.P. Characterisation of non-Saccharomyces yeasts using different methodologies and evaluation of their compatibility with malolactic fermentation. S. Afr. J. Enol. Vitic. 2017, 38, 46–63. [Google Scholar] [CrossRef]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Implications of new research and technologies for malolactic fermentation in wine. Appl. Microbiol. Biotechnol. 2014, 98, 8111–8132. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Hranilovic, A.; Bely, M.; Masneuf-Pomarede, I.; Jiranek, V.; Albertin, W. The evolution of Lachancea thermotolerans is driven by geographical determination, anthropisation and flux between different ecosystems. PLoS ONE 2017, 12, e0184652. [Google Scholar] [CrossRef] [PubMed]
- Hranilovic, A.; Gambetta, J.M.; Schmidtke, L.; Boss, P.K.; Grbin, P.R.; Masneuf-Pomarede, I.; Bely, M.; Albertin, W.; Jiranek, V. Oenological traits of Lachancea thermotolerans show signs of domestication and allopatric differentiation. Sci. Rep. 2018, 8, 14812. [Google Scholar] [CrossRef] [PubMed]
- du Plessis, H.; du Toit, M.; Nieuwoudt, H.; van der Rijst, M.; Kidd, M.; Jolly, N. Effect of Saccharomyces, Non-Saccharomyces Yeasts and Malolactic Fermentation Strategies on Fermentation Kinetics and Flavor of Shiraz Wines. Fermentation 2017, 3, 64. [Google Scholar] [CrossRef]
- Mylona, A.E.; Del Fresno, J.M.; Palomero, F.; Loira, I.; Bañuelos, M.A.; Morata, A.; Calderón, F.; Benito, S.; Suárez-Lepe, J.A. Use of Schizosaccharomyces strains for wine fermentation-Effect on the wine composition and food safety. Int. J. Food Microbiol. 2016, 232, 63–72. [Google Scholar] [CrossRef]
- Ruiz, J.; Kiene, F.; Belda, I.; Fracassetti, D.; Marquina, D.; Navascués, E.; Calderón, F.; Benito, A.; Rauhut, D.; Santos, A.; et al. Effects on varietal aromas during wine making: A review of the impact of varietal aromas on the flavor of wine. Appl. Microbiol. Biotechnol. 2019, 103, 7425–7450. [Google Scholar] [CrossRef]
- Song, Z.; Du, H.; Zhang, M.; Nie, Y.; Xu, Y. Schizosaccharomyces pombe can reduce acetic acid produced by Baijiu spontaneous fermentation microbiota. Microorganisms 2019, 7, 606. [Google Scholar] [CrossRef]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbühler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Benito, S. The Management of Compounds that Influence Human Health in Modern Winemaking from an HACCP Point of View. Fermentation 2019, 5, 33. [Google Scholar] [CrossRef]
- Domizio, P.; Liu, Y.; Bisson, L.F.; Barile, D. Use of non-Saccharomyces wine yeasts as novel sources of mannoproteins in wine. Food Microbiol. 2014, 43, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Navascués, E.; Marquina, D.; Santos, A.; Calderón, F.; Benito, S. Outlining the influence of non-conventional yeasts in wine ageing over lees. Yeast 2016, 33, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Escott, C.; Loira, I.; Del Fresno, J.M.; Morata, A.; Tesfaye, W.; Calderon, F.; Benito, S.; Suárez-Lepe, J.A. The effects of pre-fermentative addition of oenological tannins on wine components and sensorial qualities of red wine. Molecules 2016, 21, 1445. [Google Scholar] [CrossRef] [PubMed]
- Escribano, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine aromatic compound production and fermentative behaviour within different non-Saccharomyces species and clones. J. Appl. Microbiol. 2018, 124, 1521–1531. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine aroma evolution throughout alcoholic fermentation sequentially inoculated with non-Saccharomyces/Saccharomyces yeasts. Food Res. Int. 2018, 112, 17–24. [Google Scholar] [CrossRef]
- Liu, S.; Laaksonen, O.; Yang, B. Volatile composition of bilberry wines fermented with non-Saccharomyces and Saccharomyces yeasts in pure, sequential and simultaneous inoculations. Food Microbiol. 2019, 80, 25–39. [Google Scholar] [CrossRef]
- Burns, T.R.; Osborne, J.P. Loss of pinot noir wine color and polymeric pigment after malolactic fermentation and potential causes. Am. J. Enol. Vitic. 2014, 66, 130–137. [Google Scholar] [CrossRef]
- Strickland, M.T.; Schopp, L.M.; Edwards, C.G.; Osborne, J.P. Impact of Pediococcus spp. on pinot noir wine quality and growth of Brettanomyces. Am. J. Enol. Vitic. 2016, 67, 188–198. [Google Scholar] [CrossRef]
- Wang, S.; Li, S.; Zhao, H.; Gu, P.; Chen, Y.; Zhang, B.; Zhu, B. Acetaldehyde released by Lactobacillus plantarum enhances accumulation of pyranoanthocyanins in wine during malolactic fermentation. Food Res. Int. 2018, 108, 254–263. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lacchancea thermotolerans | Schizosaccharomyces pombe | ||
|---|---|---|---|
| Advantages | Disadvantages | Advantages | Disadvantages |
| Acidity ↑ | Fermentative power ↓ | Malic acid ↓ | Acetic acid ↑ |
| Lactic acid ↑ | SO2 resistance ↓ | Polysaccharides ↑ | |
| Color intensity ↑ | Color intensity ↑ | ||
| Acetic acid ↓ | Color stability ↑ | ||
| Fruity esters ↑ | Biogenic amines ↓ | ||
| Ethyl carbamate ↓ | |||
| Fermentation Duration | Ethanol | Glycerol | Malic Acid | Lactic Acid | Citric Acid | Acetic Acid | pH | Total Acidity | Acetaldehyde | Polysaccharides | Biogenic Amines | Urea | Higher Alcohols | Esters | Terpenes | Color Intensity | Anthocyanins | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Benito et al. 2015 [30] | SC | = | ↑ | = | = | = | = | = | = | n.d.a | n.d.a | n.d.a | = | = | n.d.a | n.d.a | n.d.a | = | n.d.a |
| SC+MLF | ↑↑↑ | ↑ | = | ↓↓ | ↑ | ↓ | ↑ | ↑ | n.d.a | n.d.a | n.d.a | ↑ | n.d.a | n.d.a | n.d.a | n.d.a | ↓↓ | n.d.a | |
| LT+SC | ↑ | ↓ | ↑ | = | ↑↑ | = | ↓ | ↓ | n.d.a | n.d.a | n.d.a | = | = | n.d.a | n.d.a | n.d.a | ↑ | n.d.a | |
| LT+SC+MLF | ↑↑↑ | ↓ | ↑ | ↓↓ | ↑↑↑ | ↓ | ↑ | ↑ | n.d.a | n.d.a | n.d.a | ↑ | n.d.a | n.d.a | n.d.a | n.d.a | ↓ | n.d.a | |
| LT+SP | ↑↑ | ↓ | ↑↑ | ↓↓ | ↑↑ | = | = | ↓ | n.d.a | n.d.a | n.d.a | = | ↓↓ | n.d.a | n.d.a | n.d.a | ↑↑ | n.d.a | |
| SP | ↑ | ↓ | ↑↑ | ↓↓ | = | = | = | ↑↑ | n.d.a | n.d.a | n.d.a | = | ↓↓ | n.d.a | n.d.a | n.d.a | ↑↑↑ | n.d.a | |
| Benito et al. 2016 [32] | SC | = | ↑ | = | = | = | = | = | = | n.d.a | n.d.a | n.d.a | = | = | ↑ | = | n.d.a | = | n.d.a |
| SC+MLF | ↑↑↑ | ↑ | = | ↓↓ | ↑ | ↓ | ↑ | ↑ | n.d.a | n.d.a | = | ↑ | ↑ | ↑ | ↑ | n.d.a | ↓↓ | n.d.a | |
| LT+SC | ↑ | ↓ | ↑ | = | ↑↑ | = | = | ↓ | n.d.a | n.d.a | n.d.a | = | = | ↓ | ↑↑ | n.d.a | ↑ | n.d.a | |
| LT+SC+MLF | ↑↑↑ | ↓ | ↑ | ↓↓ | ↑↑↑ | ↓ | ↑ | ↑ | n.d.a | n.d.a | ↓ | ↑ | ↑ | ↑ | ↑ | n.d.a | ↓ | n.d.a | |
| LT+SP | ↑↑ | ↓ | ↑↑ | ↓↓ | ↑↑ | = | = | ↓ | n.d.a | n.d.a | ↑ | = | ↓↓ | ↓ | ↑↑ | n.d.a | ↑↑ | n.d.a | |
| SP | ↑ | ↓ | ↑↑↑ | ↓↓ | = | = | = | ↑↑ | n.d.a | n.d.a | ↑↑ | = | ↓↓ | ↓↓ | ↓ | n.d.a | ↑↑↑ | n.d.a | |
| Benito et al. 2017 [33] | SC | = | ↑ | = | = | = | = | = | = | n.d.a | = | n.d.a | n.d.a | = | n.d.a | n.d.a | n.d.a | = | = |
| SC+MLF | ↑↑↑ | ↑ | = | ↓↓ | ↑ | ↓ | ↑ | ↑ | n.d.a | ↓↓ | n.d.a | n.d.a | ↑ | n.d.a | n.d.a | n.d.a | ↓↓ | ↓ | |
| LT+SC | ↑ | ↓ | ↑ | = | ↑↑ | = | = | ↓ | n.d.a | ↓ | n.d.a | n.d.a | = | n.d.a | n.d.a | n.d.a | ↑ | ↑ | |
| LT+SC+MLF | ↑↑↑ | ↓ | ↑ | ↓↓ | ↑↑↑ | ↓ | ↑ | ↑ | n.d.a | ↓↓ | n.d.a | n.d.a | ↑ | n.d.a | n.d.a | n.d.a | ↓ | ↓ | |
| LT+SP | ↑↑ | ↓ | ↑↑ | ↓↓ | ↑↑ | = | = | ↓ | n.d.a | ↑ | n.d.a | n.d.a | ↓↓ | n.d.a | n.d.a | n.d.a | ↑↑ | ↑ | |
| SP | ↑ | ↓ | ↑↑ | ↓↓ | = | = | = | ↑↑ | n.d.a | ↑↑ | n.d.a | n.d.a | ↓↓ | n.d.a | n.d.a | n.d.a | ↑↑↑ | ↑↑ | |
| Fresno et al. 2017 [37] | SC | = | ↑ | = | = | = | n.d.a | ↓ | = | = | n.d.a | n.d.a | n.d.a | n.d.a | ↑ | = | n.d.a | = | ↑ |
| SC+MLF | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | |
| LT+SC | = | ↓ | ↑ | = | ↑↑ | n.d.a | ↓ | = | ↓ | n.d.a | n.d.a | n.d.a | n.d.a | ↑ | = | n.d.a | = | = | |
| LT+SC+MLF | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | |
| LT+SP | = | ↓↓ | ↓ | ↓↓ | ↑ | n.d.a | ↑ | ↑ | ↓↓ | n.d.a | n.d.a | n.d.a | n.d.a | ↓ | ↑↑ | n.d.a | ↑ | = | |
| SP | = | ↑ | ↑↑ | ↓↓ | = | n.d.a | ↑↑ | ↑ | ↓↓ | n.d.a | n.d.a | n.d.a | n.d.a | ↓↓ | ↓ | n.d.a | ↑ | ↑↑ | |
| Escott et al. 2018a [34] | SC | = | ↑ | n.d.a | = | = | n.d.a | = | = | = | n.d.a | n.d.a | n.d.a | n.d.a | = | = | n.d.a | = | ↑ |
| SC+MLF | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | |
| LT+SC | = | ↑ | n.d.a | ↓ | ↑ | n.d.a | ↓↓ | = | ↓ | n.d.a | n.d.a | n.d.a | n.d.a | ↑↑ | ↑ | n.d.a | ↑ | = | |
| LT+SC+MLF | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | |
| LT+SP | = | ↓ | n.d.a | ↓↓ | ↑ | n.d.a | ↓↓ | ↑ | ↓↓ | n.d.a | n.d.a | n.d.a | n.d.a | ↑ | ↑↑ | n.d.a | ↑ | = | |
| SP | = | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | |
| Escott et al. 2018b [35] | SC | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a |
| SC+MLF | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | |
| LT+SC | = | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | = | = | n.d.a | = | = | |
| LT+SC+MLF | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | |
| LT+SP | = | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | = | = | n.d.a | ↑ | = | |
| SP | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | |
| Benito et al. 2019 [36] | SC | = | ↑ | = | = | = | = | = | = | n.d.a | = | = | n.d.a | = | n.d.a | n.d.a | n.d.a | = | n.d.a |
| SC+MLF | ↑↑↑ | ↑ | = | ↓↓ | ↑ | ↓ | ↑ | ↑ | n.d.a | ↓↓ | = | n.d.a | ↑ | n.d.a | n.d.a | n.d.a | ↓↓ | n.d.a | |
| LT+SC | ↑ | ↓ | = | = | ↑↑ | = | = | ↓ | n.d.a | ↓ | ↓ | n.d.a | = | n.d.a | n.d.a | n.d.a | ↑ | n.d.a | |
| LT+SC+MLF | ↑↑↑ | ↓ | = | ↓↓ | ↑↑↑ | ↓ | ↑ | ↑ | n.d.a | ↓↓ | ↓ | n.d.a | ↑ | n.d.a | n.d.a | n.d.a | ↓ | n.d.a | |
| LT+SP | ↑↑ | ↓ | ↑ | ↓↓ | ↑↑ | = | = | ↓ | n.d.a | ↑ | ↑ | n.d.a | ↓↓ | n.d.a | n.d.a | n.d.a | ↑↑ | n.d.a | |
| SP | ↑ | ↓ | ↑ | ↓↓ | = | = | = | ↑↑ | n.d.a | ↑↑ | ↑↑ | n.d.a | ↓↓ | n.d.a | n.d.a | n.d.a | ↑↑↑ | n.d.a | |
| Wang et al. 2019 [39] | SC | = | ↑ | n.d.a | = | ↑ | n.d.a | = | = | = | n.d.a | n.d.a | n.d.a | n.d.a | ↑ | ↑ | = | = | n.d.a |
| SC+MLF | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | |
| LT+SC | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | |
| LT+SC+MLF | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | n.d.a | |
| LT+SP | ↑↑ | ↓↓ | n.d.a | ↓ | ↑↑ | n.d.a | ↓ | ↓ | ↑ | n.d.a | n.d.a | n.d.a | n.d.a | ↓ | = | ↑↑ | ↑ | n.d.a | |
| SP | ↑↑ | ↓ | n.d.a | ↓ | = | n.d.a | ↓↓ | ↑ | ↓ | n.d.a | n.d.a | n.d.a | n.d.a | ↓↓ | = | ↑↑ | ↑↑ | n.d.a |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benito, S. Combined Use of Lachancea thermotolerans and Schizosaccharomyces pombe in Winemaking: A Review. Microorganisms 2020, 8, 655. https://doi.org/10.3390/microorganisms8050655
Benito S. Combined Use of Lachancea thermotolerans and Schizosaccharomyces pombe in Winemaking: A Review. Microorganisms. 2020; 8(5):655. https://doi.org/10.3390/microorganisms8050655
Chicago/Turabian StyleBenito, Santiago. 2020. "Combined Use of Lachancea thermotolerans and Schizosaccharomyces pombe in Winemaking: A Review" Microorganisms 8, no. 5: 655. https://doi.org/10.3390/microorganisms8050655
APA StyleBenito, S. (2020). Combined Use of Lachancea thermotolerans and Schizosaccharomyces pombe in Winemaking: A Review. Microorganisms, 8(5), 655. https://doi.org/10.3390/microorganisms8050655