In Vitro Antioxidant and Anti-Neuroinflammatory Effects of Elsholtzia blanda (Benth.) Benth.


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Plant Material and Identification
2.3. Cell Culture and Viability
2.4. NO Measurements
2.5. Intracellular ROS Assay
2.6. IL-6 and TNF-α Production
2.7. Western Blot Analysis
2.8. Total RNA Extraction
2.9. Real-Time Reverse Transcription–Polymerase Chain Reaction
2.10. Statistical Analysis
3. Results
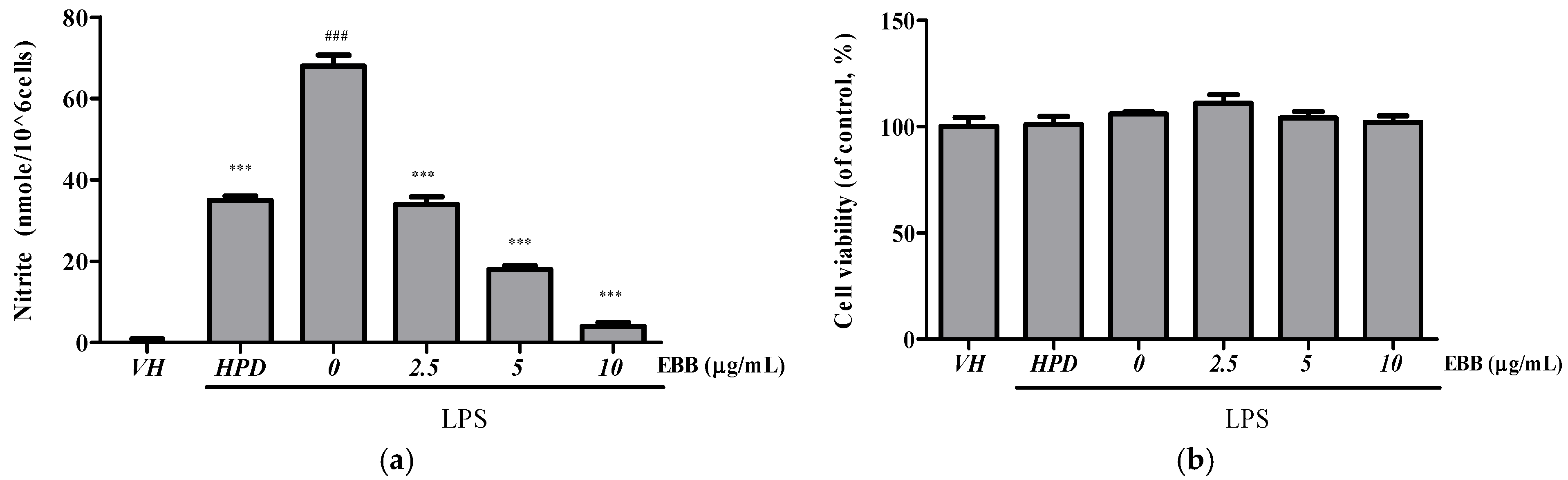
3.1. Effects of EBB on NO Production and Cell Viability in LPS-Stimulated BV2 Cells
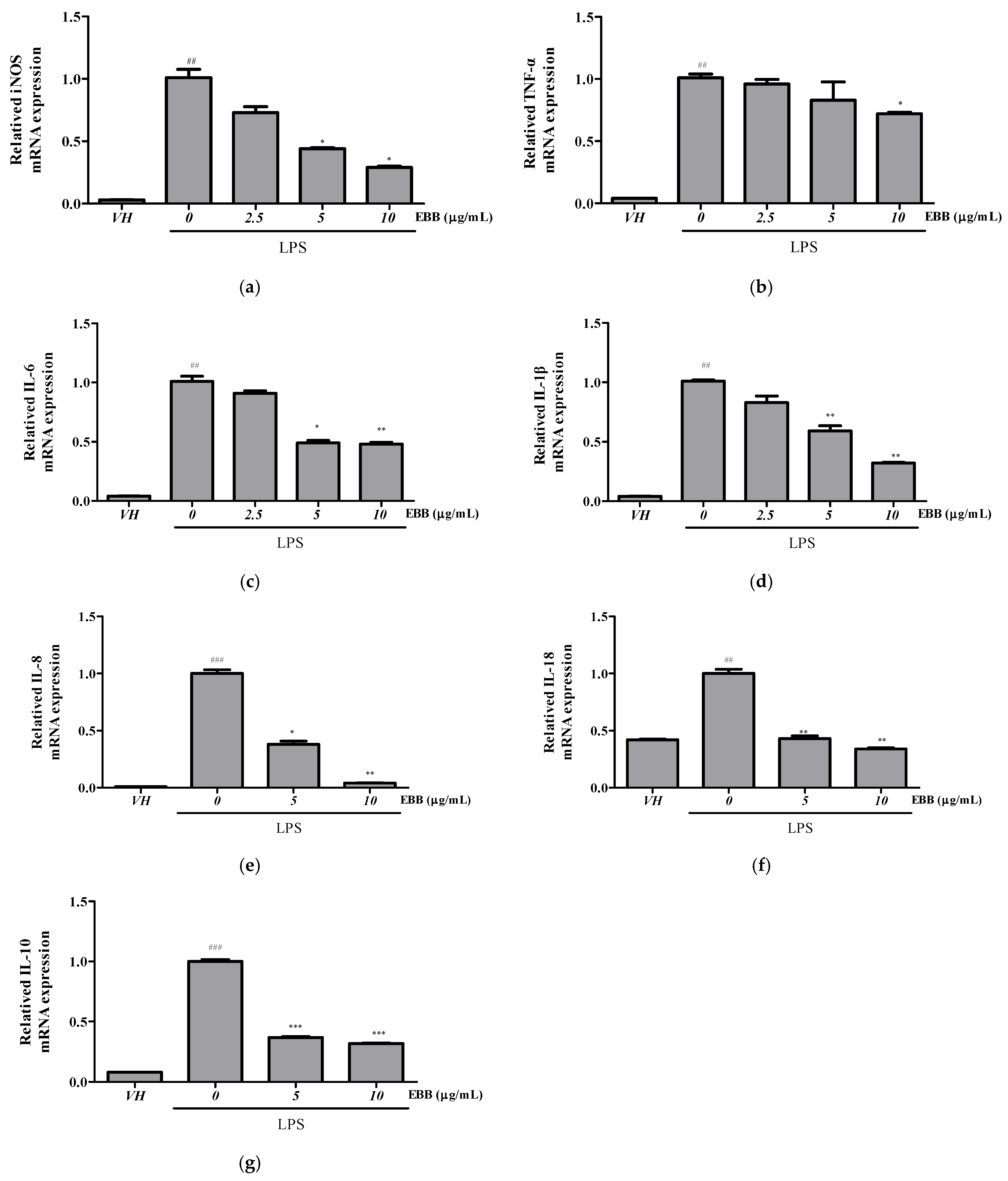
3.2. Effects of EBB on Cytokine Production and mRNA Expression in LPS-Stimulated BV2 Cells
3.3. Effects of EBB on Inflammatory-Related Proteins of LPS-Stimulated BV2 Cells
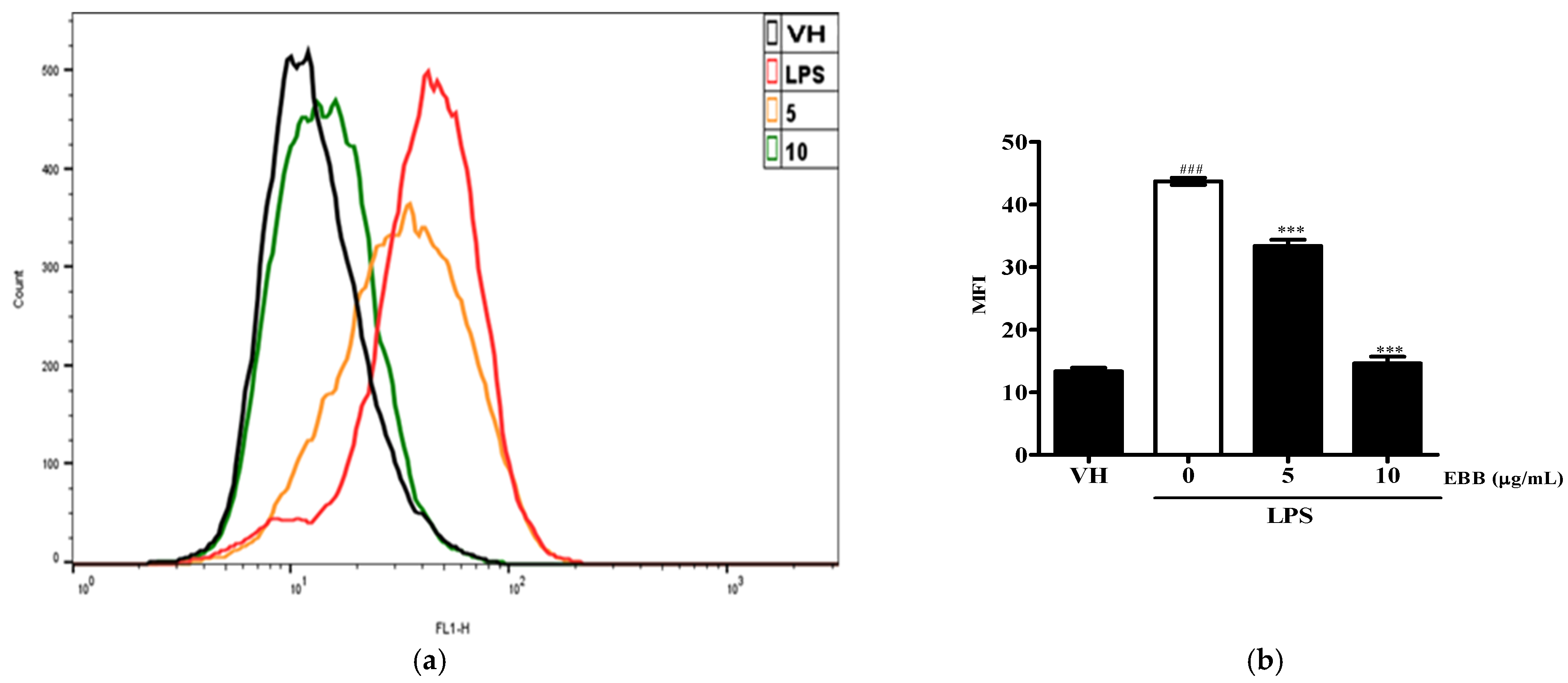
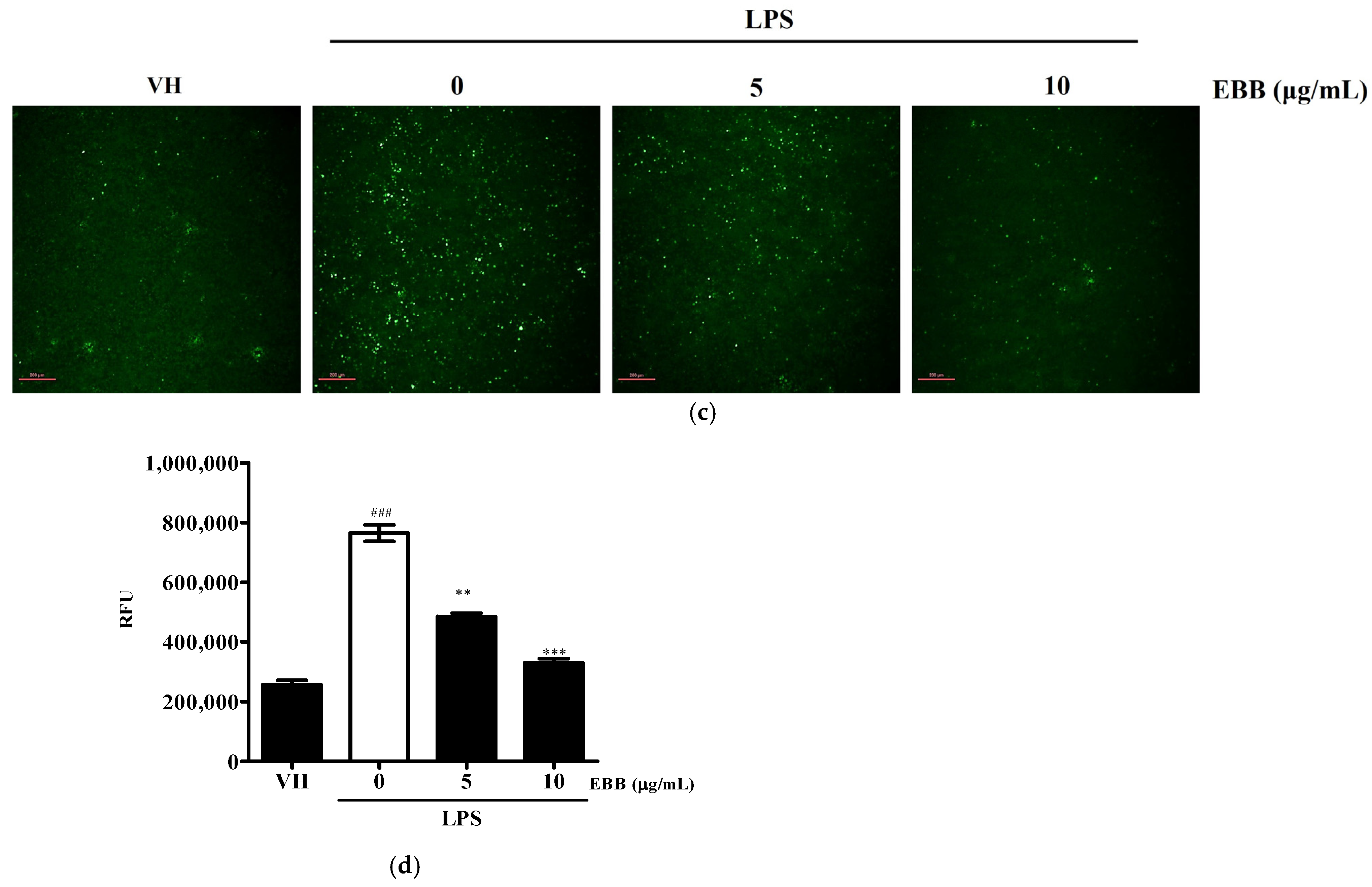
3.4. Effects of EBB on ROS Production of LPS-Treated Microglial Cells
3.5. Anti-Inflammatory Effects of EBB Are Mediated by the HO-1 Signaling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, C.; Jiang, J.; Tan, Y.; Chen, S. Microglia in Neurodegenerative Diseases: Mechanism and Potential Therapeutic Targets. Signal Transduct. Target. Ther. 2023, 8, 359. [Google Scholar] [CrossRef]
- Kim, Y.S.; Joh, T.H. Microglia, Major Player in the Brain Inflammation: Their Roles in the Pathogenesis of Parkinson’s Disease. Exp. Mol. Med. 2006, 38, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Henn, A.; Lund, S.; Hedtjärn, M.; Schrattenholz, A.; Pörzgen, P.; Leist, M. The Suitability of BV2 Cells as Alternative Model System for Primary Microglia Cultures or for Animal Experiments Examining Brain Inflammation. ALTEX 2009, 26, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.A.; Martin, D.; Manuelidis, L. Microglia from Creutzfeldt-Jakob Disease-Infected Brains Are Infectious and Show Specific MRNA Activation Profiles. J. Virol. 2002, 76, 10905–10913. [Google Scholar] [CrossRef] [PubMed]
- Wang-Sheng, C.; Jie, A.; Jian-Jun, L.; Lan, H.; Zeng-Bao, X.; Chang-Qing, L. Piperine Attenuates Lipopolysaccharide (LPS)-Induced Inflammatory Responses in BV2 Microglia. Int. Immunopharmacol. 2017, 42, 44–48. [Google Scholar] [CrossRef]
- Cho, H.; Kim, D.-U.; Oh, J.-Y.; Park, S.-J.; Kweon, B.; Bae, G.-S. Anti-Neuroinflammatory Effects of Arecae Pericarpium on LPS-Stimulated BV2 Cells. Curr. Issues Mol. Biol. 2024, 46, 884–895. [Google Scholar] [CrossRef]
- Kweon, B.; Oh, J.; Lim, Y.; Noh, G.; Yu, J.; Kim, D.; Jang, M.; Kim, D.; Bae, G. Anti-Inflammatory Effects of Honeysuckle Leaf Against Lipopolysaccharide-Induced Neuroinflammation on BV2 Microglia. Nutrients 2024, 16, 3954. [Google Scholar] [CrossRef]
- Nam, H.Y.; Nam, J.H.; Yoon, G.; Lee, J.-Y.; Nam, Y.; Kang, H.-J.; Cho, H.-J.; Kim, J.; Hoe, H.-S. Ibrutinib Suppresses LPS-Induced Neuroinflammatory Responses in BV2 Microglial Cells and Wild-Type Mice. J. Neuroinflammation 2018, 15, 271. [Google Scholar] [CrossRef]
- Xiao, Y.; Yang, C.; Si, N.; Chu, T.; Yu, J.; Yuan, X.; Chen, X.-T. Epigallocatechin-3-Gallate Inhibits LPS/AβO-Induced Neuroinflammation in BV2 Cells through Regulating the ROS/TXNIP/NLRP3 Pathway. J. Neuroimmune Pharmacol. 2024, 19, 31. [Google Scholar] [CrossRef]
- Sun, L.; Apweiler, M.; Tirkey, A.; Klett, D.; Normann, C.; Dietz, G.P.H.; Lehner, M.D.; Fiebich, B.L. Anti-Neuroinflammatory Effects of Ginkgo Biloba Extract EGb 761 in LPS-Activated BV2 Microglial Cells. Int. J. Mol. Sci. 2024, 25, 8108. [Google Scholar] [CrossRef]
- Fei, X.; Chen, L.; Gao, J.; Jiang, X.; Sun, W.; Cheng, X.; Zhao, T.; Zhao, M.; Zhu, L. P53 Lysine-Lactylated Modification Contributes to Lipopolysaccharide-Induced Proinflammatory Activation in BV2 Cell under Hypoxic Conditions. Neurochem. Int. 2024, 178, 105794. [Google Scholar] [CrossRef] [PubMed]
- Mairuae, N.; Buranrat, B.; Cheepsunthorn, P. Crude Extracts of Momordica cochinchinensis (Lour.) Spreng Exerts Antioxidant and Anti-Neuroinflammatory Properties in LPS-Stimulated BV2 Microglia. Trop. J. Pharm. Res. 2024, 23, 91–97. [Google Scholar] [CrossRef]
- Mairuae, N.; Buranrat, B.; Yannasithinon, S.; Cheepsunthorn, P. Oroxylum indicum Kurz (L) Leaf Extract Exerted Antioxidant and Anti-Inflammatory Effects on LPS-Stimulated BV2 Microglial Cells. Trop. J. Pharm. Res. 2024, 23, 1409–1415. [Google Scholar] [CrossRef]
- Foresti, R.; Bains, S.K.; Pitchumony, T.S.; de Castro Brás, L.E.; Drago, F.; Dubois-Randé, J.-L.; Bucolo, C.; Motterlini, R. Small Molecule Activators of the Nrf2-HO-1 Antioxidant Axis Modulate Heme Metabolism and Inflammation in BV2 Microglia Cells. Pharmacol. Res. 2013, 76, 132–148. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-I.; Cheng, C.-I.; Kang, Y.-F.; Chang, P.-C.; Lin, I.-P.; Kuo, Y.-H.; Jhou, A.-J.; Lin, M.-Y.; Chen, C.-Y.; Lee, C.-H. Hispidulin Inhibits Neuroinflammation in Lipopolysaccharide-Activated BV2 Microglia and Attenuates the Activation of Akt, NF-ΚB, and STAT3 Pathway. Neurotox Res. 2020, 38, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Lo, J.; Wu, H.-E.; Liu, C.-C.; Chang, K.-C.; Lee, P.-Y.; Liu, P.-L.; Huang, S.-P.; Wu, P.-C.; Lin, T.-C.; Lai, Y.-H. Nordalbergin Exerts Anti-Neuroinflammatory Effects by Attenuating MAPK Signaling Pathway, NLRP3 Inflammasome Activation and ROS Production in LPS-Stimulated BV2 Microglia. Int. J. Mol. Sci. 2023, 24, 7300. [Google Scholar] [CrossRef]
- Nguyen, P.L.; Bui, B.P.; Lee, H.; Cho, J. A Novel 1, 8-Naphthyridine-2-Carboxamide Derivative Attenuates Inflammatory Responses and Cell Migration in LPS-Treated BV2 Cells via the Suppression of ROS Generation and TLR4/Myd88/NF-ΚB Signaling Pathway. Int. J. Mol. Sci. 2021, 22, 2527. [Google Scholar] [CrossRef]
- Qin, J.; Ma, Z.; Chen, X.; Shu, S. Microglia Activation in Central Nervous System Disorders: A Review of Recent Mechanistic Investigations and Development Efforts. Front. Neurol. 2023, 14, 1103416. [Google Scholar] [CrossRef]
- Giovannini, M.G.; Scali, C.; Prosperi, C.; Bellucci, A.; Pepeu, G.; Casamenti, F. Experimental Brain Inflammation and Neurodegeneration as Model of Alzheimer’s Disease: Protective Effects of Selective COX-2 Inhibitors. Int. J. Immunopathol. Pharmacol. 2003, 16, 31–40. [Google Scholar]
- Skrzypczak-Wiercioch, A.; Sałat, K. Lipopolysaccharide-Induced Model of Neuroinflammation: Mechanisms of Action, Research Application and Future Directions for Its Use. Molecules 2022, 27, 5481. [Google Scholar] [CrossRef]
- Yang, L.; Zhou, R.; Tong, Y.; Chen, P.; Shen, Y.; Miao, S.; Liu, X. Neuroprotection by Dihydrotestosterone in LPS-Induced Neuroinflammation. Neurobiol. Dis. 2020, 140, 104814. [Google Scholar] [CrossRef] [PubMed]
- Simpson, D.S.A.; Oliver, P.L. ROS Generation in Microglia: Understanding Oxidative Stress and Inflammation in Neurodegenerative Disease. Antioxidants 2020, 9, 743. [Google Scholar] [CrossRef] [PubMed]
- Batista, C.R.A.; Gomes, G.F.; Candelario-Jalil, E.; Fiebich, B.L.; De Oliveira, A.C.P. Lipopolysaccharide-Induced Neuroinflammation as a Bridge to Understand Neurodegeneration. Int. J. Mol. Sci. 2019, 20, 2293. [Google Scholar] [CrossRef] [PubMed]
- Lull, M.E.; Block, M.L. Microglial Activation and Chronic Neurodegeneration. Neurotherapeutics 2010, 7, 354–365. [Google Scholar] [CrossRef]
- Wilks, A. Heme Oxygenase: Evolution, Structure, and Mechanism. Antioxid. Redox. Signal 2002, 4, 603–614. [Google Scholar] [CrossRef]
- Wu, Y.-H.; Hsieh, H.-L. Roles of Heme Oxygenase-1 in Neuroinflammation and Brain Disorders. Antioxidants 2022, 11, 923. [Google Scholar] [CrossRef]
- Guzmán-Beltrán, S.; Espada, S.; Orozco-Ibarra, M.; Pedraza-Chaverri, J.; Cuadrado, A. Nordihydroguaiaretic Acid Activates the Antioxidant Pathway Nrf2/HO-1 and Protects Cerebellar Granule Neurons against Oxidative Stress. Neurosci. Lett. 2008, 447, 167–171. [Google Scholar] [CrossRef]
- Hahn, D.; Shin, S.H.; Bae, J.-S. Natural Antioxidant and Anti-Inflammatory Compounds in Foodstuff or Medicinal Herbs Inducing Heme Oxygenase-1 Expression. Antioxidants 2020, 9, 1191. [Google Scholar] [CrossRef]
- Nwe, Y.Y.; Thwe, K.M.; Wai, T.P.; Moe, M.M. Phytochemical Investigation into Leaves of Elsholtzia Blanda (Benth.) Benth. and Its Antimicrobial Activity. J. Myanmar Acad. Arts Sci. 2020, 18, 11. [Google Scholar]
- Guo, Z.; Liu, Z.; Wang, X.; Liu, W.; Jiang, R.; Cheng, R.; She, G. Elsholtzia: Phytochemistry and Biological Activities. Chem. Cent. J. 2012, 6, 1–8. [Google Scholar] [CrossRef]
- Haiyun, L.; Yijia, L.; Honggang, L.; Honghai, W. Protective Effect of Total Flavones from Elsholtzia Blanda (TFEB) on Myocardial Ischemia Induced by Coronary Occlusion in Canines. J. Ethnopharmacol. 2004, 94, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Devi, N.K.; Dutta, S.; Das, R.; Devi, A.S.; Devi, N.M.; Devi, K.S. Nephroprotective Effect of Elsholtzia Blanda Benth. Paracetamol. Induc. Toxic. Albino Rats. Int. J. Recent. Sci. Res. 2018, 9, 28186–28189. [Google Scholar]
- Ling, H.; Lou, Y. Total Flavones from Elsholtzia Blanda Reduce Infarct Size during Acute Myocardial Ischemia by Inhibiting Myocardial Apoptosis in Rats. J. Ethnopharmacol. 2005, 101, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Zhou, C.X.; Lou, Y.J.; Duan, Z.H.; Zhao, Y. Chemical Constituents from Elsholtzia Blanda. Zhongguo Zhong Yao Za Zhi 2005, 30, 1589–1591. [Google Scholar]
- Jin-Shun, L.; Tong, S.; Zhen, G.U.O.; Xu-Wei, S.; Shang-Zhen, Z. The Chemical Constituents of Elsholtzia Blanda. J. Integr. Plant Biol. 2001, 43, 545. [Google Scholar]
- Dung, D.T.; Yen, P.H.; Trang, D.T.; Tai, B.H.; Kiem, P. Van Phenolic Constituents of Elsholtzia Blanda Benth. Nat. Prod. Commun. 2023, 18, 1934578X231192213. [Google Scholar]
- Ishwori, L.; Anupam, D.T.; Singh, P.K.; Dutta, C.M.; Deepa, N. Antibacterial Activity of Some Selected Plants Traditionally Used as Medicine in Manipur. Afr. J. Biotechnol. 2014, 13, 1491–1495. [Google Scholar] [CrossRef]
- Thi Dung, D.; Thi Trang, D.; Hai yen, P.; Huy Hoang, N.; Huu Tai, B.; Van Kiem, P. Elsholblanosides A− D, Four New Oleuropeic Acid Derivatives Isolated from Elsholtzia Blanda and Their Inhibition of NO Production in LPS-activated RAW264. 7 Cells. Chem Biodivers 2023, 20, e202300785. [Google Scholar] [CrossRef]
- Saturnino, C.; Sinicropi, M.S.; Parisi, O.I.; Iacopetta, D.; Popolo, A.; Marzocco, S.; Autore, G.; Caruso, A.; Cappello, A.R.; Longo, P. Acetylated Hyaluronic Acid: Enhanced Bioavailability and Biological Studies. Biomed. Res. Int. 2014, 2014, 921549. [Google Scholar] [CrossRef]
- Eruslanov, E.; Kusmartsev, S. Identification of ROS Using Oxidized DCFDA and Flow-Cytometry. In Advanced Protocols in Oxidative Stress II; Springer: Heidelberg, Germany, 2009; pp. 57–72. [Google Scholar]
- Baek, S.-H.; Park, T.; Kang, M.-G.; Park, D. Anti-Inflammatory Activity and ROS Regulation Effect of Sinapaldehyde in LPS-Stimulated RAW 264.7 Macrophages. Molecules 2020, 25, 4089. [Google Scholar] [CrossRef]
- Mishra, M.; Tiwari, S.; Gomes, A. V Protein Purification and Analysis: Next Generation Western Blotting Techniques. Expert. Rev. Proteom. 2017, 14, 1037–1053. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Sharad, S.; Minhas, G.; Sharma, D.R.; Bhatia, K.; Sharma, N.K. DNA, RNA Isolation, Primer Designing, Sequence Submission, and Phylogenetic Analysis. In Basic Biotechniques for Bioprocess and Bioentrepreneurship; Elsevier: Oxford, UK, 2023; pp. 197–206. [Google Scholar]
- Huang, W.; Zhang, M.; Qiu, Q.; Zhang, J.; Hua, C.; Chen, G.; Xie, H. Metabolomics of Human Umbilical Vein Endothelial Cell-Based Analysis of the Relationship between Hyperuricemia and Dyslipidemia. Nutr. Metab. Cardiovasc. Dis. 2024, 34, 1528–1537. [Google Scholar] [CrossRef] [PubMed]
- Balak, C.D.; Han, C.Z.; Glass, C.K. Deciphering Microglia Phenotypes in Health and Disease. Curr. Opin. Genet. Dev. 2024, 84, 102146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, Y.; Liu, T.; Mao, Y.; Peng, B. Novel Microglia-Based Therapeutic Approaches to Neurodegenerative Disorders. Neurosci. Bull. 2023, 39, 491–502. [Google Scholar] [CrossRef]
- Xu, Y.; Gao, W.; Sun, Y.; Wu, M. New Insight on Microglia Activation in Neurodegenerative Diseases and Therapeutics. Front. Neurosci. 2023, 17, 1308345. [Google Scholar] [CrossRef]
- Hines, D.J.; Choi, H.B.; Hines, R.M.; Phillips, A.G.; MacVicar, B.A. Prevention of LPS-Induced Microglia Activation, Cytokine Production and Sickness Behavior with TLR4 Receptor Interfering Peptides. PLoS ONE 2013, 8, e60388. [Google Scholar] [CrossRef]
- Plata-Salaman, C.R. Brain Cytokines and Disease. Acta Neuropsychiatr. 2002, 14, 262–278. [Google Scholar] [CrossRef]
- Salvemini, D.; Kim, S.F.; Mollace, V. Reciprocal Regulation of the Nitric Oxide and Cyclooxygenase Pathway in Pathophysiology: Relevance and Clinical Implications. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R473–R487. [Google Scholar] [CrossRef]
- Linares, D.; Taconis, M.; Mana, P.; Correcha, M.; Fordham, S.; Staykova, M.; Willenborg, D.O. Neuronal Nitric Oxide Synthase Plays a Key Role in CNS Demyelination. J. Neurosci. 2006, 26, 12672–12681. [Google Scholar] [CrossRef]
- Wei, S.; Yang, D.; Yang, J.; Zhang, X.; Zhang, J.; Fu, J.; Zhou, G.; Liu, H.; Lian, Z.; Han, H. Overexpression of Toll-like Receptor 4 Enhances LPS-Induced Inflammatory Response and Inhibits Salmonella Typhimurium Growth in Ovine Macrophages. Eur. J. Cell Biol. 2019, 98, 36–50. [Google Scholar] [CrossRef]
- Ahmed, T.; Zulfiqar, A.; Arguelles, S.; Rasekhian, M.; Nabavi, S.F.; Silva, A.S.; Nabavi, S.M. Map Kinase Signaling as Therapeutic Target for Neurodegeneration. Pharmacol. Res. 2020, 160, 105090. [Google Scholar] [CrossRef]
- Zarkovic, N. Roles and Functions of ROS and RNS in Cellular Physiology and Pathology. Cells 2020, 9, 767. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.-H.; Chen, D.-Q.; Jin, Y.-H.; Han, Y.-H.; Sun, H.-N.; Kwon, T. Hispidin Inhibits LPS-Induced Nitric Oxide Production in BV-2 Microglial Cells via pendent MAPK Signaling. Exp. Ther. Med. 2021, 22, 970. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.A.; Selvaraj, B.; Vu, H.T.; Nguyen, Q.N.S.; Oanh, L.H.; Lee, H.; Tran, Q.L.; Oanh, V.T.; Jung, S.H.; Thuong, P.T. The Ethanol Extract of Polyscias Scutellaria (EEPS) Shows Anti-Inflammatory Signaling Pathway Against LPS-Induced Inflammation in RAW 264.7 Macrophages. Nat. Prod. Commun. 2025, 20, 1934578X251324006. [Google Scholar] [CrossRef]
- Meimei, C.; Fei, Z.; Wen, X.; Huangwei, L.; Zhenqiang, H.; Rongjun, Y.; Qiang, Z.; Qiuyang, L.; Xiaozhen, L.; Yuan, Y. Taxus Chinensis (Pilg.) Rehder Fruit Attenuates Aging Behaviors and Neuroinflammation by Inhibiting Microglia Activation via TLR4/NF-ΚB/NLRP3 Pathway. J. Ethnopharmacol. 2025, 337, 118943. [Google Scholar] [CrossRef]
- Minasyan, A.; Pires, V.; Gondcaille, C.; Ginovyan, M.; Mróz, M.; Savary, S.; Cherkaoui-Malki, M.; Kusznierewicz, B.; Bartoszek, A.; Andreoletti, P. Ribes Nigrum Leaf Extract Downregulates Pro-Inflammatory Gene Expression and Regulates Redox Balance in Microglial Cells. BMC Complement. Med. Ther. 2025, 25, 49. [Google Scholar] [CrossRef]
- Lim, J.S.; Li, X.; Lee, D.Y.; Yao, L.; Yoo, G.; Kim, Y.; Eum, S.M.; Cho, Y.-C.; Yoon, S.; Park, S.-J. Antioxidant and Anti-Inflammatory Activities of Methanol Extract of Senna Septemtrionalis (Viv.) HS Irwin & Barneby Through Nrf2/HO-1-Mediated Inhibition of NF-ΚB Signaling in LPS-Stimulated Mouse Microglial Cells. Int. J. Mol. Sci. 2025, 26, 1932. [Google Scholar]
- Gill, A.J.; Kovacsics, C.E.; Cross, S.A.; Vance, P.J.; Kolson, L.L.; Jordan-Sciutto, K.L.; Gelman, B.B.; Kolson, D.L. Heme Oxygenase-1 Deficiency Accompanies Neuropathogenesis of HIV-Associated Neurocognitive Disorders. J. Clin. Investig. 2014, 124, 4459–4472. [Google Scholar] [CrossRef]
- Feng, C.-W.; Chen, N.-F.; Wen, Z.-H.; Yang, W.-Y.; Kuo, H.-M.; Sung, P.-J.; Su, J.-H.; Cheng, S.-Y.; Chen, W.-F. In Vitro and in Vivo Neuroprotective Effects of Stellettin B through Anti-Apoptosis and the Nrf2/HO-1 Pathway. Mar. Drugs 2019, 17, 315. [Google Scholar] [CrossRef]
- Li, Y.C.; Hao, J.C.; Shang, B.; Zhao, C.; Wang, L.J.; Yang, K.L.; He, X.Z.; Tian, Q.Q.; Wang, Z.L.; Jing, H.L. Neuroprotective Effects of Aucubin on Hydrogen Peroxide-Induced Toxicity in Human Neuroblastoma SH-SY5Y Cells via the Nrf2/HO-1 Pathway. Phytomedicine 2021, 87, 153577. [Google Scholar]
- Yang, Y.; Yu, L.; Zhu, T.; Xu, S.; He, J.; Mao, N.; Liu, Z.; Wang, D. Neuroprotective Effects of Lycium Barbarum Polysaccharide on Light-Induced Oxidative Stress and Mitochondrial Damage via the Nrf2/HO-1 Pathway in Mouse Hippocampal Neurons. Int. J. Biol. Macromol. 2023, 251, 126315. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Lu, Y.; Yang, F.; Li, S.; He, X.; Gao, Y.; Zhang, G.; Ren, E.; Wang, Y.; Kang, X. Rosmarinic Acid Exerts a Neuroprotective Effect on Spinal Cord Injury by Suppressing Oxidative Stress and Inflammation via Modulating the Nrf2/HO-1 and TLR4/NF-ΚB Pathways. Toxicol. Appl. Pharmacol. 2020, 397, 115014. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W.; Choi, A.M.K. Heme Oxygenase-1: Molecular Mechanisms of Gene Expression in Oxygen-Related Stress. Antioxid. Redox. Signal 2002, 4, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Biswas, C.; Shah, N.; Muthu, M.; La, P.; Fernando, A.P.; Sengupta, S.; Yang, G.; Dennery, P.A. Nuclear Heme Oxygenase-1 (HO-1) Modulates Subcellular Distribution and Activation of Nrf2, Impacting Metabolic and Anti-Oxidant Defenses. J. Biol. Chem. 2014, 289, 26882–26894. [Google Scholar] [CrossRef]
- Balogun, E.; Hoque, M.; Gong, P.; Killeen, E.; Green, C.J.; Foresti, R.; Alam, J.; Motterlini, R. Curcumin Activates the Haem Oxygenase-1 Gene via Regulation of Nrf2 and the Antioxidant-Responsive Element. Biochem. J. 2003, 371, 887–895. [Google Scholar] [CrossRef]
- Kietzmann, T.; Samoylenko, A.; Immenschuh, S. Transcriptional Regulation of Heme Oxygenase-1 Gene Expression by MAP Kinases of the JNK and P38 Pathways in Primary Cultures of Rat Hepatocytes. J. Biol. Chem. 2003, 278, 17927–17936. [Google Scholar] [CrossRef]
- Martin, D.; Rojo, A.I.; Salinas, M.; Diaz, R.; Gallardo, G.; Alam, J.; De Galarreta, C.M.R.; Cuadrado, A. Regulation of Heme Oxygenase-1 Expression through the Phosphatidylinositol 3-Kinase/Akt Pathway and the Nrf2 Transcription Factor in Response to the Antioxidant Phytochemical Carnosol. J. Biol. Chem. 2004, 279, 8919–8929. [Google Scholar] [CrossRef]
- Kaur, V.; Kumar, M.; Kumar, A.; Kaur, K.; Dhillon, V.S.; Kaur, S. Pharmacotherapeutic Potential of Phytochemicals: Implications in Cancer Chemoprevention and Future Perspectives. Biomed. Pharmacother. 2018, 97, 564–586. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Accession No. |
|---|---|---|
| mouse iNOS | Forward 5′-ATGGACCAGTATAAGGCAAGC-3′ Reverse 5′-GCTCTGGATGAGCCTATATTG-3′ | BC062378 |
| mouse TNF-α | Forward 5′-GGTGCCTATGTCTCAGCCTCTT-3′ Reverse 5′-GCCATAGAACTGATGAGAGGGAG-3′ | BC117057 |
| mouse IL-6 | Forward 5′-TACCACTTCACAAGTCGGAGGC-3′ Reverse 5′-CTGCAAGTGCATCATCGTTGTTC-3′ | BC132458 |
| mouse IL-1β | Forward 5′-TGGACCTTCCAGGATGAGGACA -3′ Reverse 5′- GTTCATCTCGGAGCCTGTAGTG -3′ | BC011437 |
| mouse IL-8 | Forward 5′-CTCTATTCTGCCAGATGCTGTCC-3′ Reverse 5′-ACAAGGCTCAGCAGAGTCACCA-3′ | BC051677 |
| mouse IL-18 | Forward 5′-GACAGCCTGTGTTCGAGGATATG-3′ Reverse 5′-TGTTCTTACAGGAGAGGGTAGAC-3′ | NM_008360 |
| mouse IL-10 | Forward 5′-CGGGAAGACAATAACTGCACCC-3′ Reverse 5′-CGGTTAGCAGTATGTTGTCCAGC-3′ | NM_010548 |
| mouse HO-1 | Forward 5′-TTACCTTCCCGAACATCGAC-3′ Reverse 5′-GCATAAATTCCCACTGCCAC-3′ | BC010757 |
| mouse GAPDH | Forward 5′-GCGAGACCCCACTAACATCA-3′ Reverse 5′-GAGTTGGGATAGGGCCTCTCTT-3′ | GU214026 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, Y.D.; Shrestha, K.K.; Baek, S.-H. In Vitro Antioxidant and Anti-Neuroinflammatory Effects of Elsholtzia blanda (Benth.) Benth. Life 2025, 15, 983. https://doi.org/10.3390/life15060983
Yoon YD, Shrestha KK, Baek S-H. In Vitro Antioxidant and Anti-Neuroinflammatory Effects of Elsholtzia blanda (Benth.) Benth. Life. 2025; 15(6):983. https://doi.org/10.3390/life15060983
Chicago/Turabian StyleYoon, Yeo Dae, Krishna K. Shrestha, and Seung-Hwa Baek. 2025. "In Vitro Antioxidant and Anti-Neuroinflammatory Effects of Elsholtzia blanda (Benth.) Benth." Life 15, no. 6: 983. https://doi.org/10.3390/life15060983
APA StyleYoon, Y. D., Shrestha, K. K., & Baek, S.-H. (2025). In Vitro Antioxidant and Anti-Neuroinflammatory Effects of Elsholtzia blanda (Benth.) Benth. Life, 15(6), 983. https://doi.org/10.3390/life15060983