Molecular Characterization of Neoseiulus barkeri Vitellogenin Genes and Vitellogenin Receptor during Reproductive Diapause

Abstract
1. Introduction
2. Materials and Methods
2.1. Source of Tested Insects
2.2. Collection of Insects
2.3. Developmental Period of N. barkeri under Diapause and Non-Diapause Conditions
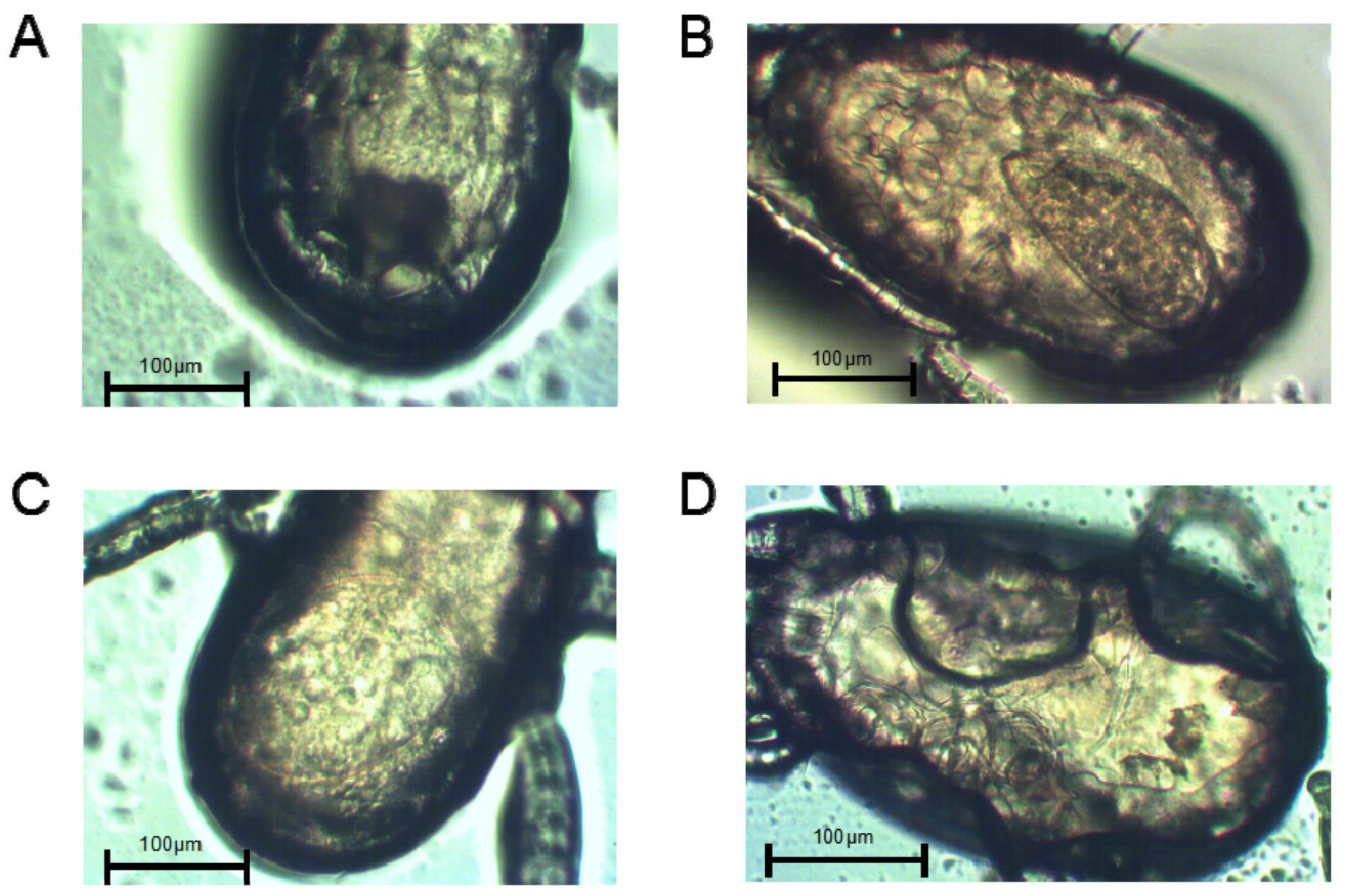
2.4. Abdominal Morphology of Female Adult Mites
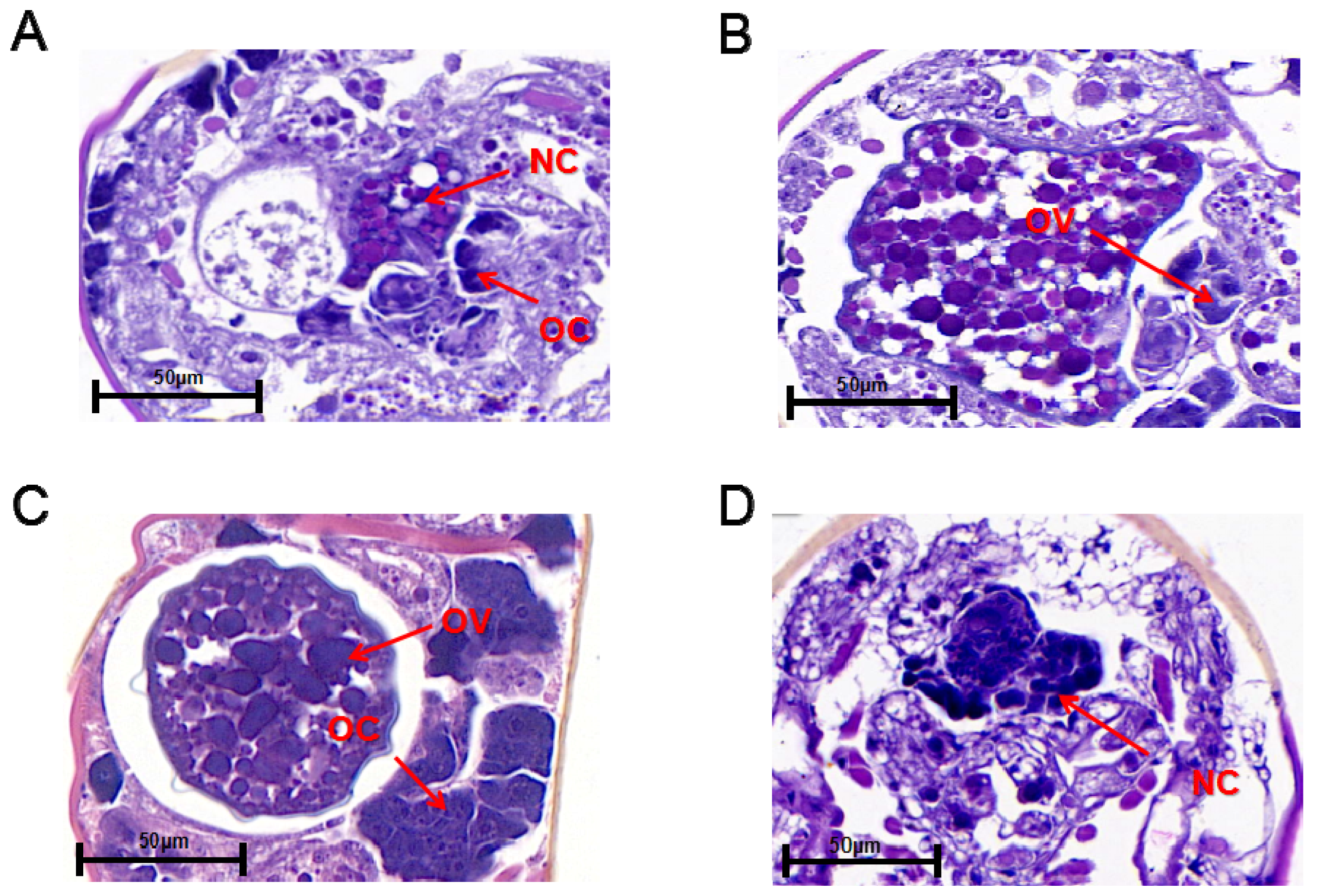
2.5. Ovarian Section
2.6. RNA Extraction and cDNA Synthesis
2.7. Bioinformatics Analyses
2.8. Quantitative Real-Time Polymerase Chain Reaction (PCR)
2.9. Statistical Analysis
3. Results
3.1. Developmental Periods of N. barkeri under Diapause and Non-Diapause Conditions
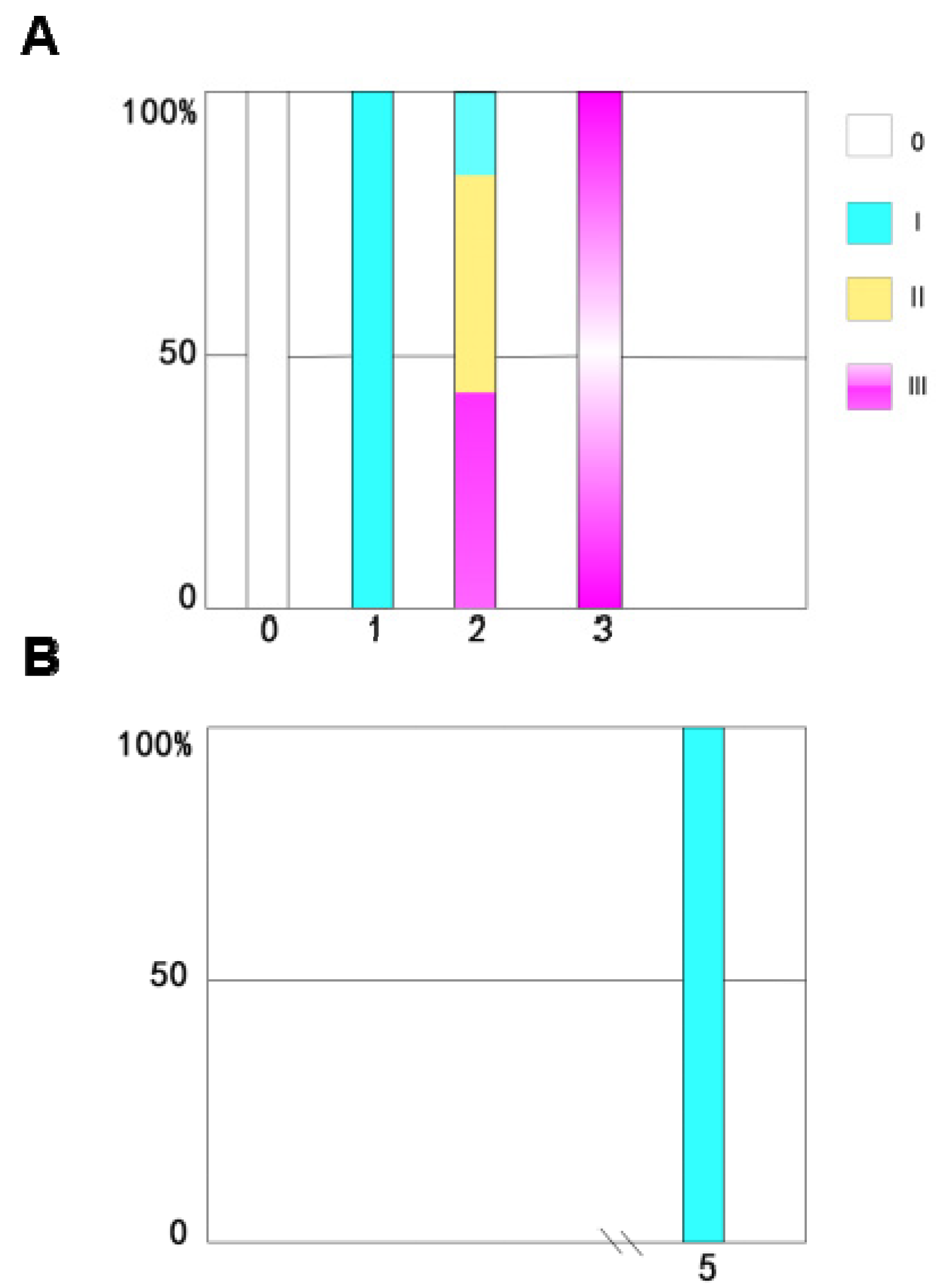
3.2. Analysis of Ovarian Development
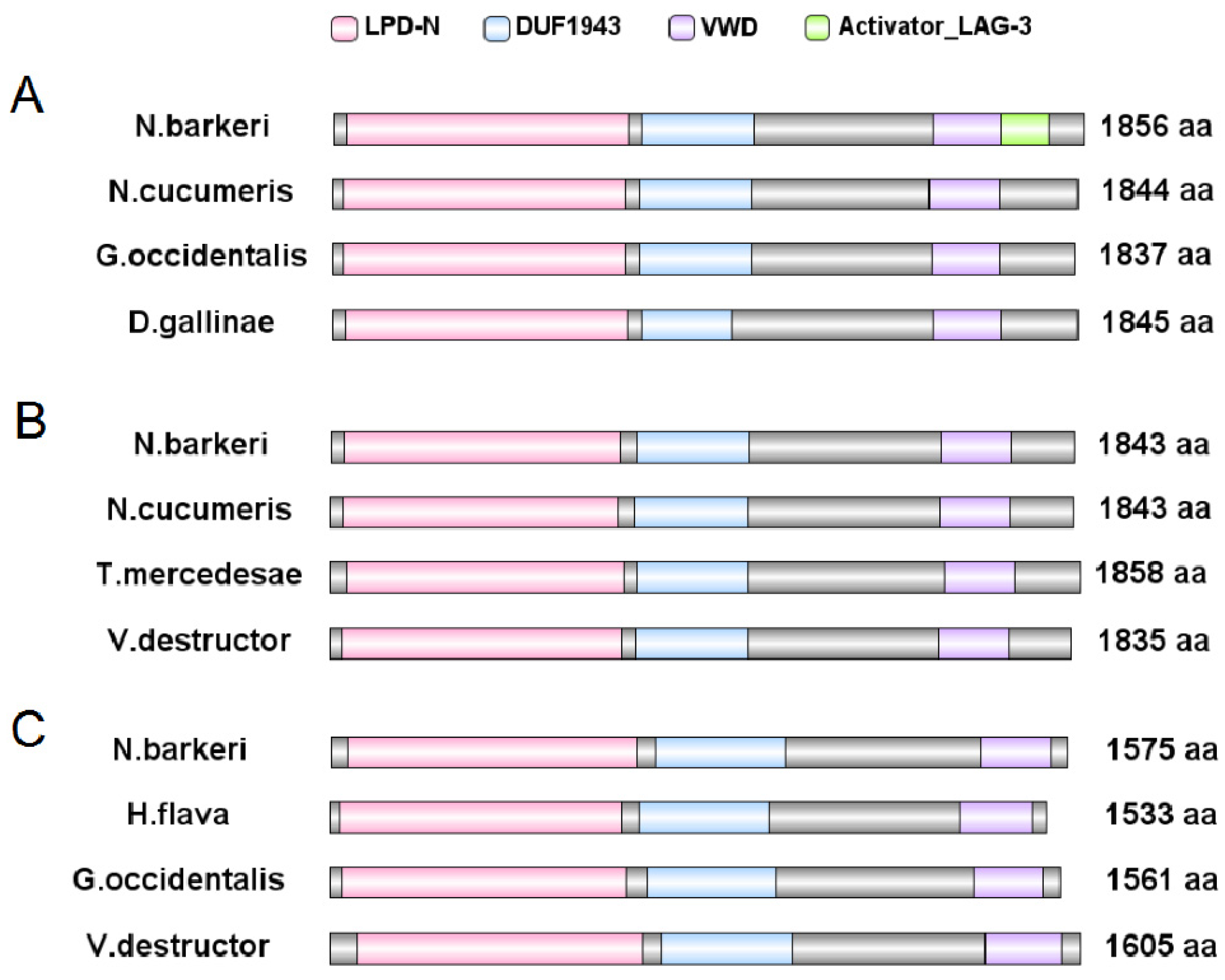
3.3. Bioinformatics Analysis
3.3.1. Bioinformatic Analysis of Vgs
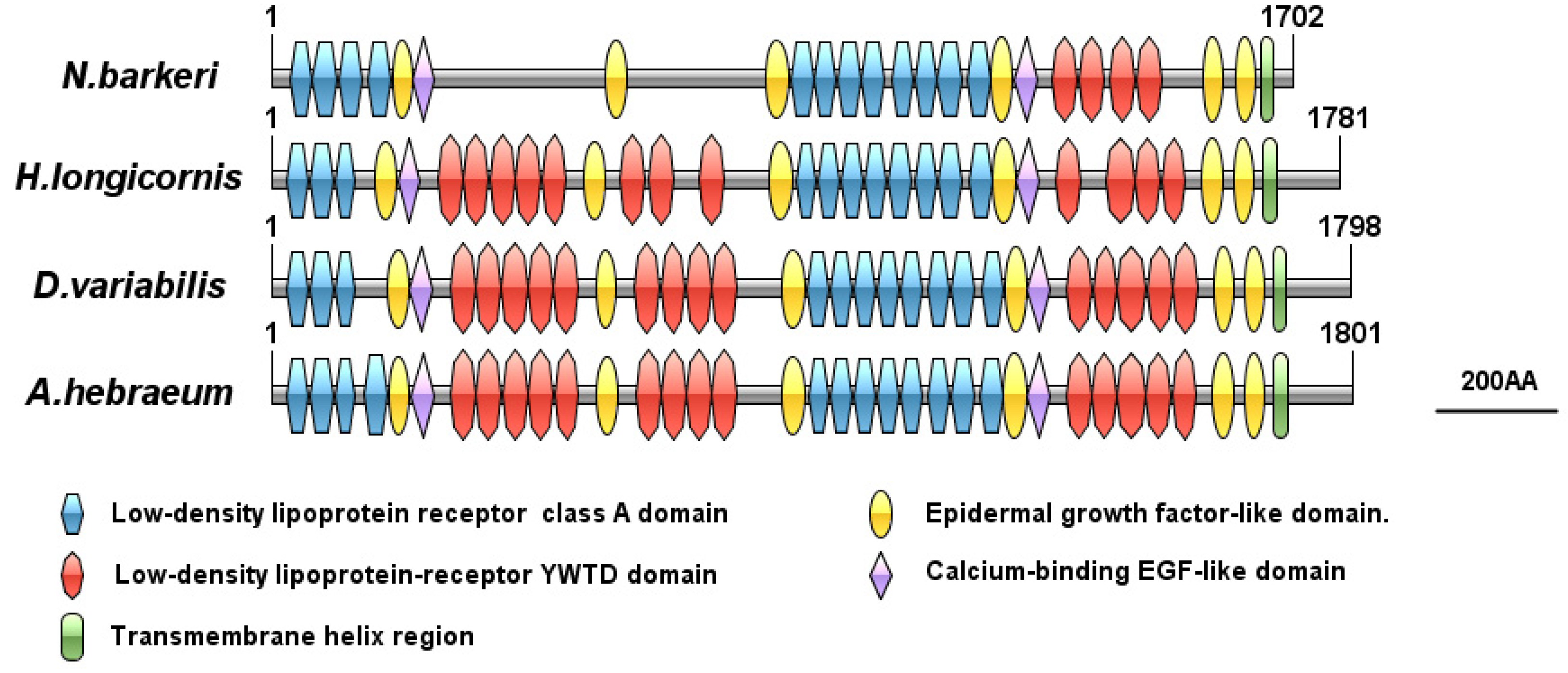
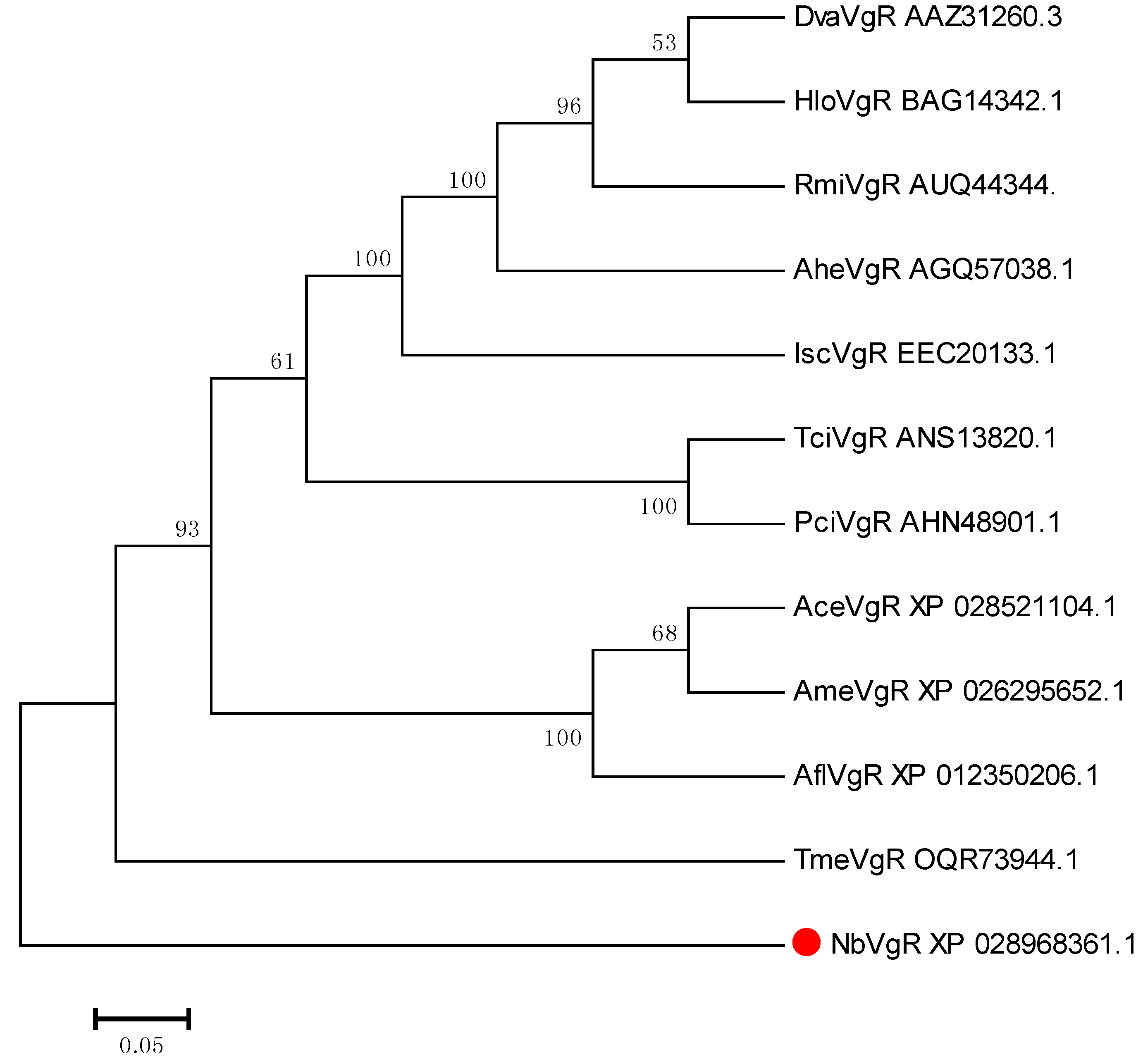
3.3.2. VgR Bioinformatics Analysis
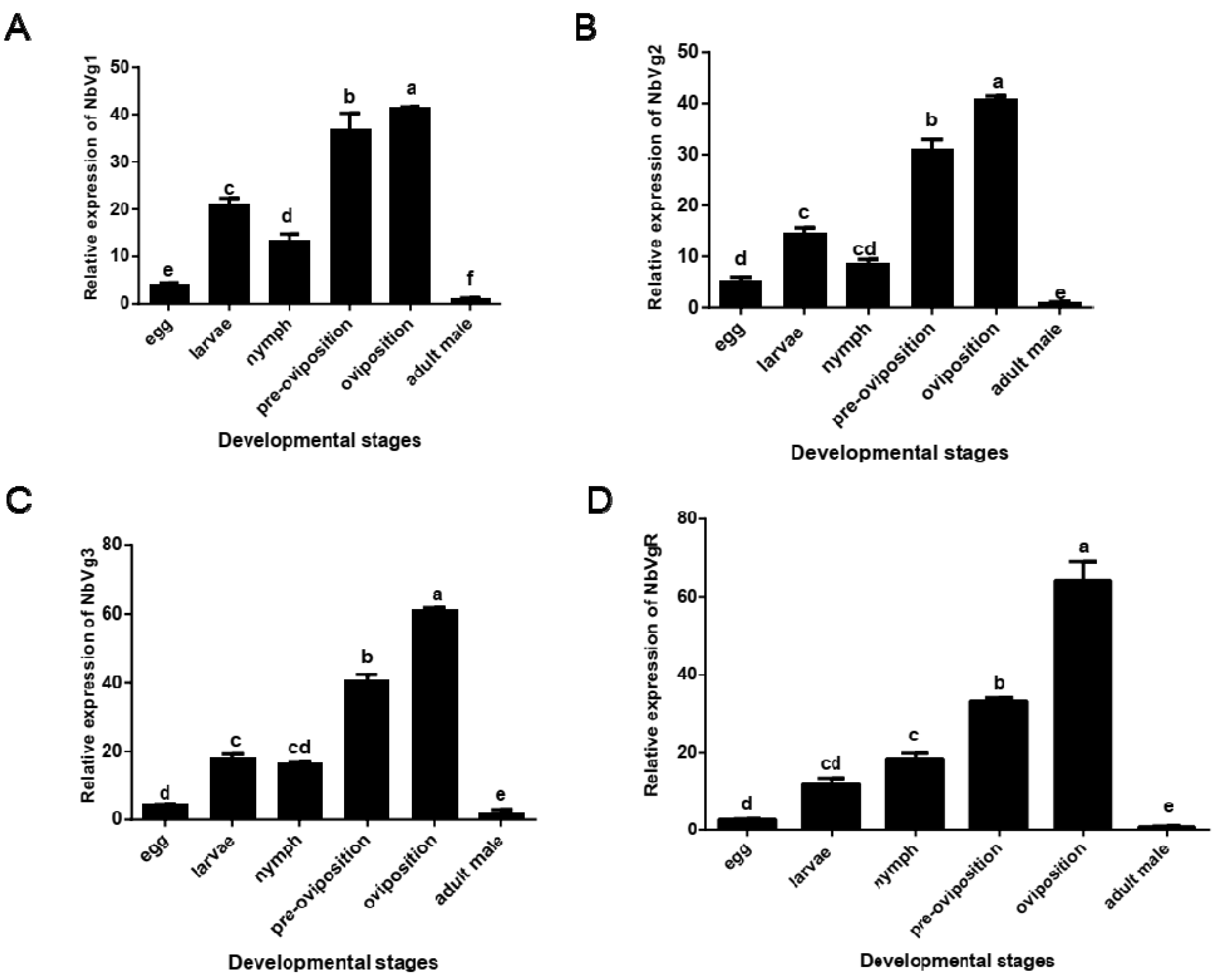
3.3.3. Transcriptional Expression of the NbVgs and NbVgR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hessein, N.A.; Parrella, M.P. Predatory mites help control thrips on floriculture crops. Calif. Agric. 1991, 44, 19–21. [Google Scholar] [CrossRef]
- Grafton Cardwell, E.E.; Ouyang, Y.; Striggow, R.A. Predacious mites for control of citrus thrips, Scirtothrips citri (Thysanoptera: Thripidae) in nursery citrus. Biol. Control 1999, 14, 29–36. [Google Scholar] [CrossRef]
- Nomikou, M.; Janssen, A.; Schraag, R.; Sabelis, W. Phytoseiid predators as potential biological control agents for Bemisia tabaci. Exp. Appl. Acarol. 2001, 25, 271–291. [Google Scholar] [CrossRef]
- Fernando, L.C.P.; Waidyarathne, K.P.; Perera, K.F.G.; Silva, P.H.P.R.D. Evidence for suppressing coconut mite, Aceria guerreronis by inundative release of the predatory mite, Neoseiulus baraki. Biol. Control 2010, 53, 108–111. [Google Scholar] [CrossRef]
- Endong, W.; Xuenong, X.; Shengyong, W. Control effects of Amblyseius barkeri on Frankliniella occidentalis on the eggplants and their natural enemy Orius sauteri in the greenhouse. Plant Prot. 2010, 36, 101–104. [Google Scholar]
- Xia, B.; Zou, Z.; Li, P.; Lin, P. Effect of temperature on development and reproduction of Neoseiulus barkeri (acari: Phytoseiidae) fed on aleuroglyphus ovatus. Exp. Appl. Acarol. 2012, 56, 33–41. [Google Scholar] [CrossRef]
- Prestwich, G.D. Comprehensive insect physiology, biochemistry, and pharmacology. Int. J. Biochem. 1985, 17, 1282–1283. [Google Scholar] [CrossRef]
- Gotoh, T. Annual Life Cycle of the Two-Spotted Spider Mite, Tetranychus urticae KOCH (Acarina: Tetranychidae), on Ribes rubrum L. in Sapporo: The Presence of Non-Diapausing Individuals. Appl. Entomol. Zool. 1986, 21, 454–460. [Google Scholar] [CrossRef]
- Wysoki, M. Studies on diapause and the resistance to low temperatures of a predacious mite, Phytoseius finimus (mesostigmata, phytoseiidae). Eniomol. Exp. Appl. 1974, 17, 22–30. [Google Scholar] [CrossRef]
- Croft, B.A. Comparative Studies on Four Strains of Typhlodromus occidentalis (Acarina: Phytoseiidae). V. Photoperiodic Induction of Diapause. Ann. Entomol. Soc. Am. 1971, 64, 962–964. [Google Scholar] [CrossRef]
- Lu, K.; Shu, Y.H.; Zhou, J.L.; Zhang, X.Y.; Zhang, X.Y.; Chen, M.X.; Yao, Q.; Zhou, Q.; Zhang, W.Q. Molecular characterization and RNA interference analysis of vitellogenin receptor from Nilaparvata lugens. J. Insect Physiol. 2015, 73, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Tufail, M.; Takeda, M. Molecular characteristics of insect vitellogenins. J. Insect Physiol. 2008, 54, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- Sappington, T.W.; Raikhel, A.S. Molecular characteristics of insect vitellogenins and vitellogenin receptors. Insect Biochem. Mol. Biol. 1998, 28, 277–300. [Google Scholar] [CrossRef]
- Huo, Y.; Liu, W.W.; Zhang, F.J.; Chen, X.Y.; Li, L.; Liu, Q.F.; Zhou, Y.J.; Wei, T.Y.; Fang, R.X.; Wang, X.F. Transovarial transmission of a plantvirus is mediated by vitellogen of its insect vector. PLoS Pathog. 2014, 10, e1003949. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.Q.; Wu, J.C.; Zhao, K.F.; Chen, Y.; Yang, G.Q. Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvata lugens (Stål) (hemiptera: Delphacidae) adult females and males exposed to two insecticides. Pestic. Biochem. Physiol. 2010, 98, 269–278. [Google Scholar] [CrossRef]
- Schip, F.D.V.H.; Samallo, J.; Broos, J.; Ophuis, J.; Mojet, M.; Gruber, M.; Geert, A.B. Nucleotide sequence of a chicken vitellogenin gene and derived amino acid sequence of the encoded yolk precursor protein. J. Mol. Biol. 1987, 196, 245–260. [Google Scholar] [CrossRef]
- Wahli, W.; Dawid, I.B.; Ryffel, G.U.; Wyler, T.; Jaggi, R.B.; Weber, R. Vitellogenin in Xenopus laevis is encoded in a small family of genes. Cell 1979, 16, 535–549. [Google Scholar] [CrossRef]
- Kawakami, Y.; Goto, S.G.; Ito, K.; Numata, H. Suppression of ovarian development and vitellogenin gene expression in the adult diapause of the two-spotted spider mite Tetranychus urticae. J. Insect Physiol. 2009, 55, 70–77. [Google Scholar] [CrossRef]
- Blumenthal, T.; Squire, M.; Kirtland, S.; Cane, J.; Donegan, M.; Spieth, J.; Sharrock, W. Cloning of a yolk protein gene family from Caenorhabditis elegans. J. Mol. Biol. 1984, 174, 1–18. [Google Scholar] [CrossRef]
- Sappington, T.W.; Raikhel, A.S. Insect vitellogenin/yolk protein receptors. Reprod. Biol. Invertebr. 2005, 229–264. [Google Scholar]
- Okabayashi, K.; Shoji, H.; Nakamura, T.; Hashimoto, O.; Asashima, M.; Sugino, H. cDNA cloning and expression of the Xenopus laevis vitellogenin receptor. Biochem. Biophys. Res. Commun. 1996, 224, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Raikhel, A.S.; Dhadialla, T.S. Accumulation of yolk proteins in insect oocytes. Annu. Rev. Entonzol. 1992, 37, 217–251. [Google Scholar] [CrossRef]
- Zhong, R.; Ding, T.B.; Niu, J.Z.; Xia, W.K.; Liao, C.Y.; Dou, W.; Wang, J.J. Molecular characterization of vitellogenin and its receptor genes from citrus red mite, Panonychus citri (McGregor). Int. J. Mol. Sci. 2015, 16, 4759–4773. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Chen, F.; Luo, R.; Pan, Q.; Wang, C.; Yu, S.; Cong, L.; Liu, H.; Li, H.; Ran, C. Gene cloning and difference analysis of vitellogenin in Neoseiulus barkeri (Hughes). Bull. Entomol. Res. 2018, 108, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Y.; Zhang, C.X. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect Sci. 2013, 20, 254–260. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Veerman, A. Diapause in phytoseiid mites: A review. Exp. Appl. Acarol. 1992, 14, 1–60. [Google Scholar] [CrossRef]
- Shikina, S.; Chen, C.J.; Chung, Y.J.; Shao, Z.F.; Liou, J.Y.; Tseng, H.P.; Lee, Y.H.; Chang, C.F. Yolk formation in a stony coral Euphyllia ancora (Cnidaria, Anthozoa): Insight into the evolution of vitellogenesis in nonbilaterian animals. Endocrinology 2013, 154, 3447–3459. [Google Scholar] [CrossRef]
- Alexander, D.S.; Kaufman, W.R. Molecular characterization of the vitellogenin receptor from the tick, Amblyomma hebraeum (Acari: Ixodidae). Insect Biochem. Mol. Biol. 2014, 5, 821–833. [Google Scholar]
- Chippendale, G.M.; Yin, C.M. Endocrine interactions controlling the larval diapause of the southwestern corn borer, Diatraea grandiosella. J. Insect Physiol. 1976, 22, 989–995. [Google Scholar] [CrossRef]
- Lin, C.; Yang, W.J.; Jiang, X.Z.; Niu, J.Z.; Shen, G.M.; Ran, C.; Wang, J.J. The Essential Role of Vitellogenin Receptor in Ovary Development and Vitellogenin Uptake in Bactrocera dorsalis (Hendel). Int. J. Mol. Sci. 2015, 16, 18368–18383. [Google Scholar]
- Taylor, D.; Chinzei, Y.; Miura, K.; Ando, K. Vitellogenin synthesis, processing and hormonal regulation in the tick, Ornithodoros parkeri (Acari: Argasidae). Insect Biochem. 1991, 21, 723–733. [Google Scholar] [CrossRef]
- Qian, C.; Fu, W.W.; Wei, G.Q.; Wang, L.; Liu, Q.N.; Dai, L.S.; Sun, Y.; Zhu, B.J.; Liu, C.L. Identification and expression analysis of vitellogenin receptor from the Wild Silkworm, Bombyx mandarina. Arch. Insect Biochem. Physiol. 2015, 89, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Mao, J.; Zeng, F. Chrysopa septempunctata (Neuroptera: Chrysopidae) Vitellogenin Functions through Effects on Egg Production and Hatching. J. Econ. Entomol. 2015, 108, 2779–2788. [Google Scholar] [CrossRef] [PubMed]
- Guidugli-Lazzarini, K.R.; Nascimento, A.M.D.; Tanaka, E.D.; Piulachs, M.D.; Hartfelder, K.; Márcia, G.B.; Paulino, L.Z.S. Expression analysis of putative vitellogenin and lipophorin receptors in honey bee (Apis mellifera L.) queens and workers. J. Insect Physiol. 2008, 54, 1138–1147. [Google Scholar] [CrossRef]
- Kawakami, Y.; Numata, H. Effects of a Pyrethroid on Ovarian Development in Diapause Females of the Two Spotted Spider Mite. J. Acarol. Soc. Jpn. 2013, 22, 45–47. [Google Scholar] [CrossRef][Green Version]
- Okumura, T.; Yamano, K.; Sakiyama, K. Vitellogenin gene expression and hemolymph vitellogenin during vitellogenesis, final maturation, and oviposition in female kuruma prawn, Marsupenaeus japonicus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 147, 1028–1037. [Google Scholar] [CrossRef]
- Avarre, J.C.; Michelis, R.; Tietz, A.; Lubzens, E. Relationship between vitellogenin and vitellin in a marine shrimp (Penaeus semisulcatus) and molecular characterization of vitellogenin complementary DNAs. Biol. Reprod. 2003, 69, 355–364. [Google Scholar] [CrossRef]
- Liu, X.; Shen, G.; Xu, H.; He, L. The fenpropathrin resistant Tetranychus cinnabarinus showed increased fecundity with high content of vitellogenin and vitellogenin receptor. Pestic. Biochem. Physiol. 2016, 134, 31–38. [Google Scholar] [CrossRef]
- Schonbaum, C.P.; Perrino, J.J.; Mahowald, A.P. Regulation of the vitellogenin receptor during Drosophila melanogaster oogenesis. Mol. Biol. Cell 2000, 11, 511–521. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′–3′) | Product Size (bp) |
|---|---|---|
| NbVg1-qF | CGACATTCCCATCTACGC | 155 |
| NbVg1-qR | AACCTTCCTGCTCCTTACC | |
| NbVg2-qF | CGCCAGGAAGAAGGTATC | 187 |
| NbVg2-qR | TGTGAGTGGGGCAAACG | |
| NbVg3-qF | GCCAGCGACCAACAGAT | 177 |
| NbVg3-qR | GCGGAAGCAAGGGTAAT | |
| NbVgR-qF | CACAAGAGGGCGAAGAGC | 117 |
| NbVgR-qR | CCATCGGAGCAGAGTCAAG | |
| NbActin-qF | TCAGCGATGTCAGTTTGAGG | 103 |
| NbActin-qR | CCTCCTCTCGCAATGAGAAC |
| Protein | Amino Acids | Molecular Weight (kDa) | Theoretical pI |
|---|---|---|---|
| NbVg1 | 1856 | 212 | 8.61 |
| NbVg2 | 1843 | 211 | 8.98 |
| NbVg3 | 1575 | 179 | 6.88 |
| NbVgR | 1702 | 191 | 5.53 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, J.; Zhang, Y.; Ma, L.; Niu, T.; Dong, T.; Sheng, R.; Li, L.; Xu, Y.; Xi, L.; Li, G. Molecular Characterization of Neoseiulus barkeri Vitellogenin Genes and Vitellogenin Receptor during Reproductive Diapause. Insects 2020, 11, 203. https://doi.org/10.3390/insects11040203
Jiang J, Zhang Y, Ma L, Niu T, Dong T, Sheng R, Li L, Xu Y, Xi L, Li G. Molecular Characterization of Neoseiulus barkeri Vitellogenin Genes and Vitellogenin Receptor during Reproductive Diapause. Insects. 2020; 11(4):203. https://doi.org/10.3390/insects11040203
Chicago/Turabian StyleJiang, Junqi, Ying Zhang, Lei Ma, Tingting Niu, Tingting Dong, Ruirui Sheng, Ling Li, Yeyu Xu, Lingyu Xi, and Guiting Li. 2020. "Molecular Characterization of Neoseiulus barkeri Vitellogenin Genes and Vitellogenin Receptor during Reproductive Diapause" Insects 11, no. 4: 203. https://doi.org/10.3390/insects11040203
APA StyleJiang, J., Zhang, Y., Ma, L., Niu, T., Dong, T., Sheng, R., Li, L., Xu, Y., Xi, L., & Li, G. (2020). Molecular Characterization of Neoseiulus barkeri Vitellogenin Genes and Vitellogenin Receptor during Reproductive Diapause. Insects, 11(4), 203. https://doi.org/10.3390/insects11040203