Hidden Genetic Variability, Can the Olive Moth Prays oleae (Lepidoptera: Yponomeutidae or Praydidae?) be a Species’ Complex?


Abstract
1. Introduction
2. Materials and Methods
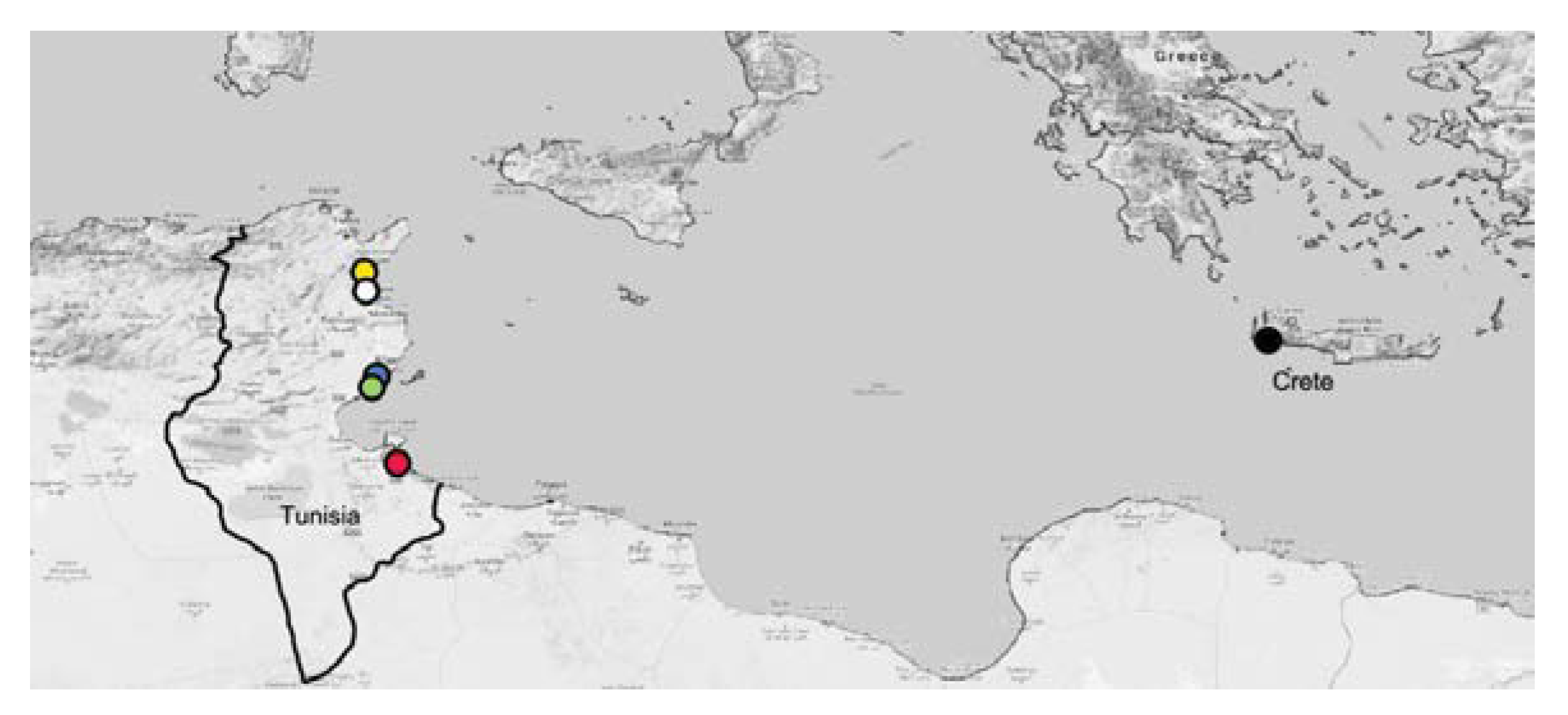
2.1. Specimen Collection
2.2. DNA Extraction, PCR and Sequencing
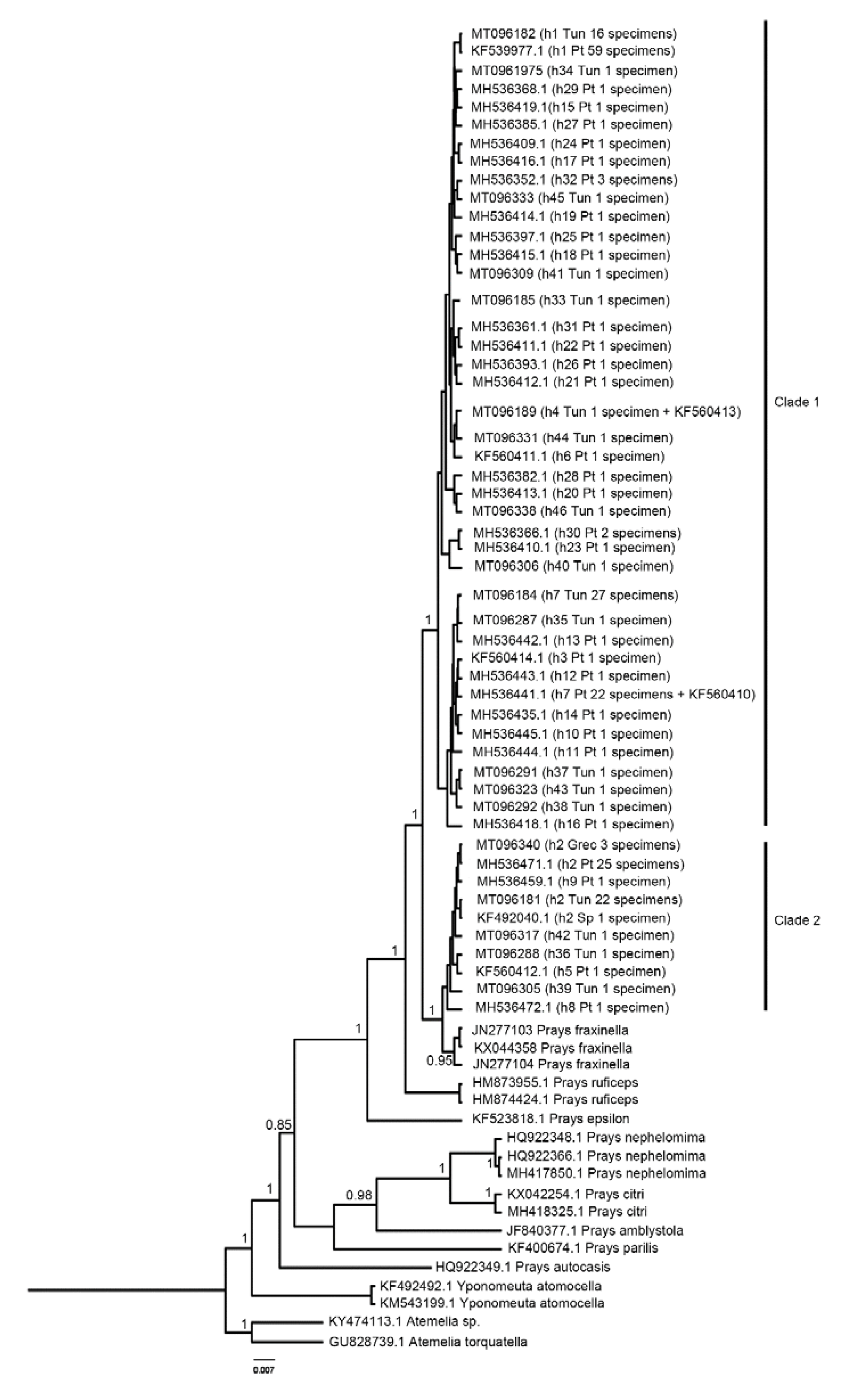
2.3. Phylogenetic Analysis
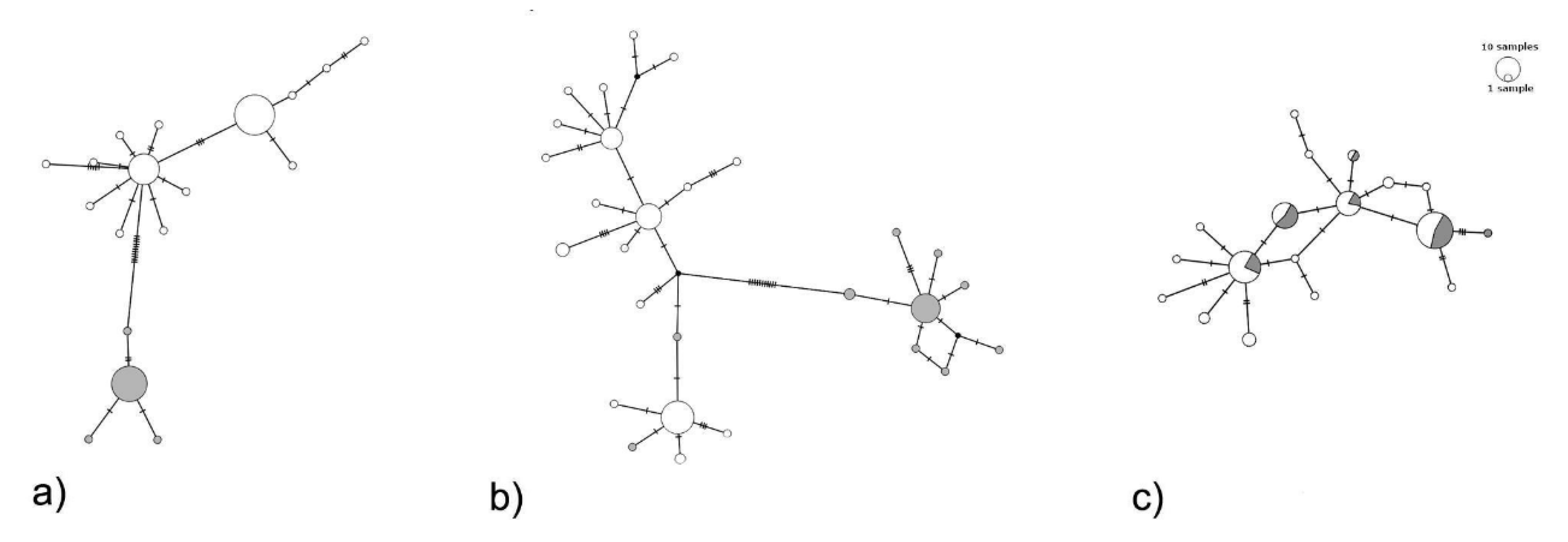
2.4. Variability and Population Structure
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nagaraju, J.; Gopinath, G.; Sharma, V.; Shukla, J.N. Lepidopteran Sex Determination: A Cascade of Surprises. Sex. Dev. 2014, 8, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Schepers, E.; Lum, A.; Rice, J.; Yalpani, N.; Gerber, R.; Jiménez-Juárez, N.; Haile, F.; Pascual, A.; Barry, J.; et al. Identification and Evaluations of Novel Insecticidal Proteins from Plants of the Class Polypodiopsida for Crop Protection against Key Lepidopteran Pests. Toxins 2019, 11, 383. [Google Scholar] [CrossRef] [PubMed]
- Ramos, P.; Campos, M.; Ramos, J.M. Long-term study on the evaluation of yield and economic losses caused by Prays oleae Bern. in the olive crop of Granada (southern Spain). Crop Prot. 1998, 17, 645–647. [Google Scholar] [CrossRef]
- Nobre, T.; Gomes, L.; Rei, F.T. Uncovered variability in olive moth (Prays oleae) questions species monophyly. PLoS ONE 2018, 13, e0207716. [Google Scholar] [CrossRef]
- Krenn, H.W. Feeding Mechanisms of Adult Lepidoptera: Structure, Function, and Evolution of the Mouthparts. Annu. Rev. Entomol. 2010, 55, 307–327. [Google Scholar] [CrossRef]
- Nardi, F.; Carapelli, A.; Boore, J.L.; Roderick, G.K.; Dallai, R.; Frati, F. Domestication of olive fly through a multi-regional host shift to cultivated olives: Comparative dating using complete mitochondrial genomes. Mol. Phylogenet. Evol. 2010, 57, 678–686. [Google Scholar] [CrossRef]
- Schimtt, T.; Seitz, A. Low diversity but high differentiation: The population genetics of Aglaope infausta (Zygaenidae: Lepidoptera). J. Biogeogr. 2004, 31, 137–144. [Google Scholar] [CrossRef]
- Whalberg, N.; Saccheri, I. The effects of Pleistocene glaciations on the phylogeography of Melitaea cinxia (Lepidoptera: Nymphalidae). Eur. J. Entomol. 2007, 104, 675–684. [Google Scholar] [CrossRef]
- Marí-Mena, N.; Lopez-Vaamonde, C.; Naveira, H.; Auger-Rozenberg, M.A.; Vila, M. Phylogeography of the Spanish Moon Moth Graellsia isabellae (Lepidoptera, Saturniidae). BMC Evol. Biol. 2016, 16, 139. [Google Scholar] [CrossRef]
- Segura, M.D.; Callejas, C.; Ochando, M.D. Bactrocera oleae: A single large population in Northern Mediterranean basin. J. Appl. Entomol. 2008, 132, 706–713. [Google Scholar] [CrossRef]
- van Asch, B.; Pereira-Castro, I.; Rei, F.; Costa, L.T. Mitochondrial haplotypes reveal olive fly (Bactrocera oleae) population substructure in the Mediterranean. Genetica 2012, 140, 181–187. [Google Scholar] [CrossRef][Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Wei, S.J.; Shi, B.C.; Gong, Y.J.; Jin, G.H.; Chen, X.X.; Meng, X.F. Genetic Structure and Demographic History Reveal Migration of the Diamondback Moth Plutella xylostella (Lepidoptera: Plutellidae) from the Southern to Northern Regions of China. PLoS ONE 2013, 8, e59654. [Google Scholar] [CrossRef] [PubMed]
- Wahlberg, N.; Wheat, C.W. Designing Novel Nuclear Markers for Genomic DNA Extractions of Lepidoptera. Syst. Biol. 2008, 57, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef]
- Larsson, H.; Källman, T.; Gyllenstrand, N.; Lascoux, M. Distribution of long-range linkage disequilibrium and Tajima’ s D values in scandinavian populations of Norway spruce (Picea abies). G3 (Bethesda) 2013, 3, 795–806. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar]
- Kelly, J.K. A test of neutrality based on interlocus associations. Genetics 1997, 146, 1197–1206. [Google Scholar]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Cywinska, A.; Hunter, F.F.; Hebert, P.D.N. Identifying Canadian mosquito species through DNA barcodes. Med. Vet. Entomol. 2006, 20, 413–424. [Google Scholar] [CrossRef]
- Linton, Y.; Lee, A.S.; Curtis, C. Discovery of a third member of the Maculipennis group in SW England. J. Eur. Mosq. Control Assoc. 2005, 19, 5–9. [Google Scholar]
- Hernández-Triana, L.M.; Chaverri, L.G.; Rodríguez-Pérez, M.A.; Prosser, S.W.J.; Hebert, P.D.N.; Gregory, T.R.; Johnson, N. DNA barcoding of Neotropical black flies (Diptera: Simuliidae): Species identification and discovery of cryptic diversity in Mesoamerica. Zootaxa 2015, 3936, 93–114. [Google Scholar] [CrossRef] [PubMed]
- Versteirt, V.Z.T.; Nagy, P.; Roelants, D.; Breman, F.C.; Damiens, D.; Dekoninck, W.; Backeljau, T.; Coosemans, M.; van Bortel, W. Identification of Belgian mosquito species (Diptera: Culicidae) by DNA barcoding. Mol. Ecol. Resour. 2015, 15, 449–457. [Google Scholar] [CrossRef]
- Tyagi, K.; Kumar, V.; Singha, D.; Chandra, K.; Laskar, B.A.; Kundu, S.; Chakraborty, R.; Chatterjee, S. DNA Barcoding studies on Thrips in India: Cryptic species and Species complexes. Sci. Rep. 2017, 7, 4898. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; de Waard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the Neotropical skipper butterfly Astrapesfulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef]
- Mandal, S.; Chhakchhuak, L.; Gurusubramanian, G.; Kumar, N.S. Mitochondrial markers for identification and phylogenetic studies in insects—A Review. DNA Barcodes 2014, 2, 1–9. [Google Scholar] [CrossRef]
- Madden, M.J.L.; Young, R.G.; Brown, J.W.; Miller, S.E.; Frewin, A.J.; Hanner, R.H. Using DNA barcoding to improve invasive pest identification at U.S. ports-of-entry. PLoS ONE 2019, 14, e0222291. [Google Scholar] [CrossRef]
- Mallet, J.; Beltrán, M.; Neukirchen, W.; Linares, M. Natural hybridization in heliconiine butterflies: The species boundary as a continuum. BMC Evol. Biol. 2007, 7, 28. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Hu, X.M.; Han, H.L.; Chen, F.; Cai, W.J.; Ruan, Q.Q.; Liu, B.; Luo, G.J.; Wang, H.; Liu, X.; et al. A two-step DNA barcoding approach for delimiting moth species: Moths of Dongling Mountain (Beijing, China) as a case study. Sci. Rep. 2018, 8, 14256. [Google Scholar] [CrossRef] [PubMed]
- Drovetski, S.V.; Raković, M.; Semenov, G.; Fadeev, I.V.; Red’kin, Y.A. Limited Phylogeographic Signal in Sex-Linked and Autosomal Loci Despite Geographically, Ecologically, and Phenotypically Concordant Structure of mtDNA Variation in the Holarctic Avian Genus Eremophila. PLoS ONE 2014, 9, e87570. [Google Scholar] [CrossRef] [PubMed]
- Dinca, V.; Balint, Z.; Voda, R.; Dapporto, L.; Hebert, P.D.N.; Vila, R. Use of genetic, climatic, and microbiological data to inform reintroduction of a regionally extinct butterfly. Conserv. Biol. 2018, 32, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Hickerson, M.J.; Meyer, C.P.; Moritz, C. DNA Barcoding Will Often Fail to Discover New Animal Species over Broad Parameter Space. Syst. Biol. 2006, 55, 729–739. [Google Scholar] [CrossRef]
- Hart, M.W.; Sunday, J. Things fall apart: Biological species form unconnected parsimony networks. Biol. Lett. 2007, 3, 509–512. [Google Scholar] [CrossRef]
- Kreuzinger, A.J.; Fiedler, K.; Letsch, H.; Grill, A. Tracing the radiation of Maniola (Nymphalidae) butterflies: New insights from phylogeography hint at one single incompletely differentiated species complex. Ecol. Evol. 2015, 5, 46–58. [Google Scholar] [CrossRef]
- Wahlberg, N.; Wheat, C.W.; Peña, C. Timing and Patterns in the Taxonomic Diversification of Lepidoptera (Butterflies and Moths). PLoS ONE 2013, 8, e80875. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Clades 1 and 2 | COI | nad5 | RpS5 | Concatenated |
|---|---|---|---|---|
| Number of sequences | 79 | 79 | 79 | 79 |
| Number of sites (bp) | 588 | 649 | 517 | 1754 |
| Number of haplotypes | 18 | 28 | 18 | 57 |
| Polymorphic sites (S) | 34 | 42 | 16 | 92 |
| Parsimony informative | 17 | 24 | 8 | 49 |
| Total number of mutations | 36 | 47 | 16 | 99 |
| Haplotype diversity (Hd) | 0.779 | 0.892 | 0.846 | 0.986 |
| Aver. nucleotide diff. (k) | 7.166 | 7.886 | 2.024 | 17.07 |
| Nucleotide diversity (Pi) | 0.012 | 0.012 | 0.003 | 0.009 |
| Clade 1 | COI | nad5 | RpS5 | Concatenated |
| Number of sequences | 55 | 55 | 55 | 55 |
| Number of sites (bp) | 588 | 649 | 517 | 1754 |
| Number of haplotypes | 14 | 18 | 17 | 45 |
| Polymorphic sites (S) | 21 | 29 | 14 | 83 |
| Parsimony informative | 5 | 13 | 8 | 46 |
| Total number of mutations | 21 | 33 | 14 | 88 |
| Haplotype diversity (Hd) | 0.683 | 0.838 | 0.874 | 0.987 |
| Aver. nucleotide diff. (k) | 2.261 | 3.521 | 2.149 | 9.237 |
| Nucleotide diversity (Pi) | 0.003 | 0.005 | 0.004 | 0.005 |
| Clade 2 | COI | nad5 | RpS5 | Concatenated |
| Number of sequences | 24 | 24 | 24 | 24 |
| Number of sites (bp) | 588 | 649 | 517 | 1754 |
| Number of haplotypes | 4 | 10 | 6 | 14 |
| Polymorphic sites (S) | 4 | 25 | 7 | 34 |
| Parsimony informative | 0 | 16 | 3 | 18 |
| Total number of mutations | 4 | 25 | 7 | 34 |
| Haplotype diversity (Hd) | 0.239 | 0.667 | 0.757 | 0.92 |
| Aver. nucleotide diff. (k) | 0.333 | 3.431 | 1.648 | 5.17 |
| Nucleotide diversity (Pi) | 0 | 0.004 | 0.003 | 0.002 |
| Clades 1 and 2 | ZnS | Significance | Tajima’s D | Significance |
|---|---|---|---|---|
| COI | 0.17 | p = 0.96; [0.04, 0.17] | −0.05 | p = 0.49; [−1.32, 1.33] |
| nad5 | 0.11 | p = 0.78; [0.04, 0.17] | −0.55 | p = 0.24; [−1.32, 1.42] |
| RpS5 | 0.04 | p = 0.20; [0.00, 0.25] | −1.08 | p = 0.10; [−1.52, 1.55] |
| Concatenated | 0.08 | p = 0.53; [0.04, 0.15] | −0.50 | p = 0.23; [−1.28, 1.23] |
| Clade 1 | ZnS | Significance | Tajima’s D | Significance |
| COI | 0.09 | p = 0.47; [0.01, 0.26] | −1.60 | p = 0.009; [−1.45, 1.51] |
| nad5 | 0.07 | p = 0.23; [0.03, 0.23] | −1.69 | p = 0.005; [−1.46, 1.63] |
| RpS5 | 0.04 | p = 0.17; [0.01, 0.29] | −0.89 | p = 0.14; [−1.57, 1.72] |
| Concatenated | 0.10 | p = 0.59; [0.05, 0.18] | −1.80 | p = 0.00; [−1.30, 1.27] |
| Clade 2 | ZnS | Significance | Tajima’s D | Significance |
| COI | 0.17 | p = 0.70; [0.03, 0.60] | −1.88 | p = 0.00; [−1.51, 1.71] |
| nad5 | 0.31 | p = 0.95; [0.05, 0.35] | −1.81 | p = 0.01; [−1.56, 1.38] |
| RpS5 | 0.21 | p = 0.77; [0.00, 0.45] | −0.37 | p = 0.34; [−1.57, 1.67] |
| Concatenated | 0.20 | p = 0.81; [0.06, 0.30] | −1.64 | p = 0.01; [−1.51, 1.39] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pazian, M.; Nobre, T.; Blibech, I.; Rei, F.T. Hidden Genetic Variability, Can the Olive Moth Prays oleae (Lepidoptera: Yponomeutidae or Praydidae?) be a Species’ Complex? Insects 2020, 11, 204. https://doi.org/10.3390/insects11040204
Pazian M, Nobre T, Blibech I, Rei FT. Hidden Genetic Variability, Can the Olive Moth Prays oleae (Lepidoptera: Yponomeutidae or Praydidae?) be a Species’ Complex? Insects. 2020; 11(4):204. https://doi.org/10.3390/insects11040204
Chicago/Turabian StylePazian, Marlon, Tânia Nobre, Imen Blibech, and Fernando T Rei. 2020. "Hidden Genetic Variability, Can the Olive Moth Prays oleae (Lepidoptera: Yponomeutidae or Praydidae?) be a Species’ Complex?" Insects 11, no. 4: 204. https://doi.org/10.3390/insects11040204
APA StylePazian, M., Nobre, T., Blibech, I., & Rei, F. T. (2020). Hidden Genetic Variability, Can the Olive Moth Prays oleae (Lepidoptera: Yponomeutidae or Praydidae?) be a Species’ Complex? Insects, 11(4), 204. https://doi.org/10.3390/insects11040204