Effects of the Fungal Endophyte Epichloë festucae var. lolii on Growth and Physiological Responses of Perennial Ryegrass cv. Fairway to Combined Drought and Pathogen Stresses


Abstract
1. Introduction
2. Materials and Methods
2.1. Plants, Fungi, and Potting Medium
2.2. Experiment Design and Management
2.3. Protocols for Measurement and Determination of the Various Parameters
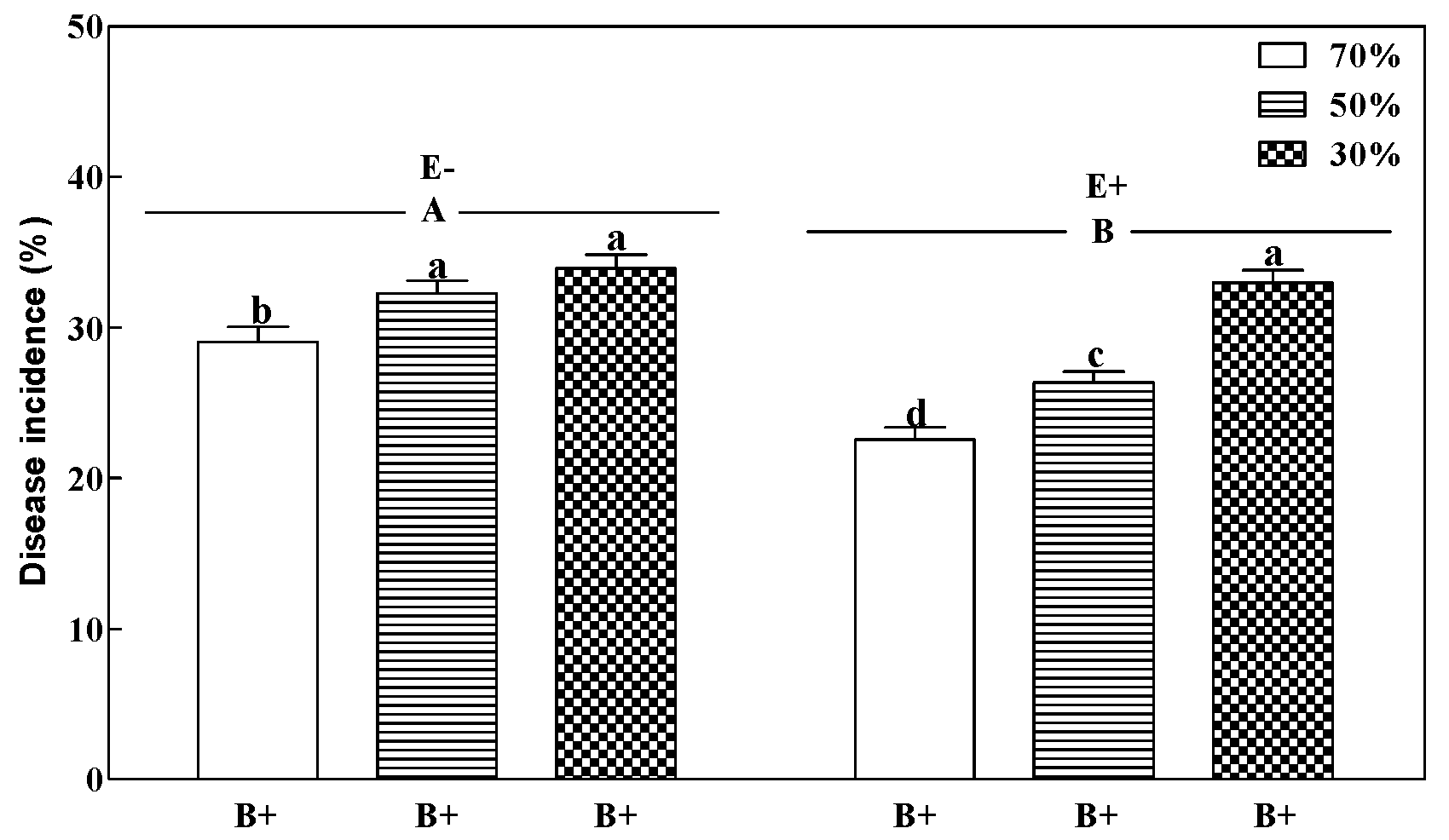
2.3.1. Disease Incidence
2.3.2. Chlorophyll Content
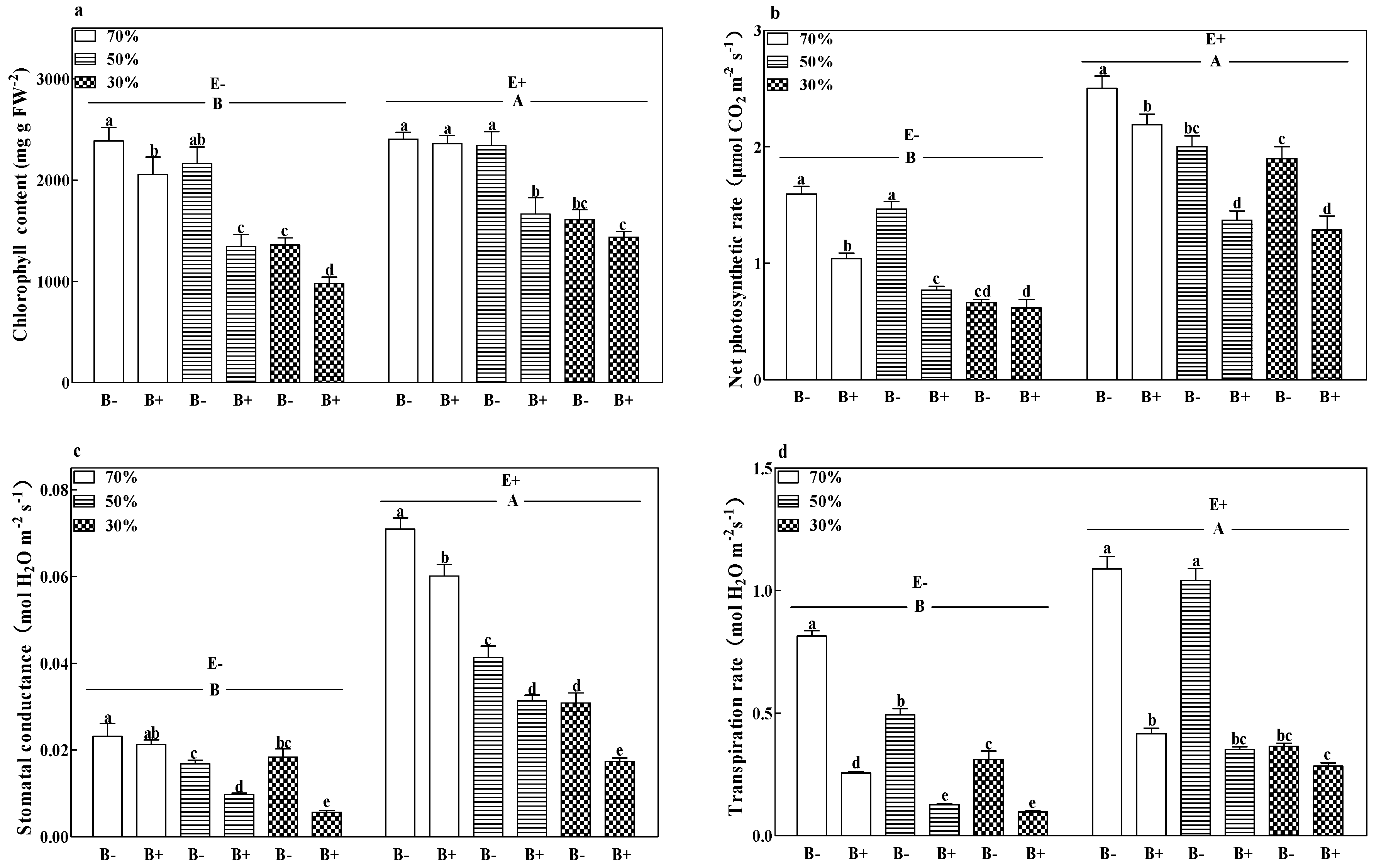
2.3.3. Photosynthetic Parameters
2.3.4. Catalase (CAT) and Peroxidase (POD) Enzyme Activities
2.3.5. Malondialdehyde (MDA)
2.3.6. Proline
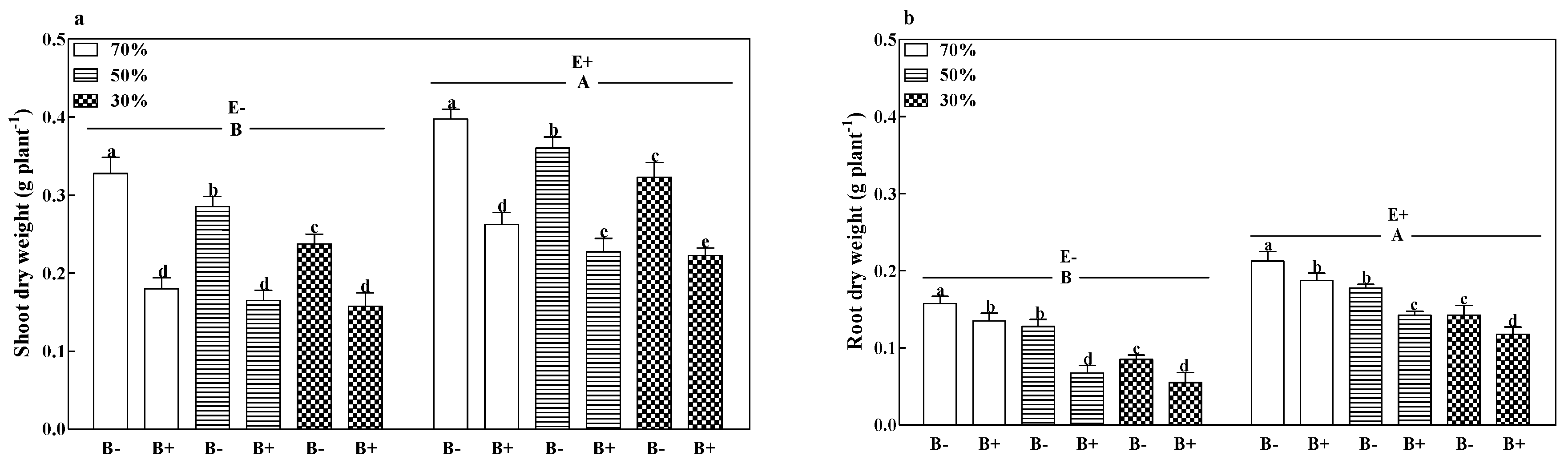
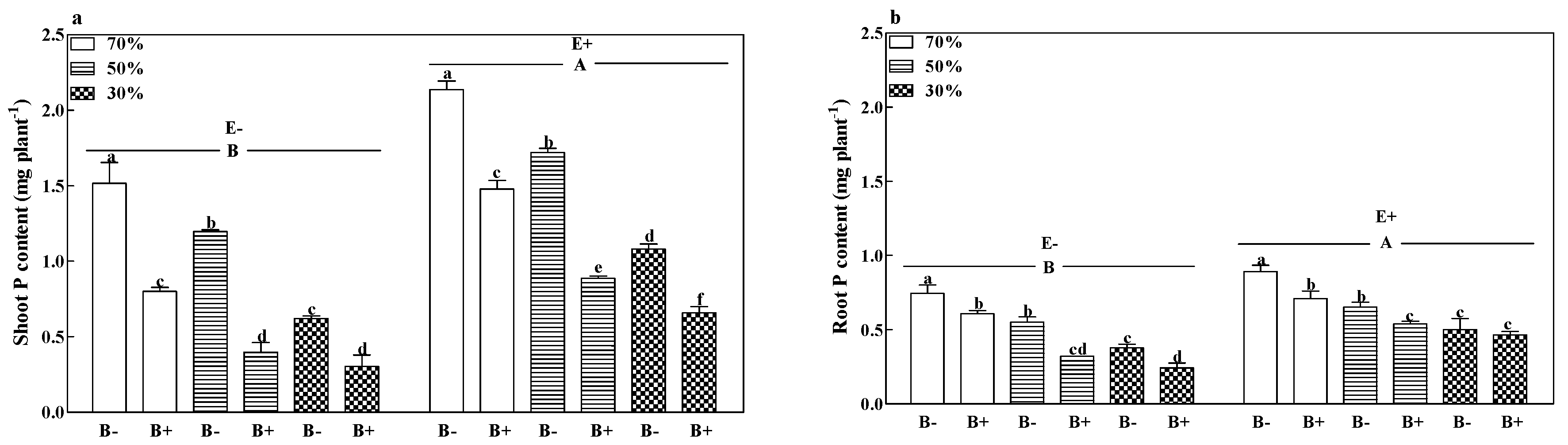
2.3.7. Plant Dry Weight and Phosphorus Content
2.4. Statistical Analysis
3. Results
3.1. Disease Incidence
3.2. Photosynthetic Parameters
3.3. Plant Growth
3.4. Phosphorous Contents
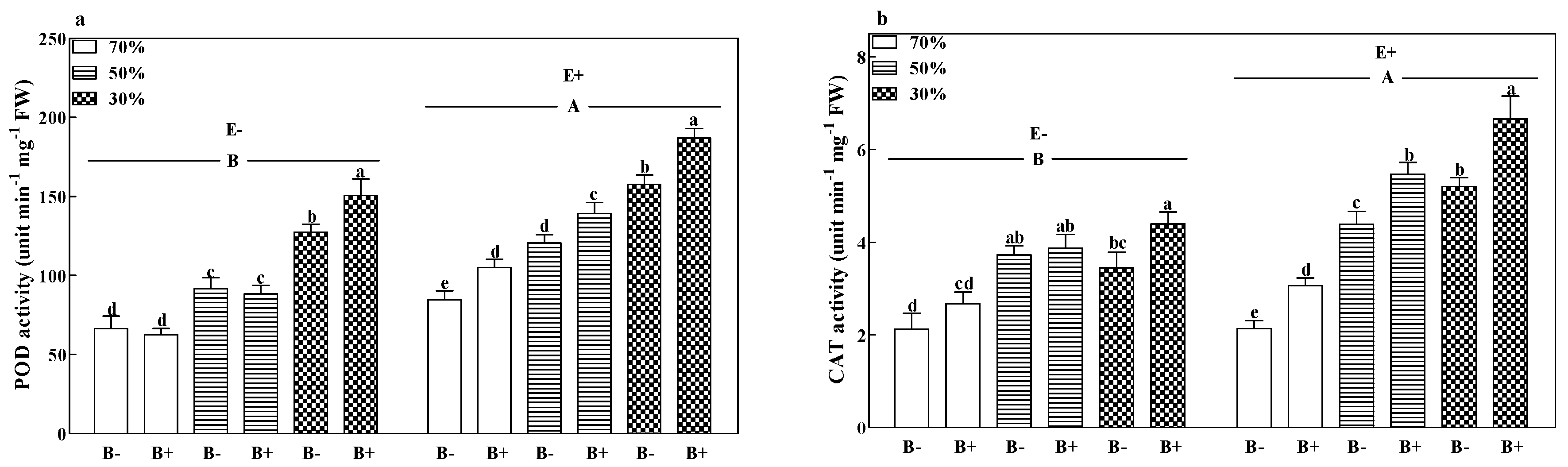
3.5. Enzyme Activity
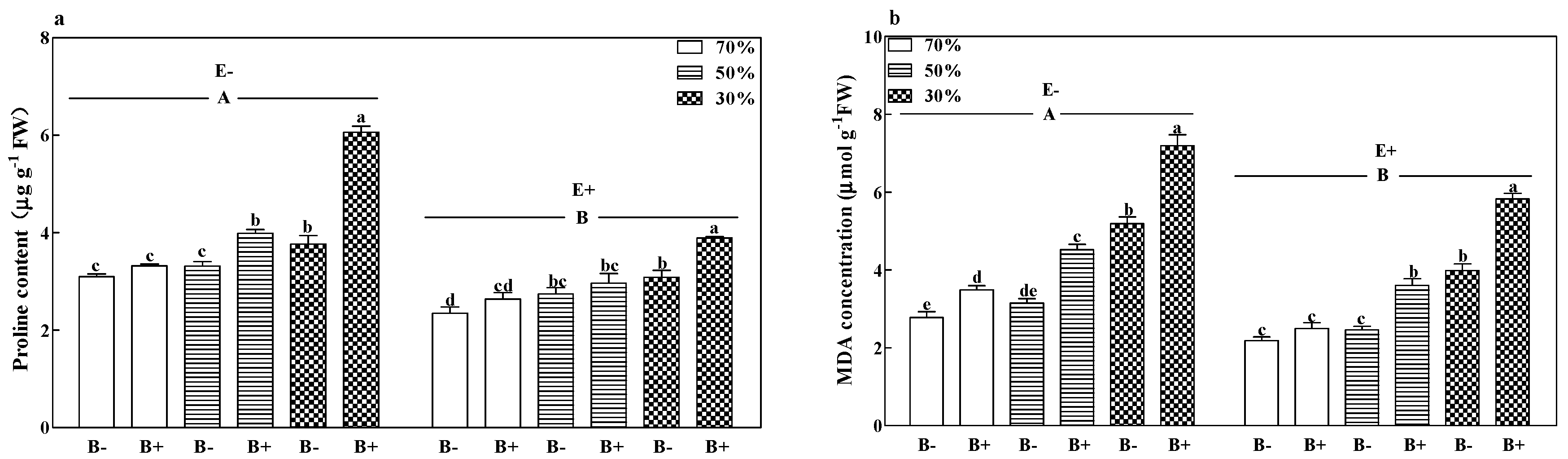
3.6. Proline Content and MDA Concentration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yordanov, I.; Velikova, V.; Tsonev, T. Plant Responses to Drought, Acclimation, and Stress Tolerance. Photosynthetica 2000, 38, 171–186. [Google Scholar] [CrossRef]
- Reed, K.F.M. Improving the adaptation of perennial ryegrass, tall fescue, phalaris, and cocksfoot for Australia. N. Z. J. Agric. Res. 1996, 39, 457–464. [Google Scholar] [CrossRef]
- Clarke, R.G.; Eagling, D.R. Effects of pathogens on perennial pasture grasses. N. Z. J. Agric. Res. 1994, 37, 319–327. [Google Scholar] [CrossRef][Green Version]
- Jung, G.A.; Van Wijk, A.J.P.; Hunt, W.F.; Watson, C.E. Ryegrasses. In Cool-Season Forage Grasses; Moser, L.E., Buxton, D.R., Casler, M.D., Eds.; ASA CSSA SSSA: Madison, WI, USA, 1996; pp. 605–641. [Google Scholar]
- Uddin, W.; Soika, M.D.; Moorman, F.E.; Sitther, V. A serious outbreak of blast disease (gray leaf spot) of perennial ryegrass in golf course fairways in Pennsylvania. Plant Dis. 1999, 83, 783. [Google Scholar] [CrossRef]
- Smiley, R.W.; Dernoeden, P.H.; Clarke, B.B. Compendium of Turfgrass Diseases. In The American Phytopathological Society; Smiley, R.W., Dernoeden, P.H., Clarke, B.B., Eds.; The American Phytopathological Society: St. Paul, MI, USA, 1992. [Google Scholar]
- Tian, P.; Nan, Z.B.; Li, C.J.; Spangenberg, G. Effect of the endophyte Neotyphodium lolii on susceptibility and host physiological response of perennial ryegrass to fungal pathogens. Eur. J. Plant Pathol. 2008, 122, 593–602. [Google Scholar] [CrossRef]
- Clay, K. Fungi and the food of the gods. Nature 2004, 427, 401–402. [Google Scholar] [CrossRef]
- Zhang, W.; Card, S.D.; Mace, W.J.; Christensen, M.J.; Mcgill, C.R.; Matthew, C. Defining the pathways of symbiotic Epichloë colonization in grass embryos with confocal microscopy. Mycologia 2017, 109, 153–161. [Google Scholar] [CrossRef]
- Craven, K.D. Population studies of native grass-endophyte symbioses provide clues for the roles of host jumps and hybridization in driving their evolution. Mol. Ecol. 2012, 21, 2562–2564. [Google Scholar] [CrossRef]
- Müller, C.B.; Krauss, J. Symbiosis between grasses and asexual fungal endophytes. Curr. Opin. Plant Biol. 2005, 8, 450–456. [Google Scholar] [CrossRef]
- Schardl, C.L.; Leuchtmann, A.; Spiering, M.J. Symbioses of grasses with seedborne fungal endophytes. Annu. Rev. Plant Biol. 2004, 55, 315–340. [Google Scholar] [CrossRef]
- Tian, Z.P. Effects of Epichloë festucae Fungal Endophyte Infection in Strong Creeping Red Fescue on Abiotic and Biotic Stresses. Ph.D. Thesis, Rutgers University, Graduate School, New Brunswick, NJ, Canada, 2017. [Google Scholar]
- Peng, Q.Q.; Li, C.J.; Song, M.L.; Nan, Z.B. Effects of seed hydropriming on growth of Festuca sinensis, infected with Neotyphodium endophyte. Fungal Ecol. 2013, 6, 83–91. [Google Scholar] [CrossRef]
- Raizada, M.N.; Johnstonmonje, D. Plant and Endophyte Relationships: Nutrient Management. In Comprehensive Biotechnology; Moo-Young, M., Ed.; Academic Publishers: Burlington, VT, USA, 2011; pp. 713–727. [Google Scholar]
- Liu, H.; Chen, W.; Wu, M.; Wu, R.H.; Zhou, Y.; Gao, Y.B.; Ren, A.Z. Arbuscular mycorrhizal fungus inoculation reduces the drought resistance advantage of endophyte-infected versus endophyte-free Leymus chinensis. Mycorrhiza 2017, 27, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, X.; Han, L.; Li, D.; Song, G. Epichloë endophyte infection improved drought and heat tolerance of tall fescue through altered antioxidant enzyme activity. Eur. J. Hortic. Sci. 2017, 82, 90–97. [Google Scholar] [CrossRef]
- Wiewióra, B.; Żurek, G.; Żurek, M. Endophyte-mediated disease resistance in wild populations of perennial ryegrass (Lolium perenne). Fungal Ecol. 2015, 15, 1–8. [Google Scholar] [CrossRef]
- Li, F.; Guo, Y.E.; Christensen, M.J.; Gao, P.; Li, Y.Z.; Duan, T.Y. An arbuscular mycorrhizal fungus and Epichloë festucae var. lolii reduce Bipolaris sorokiniana disease incidence and improve perennial ryegrass growth. Mycorrhiza 2018, 28, 159–169. [Google Scholar] [CrossRef]
- Ma, M.Z.; Christensen, M.J.; Nan, Z.B. Effects of the endophyte Epichloë festucae var. lolii of perennial ryegrass (Lolium perenne) on indicators of oxidative stress from pathogenic fungi during seed germination and seedling growth. Eur. J. Plant Pathol. 2015, 141, 571–583. [Google Scholar] [CrossRef]
- White, J.F.; Cole, G.T. Endophyte–host association in forage grasses: III. In ‘vitro inhibition of fungi by Acreamnium coenophialum’. Mycologia 1985, 77, 487–489. [Google Scholar] [CrossRef]
- Elmi, A.A.; West, C.P. Endophyte infection effects on stomatal conductance, osmotic adjustment and drought recovery of tall fescue. New Phytol. 2010, 131, 61–67. [Google Scholar] [CrossRef]
- Clay, K.; Schardl, C. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. Am. Nat. 2002, 160, S99–S127. [Google Scholar] [CrossRef]
- Faeth, S.H.; Bush, L.P.; Sullivan, T.J. Peramine Alkaloid variation in Neotyphodium-infected Arizona Fescue: Effects of endophyte and host genotype and environment. J. Chem. Ecol. 2002, 28, 1511–1526. [Google Scholar] [CrossRef]
- Li, F.; Deng, J.; Nzabanita, C.; Duan, T.Y. Growth and physiological responses of perennial ryegrass to an AMF and an Epichloë endophyte under different soil water contents. Symbiosis 2019, 79, 151–161. [Google Scholar] [CrossRef]
- Deng, J.; Fang, L.; Duan, T.Y. Claroideoglomus etunicatum reduces leaf spot incidence and improves drought stress resistance in perennial ryegrass. Australas. Plant Pathol. 2020, 29, 147–157. [Google Scholar] [CrossRef]
- Xia, C.; Zhang, X.X.; Christensen, M.J.; Nan, Z.B.; Li, C.J. Epichloë endophyte affects the ability of powdery mildew (Blumeria graminis) to colonise drunken horse grass (Achnatherum inebrians). Fungal Ecol. 2015, 16, 26–33. [Google Scholar] [CrossRef]
- Nan, Z.B. Incidence and distribution of endophytic fungi in seeds of some native and introduced grasses in China. Acta Pratacult. Sinica 1996, 5, 1–8. [Google Scholar]
- Yu, O.Y.; Harper, M.; Hoepfl, M.; Domermuth, D. Characterization of biochar and its effects on the water holding capacity of loamy sand soil: Comparison of hemlock biochar and switchblade grass biochar characteristics. Environ. Prog. Sustain. 2017, 36, 1474–1479. [Google Scholar] [CrossRef]
- Duan, T.; Facelli, E.; Smith, S.E.; Smith, F.A.; Nan, Z.B. Differential effects of soil disturbance and plant residue retention on function of arbuscular mycorrhizal (AM) symbiosis are not reflected in colonization of roots or hyphal development in soil. Soil Biol. Biochem. 2011, 43, 571–578. [Google Scholar] [CrossRef]
- Florea, S.; Schardl, C.L.; Hollin, W. Detection and isolation of Epichloë species, fungal endophytes of grasses. Curr. Protoc. Microbiol. 2015, 38, 19A.1.1–19A.1.24. [Google Scholar] [CrossRef]
- Starnes, W.J.; Hadley, H.H. Chlorophyll content of various strains of soybeans, Glycine max. Crop Sci. 1965, 5, 9–11. [Google Scholar] [CrossRef]
- Beers, R.F.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar]
- Chance, B.; Maehly, C. Assay of catalase and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Li, H.S.; Sun, Q.; Zhao, S.J.; Zhang, W.H. Principle and Techniques of Botanic, Chemical and Physiological Experiments, 1st ed.; Senior Education Press: Beijing, China, 2000. [Google Scholar]
- Hanson, W.C. The photometric determination of phosphorus in fertilizers using the phosphovanado-molybdate complex. J. Sci. Food Agric. 1950, 1, 172–173. [Google Scholar] [CrossRef]
- Xia, C.; Li, N.N.; Zhang, X.X.; Feng, Y.; Christensen, M.J.; Nan, Z.B. An Epichloë endophyte improves photosynthetic ability and dry matter production of its host Achnatherum inebrians infected by Blumeria graminis under various soil water conditions. Fungal Ecol. 2016, 22, 26–34. [Google Scholar] [CrossRef]
- Mitchell, P.J.; O’Grady, A.P.; Tissue, D.T.; Worledge, D.; Pinkard, E.A. Co-ordination of growth, gas exchange and hydraulics define the carbon safety margin in tree species with contrasting drought strategies. Tree Physiol. 2014, 33, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Ayres, P.G. Patterns of stomatal behaviour, transpiration, and CO2 exchange in pea following infection by powdery mildew (Erysiphe pisi). J. Exp. Bot. 1976, 27, 1196–1205. [Google Scholar] [CrossRef]
- Alam, S.M. Nutrient Uptake by Plants Under Stress Conditions. In Handbook of Plant and Crop Stress; Pessarakli, M., Ed.; Marcel Dekker: New York, NY, USA, 1999; pp. 285–314. [Google Scholar]
- Moriondo, M.; Orlandini, S.; Giuntoli, A.; Bindi, M. The effect of downy and powdery mildew on grapevine (Vitis vinifera) leaf gas exchange. J. Phytopathol. 2005, 153, 350–357. [Google Scholar] [CrossRef]
- Gao, P.; Duan, T.Y.; Christensen, M.J.; Nan, Z.B.; Liu, Q.T.; Meng, F.J.; Huang, J.F. The occurrence of rust disease, and biochemical and physiological responses on Apocynum venetum plants grown at four soil water contents, following inoculation with Melampsora apocyni. Eur. J. Plant Pathol. 2018, 150, 549–563. [Google Scholar] [CrossRef]
- Li, C.J.; Gao, J.H.; Nan, Z.B. Interactions of Neotyphodium gansuense, Achnatherum inebrians, and plant-pathogenic fungi. Mycol. Res. 2007, 111, 1220–1227. [Google Scholar] [CrossRef]
- Holzmann-Wirth, A.; Dapprich, P.; Eierdanz, S.; Heerz, D.; Paul, V.H. Anti-Fungal Substances Extracted from Neotyphodium Endophyte. In Proceedings of the 3rd International Conference on Harmful and Benefical Microorganisms in Grassland, Pasture and Turf, Soest, Germany, 4–6 October 1993; pp. 65–69. [Google Scholar]
- Nan, Z.B.; Li, C.J. Roles of the grass-Neotyphodium association in pastoral agriculture systems. Acta Ecol. Sin. 2004, 24, 605–616. [Google Scholar]
- Guo, Y.E.; Gao, P.; Li, F.; Duan, T.Y. Effects of AM fungi and grass endophytes on perennial ryegrass Bipolaris sorokiniana leaf spot disease under limited soil nutrients. Eur. J. Plant Pathol. 2019, 154, 659–671. [Google Scholar] [CrossRef]
- Saifuddin, M.; Normaniza, O. Physiological and root profile studies of four legume tree species. Life Sci. J. 2002, 9, 1509–1518. [Google Scholar]
- Parthasarathi, T.; Vanitha, K.; Lakshamanakumar, P.; Kalaiyarasi, D. Aerobic rice-mitigating water stress for the future climate change. Int. J. Agron. Plant Prod. 2012, 3, 241–254. [Google Scholar]
- Liu, N.; Lei, Y.; Gong, G.S.; Zhang, M.; Wang, X.; Zhou, Y.; Qi, X.B.; Chen, H.B.; Yang, J.Z.; Chang, X.L.; et al. Temporal and spatial dynamics of wheat powdery mildew in Sichuan Province, China. Crop Prot. 2015, 74, 150–157. [Google Scholar] [CrossRef]
- Liu, Z.H.; Xu, M.; Xiang, Z.P.; Li, X.; Chen, W.Q.; Luo, P.G. Registration of the novel wheat lines L658, L693, and L699, with resistance to Fusarium head blight, stripe rust and powdery mildew. J. Plant Regist. 2015, 9, 121–124. [Google Scholar] [CrossRef]
- Smith, S.E.; Jakobsen, I.; Grønlund, M.; Smith, A. Roles of arbuscular mycorrhizas in plant phosphorus nutrition: Interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol. 2011, 156, 1050–1057. [Google Scholar] [CrossRef]
- Zhang, X.X.; Xia, C.; Nan, Z.B. Effects of symbiotic Epichloë gansuensis endophyte on drunken horse grass (Achnatherum inebrians) growth and seed production. N. Z. J. Agric. Res. 2015, 58, 234–240. [Google Scholar] [CrossRef]
- Malinowski, D.P.; Belesky, D.P. Neotyphodium coenophialum-endophyte infection affects the ability of tall fescue to use sparingly available phosphorus. J. Plant Nutr. 1999, 22, 835–853. [Google Scholar] [CrossRef]
- Amalric, C.; Sallanon, H.; Monnet, F.; Hitmi, A.; Coudret, A. Gas exchange and chlorophyll fluorescence in symbiotic and non-symbiotic ryegrass under water stress. Photosynthetica 1999, 37, 107–112. [Google Scholar] [CrossRef]
- Pinto, L.S.R.C.; Azevedo, J.L.; Pereira, J.O.; Vieira, M.L.C.; Labate, C.A. Symptomless infection of banana and maize by endophytic fungi impairs photosynthetic efficiency. New Phytol. 2000, 147, 609–615. [Google Scholar] [CrossRef]
- Ju, Y.L.; Yue, X.F.; Zhao, X.F.; Zhao, H.; Fang, Y.L. Physiological, micro-morphological and metabolomic analysis of grapevine (Vitis vinifera) leaf of plants under water stress. Plant Physiol. Biochem. 2018, 130, 501–510. [Google Scholar] [CrossRef]
- Upadhyaya, H.; Khan, M.P. Hydrogen peroxide induces oxidative stress in detached leaves of Oryza sativa. Gen. Appl. Plant Physiol. 2007, 33, 83–95. [Google Scholar]
- Wang, D.Q.; Li, G.Q.; Wang, L. Daily dynamics of photosynthesis and water physiological characteristics of Apocynum venetum and A. cannabinum under drought Stress. Acta Bot. Boreali-Occident. Sin. 2012, 32, 1198–1205. [Google Scholar]
- Rodriguez, R.J.; Henson, J.; van Volkenburgh, E.; Hoy, M.; Wright, L.; Beckwith, F.; Yong-Ok, K.; Redman, R.S. Stress tolerance in plants via habitat-adapted symbiosis. Int. Soc. Microb. Ecol. 2008, 2, 404–416. [Google Scholar] [CrossRef]
- Srinivasan, K.; Jagadish, L.K.; Shenbhagaraman, R.; Muthumary, J. Antioxidant activity of endophytic fungus Phyllosticta sp. Isolated from Guazuma tomentosa. J. Phytol. 2010, 2, 37–41. [Google Scholar]
- Zhang, Y.P.; Nan, Z.B. Germination and seedling anti-oxidative enzymes of endophyte-infected populations of Elymus dahuricus under osmotic stress. Seed Sci. Technol. 2010, 38, 522–527. [Google Scholar] [CrossRef]
- Hamilton, C.E.; Gundel, P.E.; Helander, M.; Saikkonen, K. Endophytic mediation of reactive oxygen species and antioxidant activity in plants: A review. Fungal Divers. 2012, 54, 1–10. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Grass Endophyte (E) | Soil Water Content (W) | B. Sorokiniana (B) | Interactions | ||||
|---|---|---|---|---|---|---|---|---|
| ExW | ExB | WxB | ExWxB | |||||
| df | 1 | 1 | 2 | 3 | 5 | 5 | 11 | |
| Disease incidence | F | 155.033 | 152.409 | 27161.57 | 24.179 | 155.033 | 152.409 | 24.179 |
| P | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Chlorophyll content | F | 52.782 | 259.563 | 137.334 | 3.515 | 11.45 | 27.835 | 0.701 |
| P | <0.001 | <0.001 | <0.001 | 0.040 | 0.002 | <0.001 | 0.503 | |
| Net photosynthetic rate | F | 1018.041 | 244.494 | 319.359 | 28.523 | 2.636 | 13.831 | 21.097 |
| P | <0.001 | <0.001 | <0.001 | <0.001 | 0.118 | <0.001 | <0.001 | |
| Stomatal conductance | F | 1705.375 | 593.57 | 216.573 | 208.985 | 11.202 | 9.737 | 3.626 |
| P | <0.001 | <0.001 | <0.001 | <0.001 | 0.003 | 0.001 | 0.042 | |
| Transpiration rate | F | 740.504 | 624.127 | 2351.147 | 76.481 | 32.306 | 262.622 | 55.414 |
| P | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Shoot DW | F | 296.05 | 57.346 | 777.243 | 0.811 | 0.603 | 12.779 | 1.779 |
| P | <0.001 | <0.001 | <0.001 | 0.453 | 0.443 | <0.001 | 0.183 | |
| Root DW | F | 483.049 | 254.449 | 153.646 | 1.712 | 3.917 | 8.824 | 2.867 |
| P | 0.003 | <0.001 | <0.001 | 0.195 | 0.055 | 0.001 | 0.070 | |
| Total DW | F | 908.904 | 291.632 | 1208.389 | 0.111 | 0.196 | 17.12 | 1.056 |
| P | <0.001 | <0.001 | <0.001 | 0.895 | 0.661 | <0.001 | 0.359 | |
| Shoot P content | F | 734.088 | 604.166 | 1058.215 | 13.315 | 0.523 | 47.757 | 1.511 |
| P | <0.001 | <0.001 | <0.001 | <0.001 | 0.477 | <0.001 | 0.241 | |
| Root P content | F | 135.529 | 232.609 | 111.113 | 1.168 | 4.661 | 4.215 | 3.677 |
| P | <0.001 | <0.001 | <0.001 | 0.328 | 0.041 | 0.027 | 0.040 | |
| Total P content | F | 720.775 | 703.723 | 922.228 | 5.065 | 0.38 | 40.04 | 0.355 |
| P | <0.001 | <0.001 | <0.001 | 0.015 | 0.544 | <0.001 | 0.705 | |
| POD | F | 260.567 | 427.909 | 43.127 | 1.747 | 16.454 | 8.105 | 1.755 |
| P | <0.001 | <0.001 | <0.001 | 0.196 | <0.001 | 0.002 | 0.194 | |
| CAT | F | 138.935 | 241.141 | 80.877 | 30.678 | 10.288 | 3.639 | 0.817 |
| P | <0.001 | <0.001 | <0.001 | <0.001 | 0.004 | 0.042 | 0.454 | |
| MDA | F | 330.853 | 1023.702 | 542.854 | 9.4 | 6.157 | 59.414 | 0.402 |
| P | <0.001 | <0.001 | <0.001 | 0.001 | 0.020 | <0.001 | 0.673 | |
| Proline | F | 589.56 | 392.786 | 346.325 | 30.919 | 58.571 | 99.729 | 32.424 |
| P | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Duan, T.; Li, Y. Effects of the Fungal Endophyte Epichloë festucae var. lolii on Growth and Physiological Responses of Perennial Ryegrass cv. Fairway to Combined Drought and Pathogen Stresses. Microorganisms 2020, 8, 1917. https://doi.org/10.3390/microorganisms8121917
Li F, Duan T, Li Y. Effects of the Fungal Endophyte Epichloë festucae var. lolii on Growth and Physiological Responses of Perennial Ryegrass cv. Fairway to Combined Drought and Pathogen Stresses. Microorganisms. 2020; 8(12):1917. https://doi.org/10.3390/microorganisms8121917
Chicago/Turabian StyleLi, Fang, Tingyu Duan, and Yanzhong Li. 2020. "Effects of the Fungal Endophyte Epichloë festucae var. lolii on Growth and Physiological Responses of Perennial Ryegrass cv. Fairway to Combined Drought and Pathogen Stresses" Microorganisms 8, no. 12: 1917. https://doi.org/10.3390/microorganisms8121917
APA StyleLi, F., Duan, T., & Li, Y. (2020). Effects of the Fungal Endophyte Epichloë festucae var. lolii on Growth and Physiological Responses of Perennial Ryegrass cv. Fairway to Combined Drought and Pathogen Stresses. Microorganisms, 8(12), 1917. https://doi.org/10.3390/microorganisms8121917