ATPe Dynamics in Protozoan Parasites. Adapt or Perish


 and
and 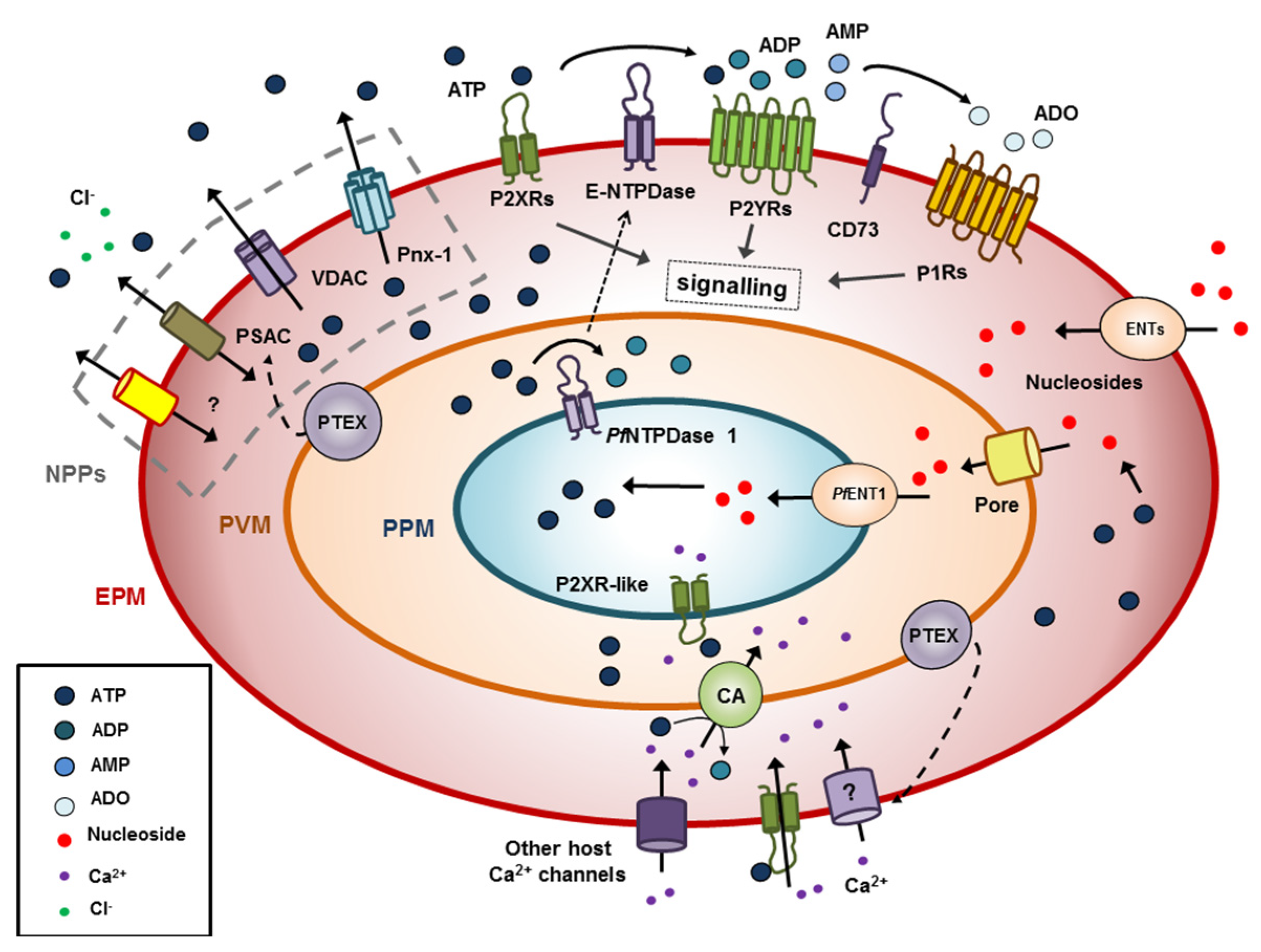{kind=link}
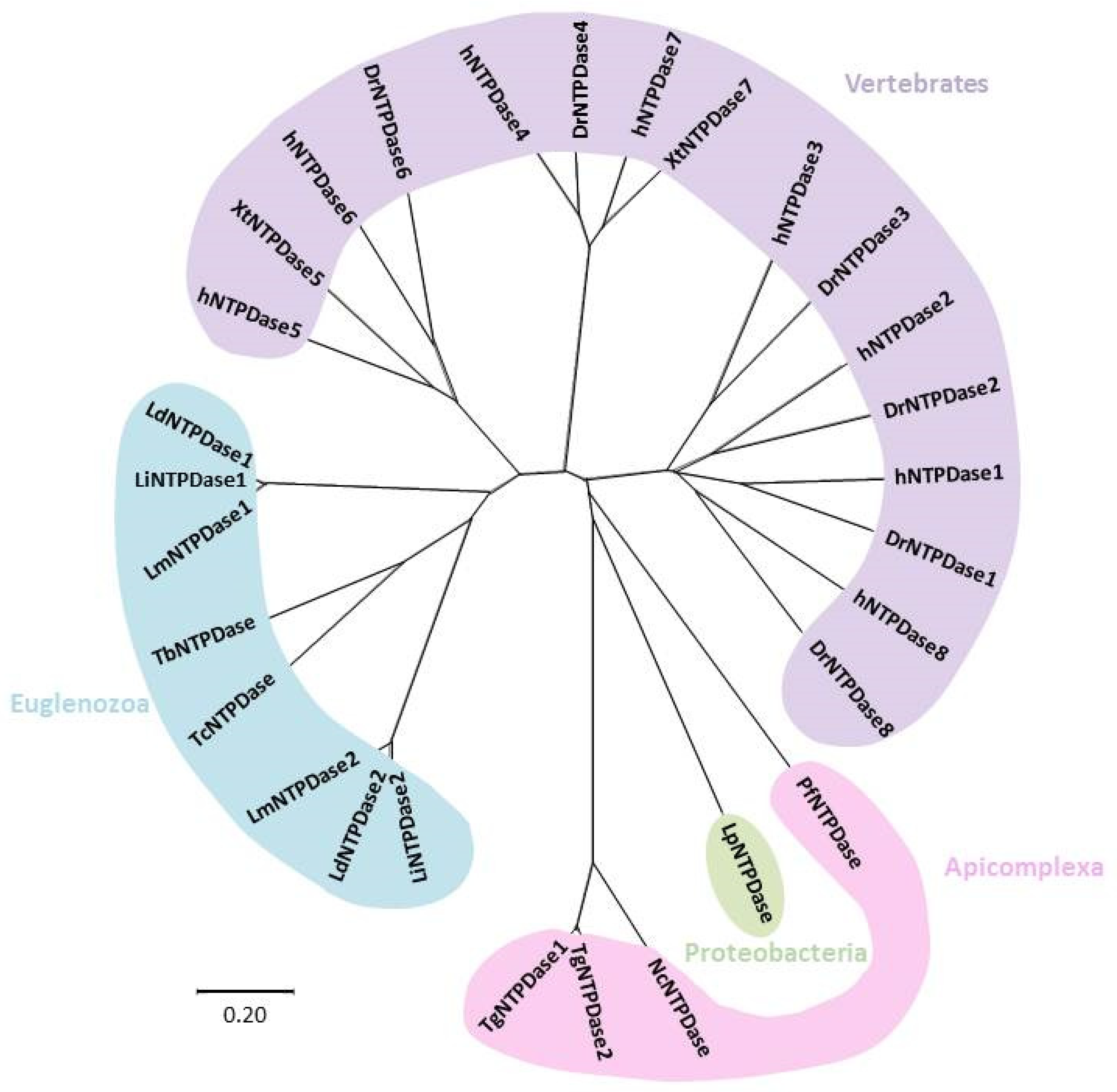{kind=link}
Abstract
1. Introduction
2. Purinergic Signaling
2.1. P2X Receptors. Evolution and Early Appearance in Protozoa
2.1.1. A Brief Tale of Three Protozoans
Monosiga brevicollis
Dictyostelium discoideum
Plasmodium falciparum, a Parasitic Protozoon
2.2. P receptors of the Host and Parasite Infection
3. Transport of ATP
4. Ecto-Nucleotidases
4.1. Toxoplasma gondii
4.2. Trypanosoma cruzi
4.3. Leishmania
4.4. Plasmodium falciparum
5. Nucleoside Transport
Purine Salvage of Parasitic Protozoa
6. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ramdani, G.; Langsley, G. ATP, an extracellular signaling molecule in red blood cells: A messenger for malaria? Biomed. J. 2014, 37, 284–292. [Google Scholar] [PubMed]
- Burnstock, G.; Verkhratsky, A. Purinergic Signalling and the Nervous System; Springer: Heidelberg, Germany, 2012; ISBN 9783642288630. [Google Scholar]
- Burnstock, G. Purinergic signalling: From discovery to current developments. Exp. Physiol. 2014, 99, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Banerjee, J.; Leung, C.T.; Peterson-Yantorno, K.; Stamer, W.D.; Civan, M.M. Mechanisms of ATP release, the enabling step in purinergic dynamics. Cell. Physiol. Biochem. 2011, 28, 1135–1144. [Google Scholar] [CrossRef]
- Taruno, A. ATP release channels. Int. J. Mol. Sci. 2018, 19, 808. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, H.; Zebisch, M.; Sträter, N. Cellular function and molecular structure of ecto-nucleotidases. Purinergic Signal. 2012, 8, 437–502. [Google Scholar] [CrossRef] [PubMed]
- Yegutkin, G.G. Enzymes involved in metabolism of extracellular nucleotides and nucleosides: Functional implications and measurement of activities. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 473–497. [Google Scholar] [CrossRef] [PubMed]
- De Koning, H.P.; Bridges, D.J.; Burchmore, R.J.S. Purine and pyrimidine transport in pathogenic protozoa: From biology to therapy. FEMS Microbiol. Rev. 2005, 29, 987–1020. [Google Scholar] [CrossRef]
- Coutinho-Silva, R.; Corrêa, G.; Sater, A.A.; Ojcius, D.M. The P2X7 receptor and intracellular pathogens: A continuing struggle. Purinergic Signal. 2009, 5, 197–204. [Google Scholar] [CrossRef]
- Coutinho-Silva, R.; Ojcius, D.M. Role of extracellular nucleotides in the immune response against intracellular bacteria and protozoan parasites. Microbes Infect. 2012, 14, 1271–1277. [Google Scholar] [CrossRef]
- Lebrun, M.; Blanchard, N. Editorial overview: Host–microbe interactions: Parasites. Curr. Opin. Microbiol. 2017, 40, viii–xi. [Google Scholar] [CrossRef]
- Savio, L.E.B.; de Mello, P.A.; da Silva, C.G.; Coutinho-Silva, R. The P2X7 receptor in inflammatory diseases: Angel or demon? Front. Pharmacol. 2018, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Akkaya, C.; Shumilina, E.; Bobballa, D.; Brand, V.B.; Mahmud, H.; Lang, F.; Huber, S.M. The Plasmodium falciparum-induced anion channel of human erythrocytes is an ATP-release pathway. Pflugers Arch. Eur. J. Physiol. 2009, 457, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, C.L.; Schachter, J.; de Sá Pinheiro, A.A.; de Souza Silva, L.; Verstraeten, S.V.; Persechini, P.M.; Schwarzbaum, P.J. Regulation of extracellular ATP in human erythrocytes infected with Plasmodium falciparum. PLoS ONE 2014, 9, e96216. [Google Scholar] [CrossRef] [PubMed]
- Levano-Garcia, J.; Dluzewski, A.R.; Markus, R.P.; Garcia, C.R.S. Purinergic signalling is involved in the malaria parasite Plasmodium falciparum invasion to red blood cells. Purinergic Signal. 2010, 6, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Tanneur, V.; Duranton, C.; Brand, V.B.; Sandu, C.D.; Akkaya, C.; Kasinathan, R.S.; Gachet, C.; Sluyter, R.; Barden, J.A.; Wiley, J.S.; et al. Purinoceptors are involved in the induction of an osmolyte permeability in malaria-infected and oxidized human erythrocytes. FASEB J. 2006, 20, 133–135. [Google Scholar] [CrossRef] [PubMed]
- Fleck, S.L.; Birdsall, B.; Babon, J.; Dluzewski, A.R.; Martin, S.R.; Morgan, W.D.; Angov, E.; Kettleborough, C.A.; Feeney, J.; Blackman, M.J.; et al. Suramin and suramin analogues inhibit merozoite surface protein-1 secondary processing and erythrocyte invasion by the malaria parasite Plasmodium falciparum. J. Biol. Chem. 2003, 278, 47670–47677. [Google Scholar] [CrossRef]
- de Souza, M.C.; De Assis, E.A.; Saar Gomes, R.; De Almeida Marques-Da-Silva, E.; Melo, M.N.; Lopes Rangel Fietto, J.; Crocco Alfonso, L.C.; Afonso, C. The influence of ecto-nucleotidases on Leishmania amazonensis infection and immune response in C57B/6 mice. Acta Trop. 2010, 115, 262–269. [Google Scholar] [CrossRef]
- Leite, P.M.; Gomes, R.S.; Figueiredo, A.B.; Serafim, T.D.; Tafuri, W.L.; de Souza, C.C.; Moura, S.A.L.; Fietto, J.L.R.; Melo, M.N.; Ribeiro-Dias, F.; et al. Ecto-nucleotidase activities of promastigotes from Leishmania (Viannia) braziliensis relates to parasite infectivity and disease clinical outcome. PLoS Negl. Trop. Dis. 2012, 6, e1850. [Google Scholar] [CrossRef]
- Khakh, B.S.; Alan North, R. P2X receptors as cell-surface ATP sensors in health and disease. Nature 2006, 442, 527–532. [Google Scholar] [CrossRef]
- Burnstock, G. Blood cells: An historical account of the roles of purinergic signalling. Purinergic Signal. 2015, 11, 411–434. [Google Scholar] [CrossRef]
- Coddou, C.; Yan, Z.; Obsil, T.; Huidobro-Toro, J.P.; Stojilkovic, S.S. Activation and regulation of purinergic P2X receptor channels. Pharmacol. Rev. 2011, 63, 641–683. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G.; Ralevic, V.; Burnstock, G. Receptor for purines and pyrimidines. Pharmacol. Rev. 1998, 50, 413–492. [Google Scholar]
- North, R.A. Molecular physiology of P2X receptors. Physiol. Rev. 2002, 82, 1013–1067. [Google Scholar] [CrossRef]
- Espelt, M.V.; de Tezanos Pinto, F.; Alvarez, C.L.; Alberti, G.S.; Incicco, J.; Leal Denis, M.F.; Davio, C.; Schwarzbaum, P.J. On the role of ATP release, ectoATPase activity, and extracellular ADP in the regulatory volume decrease of Huh-7 human hepatoma cells. Am. J. Physiol. Cell Physiol. 2013, 304, C1013–C1026. [Google Scholar] [CrossRef]
- Pafundo, D.E.; Alvarez, C.L.; Krumschnabel, G.; Schwarzbaum, P.J. A volume regulatory response can be triggered by nucleosides in human erythrocytes, a perfect osmometer no longer. J. Biol. Chem. 2010, 285, 6134–6144. [Google Scholar] [CrossRef] [PubMed]
- Fountain, S.J. Primitive ATP-activated P2X receptors: Discovery, function and pharmacology. Front. Cell. Neurosci. 2013, 7, 2007–2013. [Google Scholar] [CrossRef]
- Kaczmarek-Hájek, K.; Lörinczi, É.; Hausmann, R.; Nicke, A. Molecular and functional properties of P2X receptors-recent progress and persisting challenges. Purinergic Signal. 2012, 8, 375–417. [Google Scholar] [CrossRef]
- Newbolt, A.; Stoop, R.; Virginio, C.; Surprenant, A.; North, R.A.; Buell, G.; Rassendren, F. Membrane topology of an ATP-gated ion channel (P2X receptor). J. Biol. Chem. 1998, 273, 15177–15182. [Google Scholar] [CrossRef] [PubMed]
- Abbracchio, M.P.; Burnstock, G.; Verkhratsky, A.; Zimmermann, H. Purinergic signalling in the nervous system: An overview. Trends Neurosci. 2009, 32, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, Y. Insights into the channel gating of P2X receptors from structures, dynamics and small molecules. Acta Pharmacol. Sin. 2016, 37, 44–55. [Google Scholar] [CrossRef]
- Nicke, A.; Bäumert, H.G.; Rettinger, J.; Eichele, A.; Lambrecht, G.; Mutschler, E.; Schmalzing, G. P2X1 and P2X3 receptors form stable trimers: A novel structural motif of ligand-gated ion channels. EMBO J. 1998, 17, 3016–3028. [Google Scholar] [CrossRef] [PubMed]
- Chaumont, S.; Jiang, L.H.; Penna, A.; North, R.A.; Rassendren, F. Identification of a trafficking motif involved in the stabilization and polarization of P2X receptors. J. Biol. Chem. 2004, 279, 29628–29638. [Google Scholar] [CrossRef] [PubMed]
- Volonté, C.; D’Ambrosi, N. Membrane compartments and purinergic signalling: The purinome, a complex interplay among ligands, degrading enzymes, receptors and transporters. FEBS J. 2009, 276, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Sachs, F. Single channel properties of P2X 2 purinoceptors. J. Gen. Physiol. 1999, 113, 695–720. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.-H.; Spelta, V.; Bo, X.; Surprenant, A.; North, R.A. Status survey of the forest leopard (Panthera pardus Linnaeus, 1758) in Nepal. Subunit Arrangement P2X Receptors. 2003, 23, 8903–8910. [Google Scholar]
- Alvarez, C.L.; Corradi, G.; Lauri, N.; Marginedas-Freixa, I.; Leal Denis, M.F.; Enrique, N.; Mate, S.M.; Milesi, V.; Ostuni, M.A.; Herlax, V.; et al. Dynamic regulation of extracellular ATP in Escherichia coli. Biochem. J. 2017, 474, 1395–1416. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Burnstock, G. Biology of purinergic signalling: Its ancient evolutionary roots, its omnipresence and its multiple functional significance. BioEssays 2014, 36, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Fountain, S.J.; Cao, L.; Young, M.T.; North, R.A. Permeation properties of a P2X receptor in the green algae Ostreococcus tauri. J. Biol. Chem. 2008, 283, 15122–15126. [Google Scholar] [CrossRef] [PubMed]
- Hefetz, A.; Keeling, C.I.; Winston, M.L.; Slessor, K.N.; Nielsen, J.; Lanfear, R.; Liebig, J.; Millar, J.G.; Hanks, L.M.; Suarez, A.V.; et al. Identification of a plant receptor for extracellular ATP. Science 2014, 343, 290–295. [Google Scholar]
- Clark, G.; Roux, S.J. Apyrases, extracellular ATP and the regulation of growth. Curr. Opin. Plant Biol. 2011, 14, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Ludlow, M.J.; Ludlow, M.J. Purinergic signalling in Dictyostelium discoideum. Ph.D. Thesis, University of Leicester, Leicester, UK, 2008. [Google Scholar]
- Fountain, S.J.; Parkinson, K.; Young, M.T.; Cao, L.; Thompson, C.R.L.; North, R.A. An intracellular P2X receptor required for osmoregulation in Dictyostelium discoideum. Nature 2007, 448, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, K.; Ohno, Y. Neighboring glycine residues are essential for P2X2 receptor/channel function. Eur. J. Pharmacol. 1999, 370, 247–248. [Google Scholar] [CrossRef]
- Agboh, K.C.; Webb, T.E.; Evans, R.J.; Ennion, S.J. Functional characterization of a P2X receptor from Schistosoma mansoni. J. Biol. Chem. 2004, 279, 41650–41657. [Google Scholar] [CrossRef] [PubMed]
- Hayasi, M.; Takahashi, M. Ciliary adenosinetriphosphatase from a slow swimming mutant of Paramecium caudatum. J. Biol. Chem. Biol. Chem. 1979, 254, 11561–11565. [Google Scholar]
- Kim, M.Y.; Kuruvilla, H.G.; Raghu, S.; Hennessey, T.M. ATP reception and chemosensory adaptation in Tetrahymena thermophila. J. Exp. Biol. 1999, 202, 407–416. [Google Scholar] [PubMed]
- Inverso, J.A.; Song, Y.; Santos-Buch, C.A. Plasma membrane ATP receptors in Trypanosoma cruzi trypomastigotes. Receptor 1995, 5, 197–206. [Google Scholar] [PubMed]
- Pothier, F.; Forget, J.; Sullivan, R.; Couillard, P. ATP and the contractile vacuole in Amoeba proteus: Mechanism of action of exogenous ATP and related nucleotides. J. Exp. Zool. 1987, 243, 379–387. [Google Scholar] [CrossRef]
- Uyeda, T.Q.P.; Furuya, M. ATP-induced relative movement between microfilaments and microtubules in myxomycete flagellates. Protoplasma 1987, 140, 190–192. [Google Scholar] [CrossRef]
- Yue, J.; Sun, G.; Hu, X.; Huang, J. The scale and evolutionary significance of horizontal gene transfer in the choanoflagellate Monosiga brevicollis. BMC Genom. 2013, 14, 729. [Google Scholar] [CrossRef]
- King, N.; Westbrook, M.J.; Young, S.L.; Kuo, A.; Abedin, M.; Chapman, J.; Fairclough, S.; Hellsten, U.; Isogai, Y.; Letunic, I.; et al. The genome of the choanoflagellate Monosiga brevicollis and the origin of metazoans. Nature 2008, 451, 783–788. [Google Scholar] [CrossRef]
- Eichinger, I.; Pachebat, J.A.; Glöckner, G.; Rajandream, M.A.; Sucgang, R.; Berriman, M.; Song, J.; Olsen, R.; Szafranski, K.; Xu, Q.; et al. The genome of the social amoeba Dictyostelium discoideum. Nature 2005, 435, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Leal Denis, M.F.; Incicco, J.J.; Espelt, M.V.; Verstraeten, S.V.; Pignataro, O.P.; Lazarowski, E.R.; Schwarzbaum, P.J. Kinetics of extracellular ATP in mastoparan 7-activated human erythrocytes. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 4692–4707. [Google Scholar] [CrossRef] [PubMed]
- Leal Denis, M.F.; Alvarez, H.A.; Lauri, N.; Alvarez, C.L.; Chara, O.; Schwarzbaum, P.J. Dynamic regulation of cell volume and extracellular ATP of human erythrocytes. PLoS ONE 2016, 11, e0158305. [Google Scholar] [CrossRef] [PubMed]
- Surprenant, A.; North, R.A. Signaling at Purinergic P2X Receptors. Annu. Rev. Physiol. 2009, 71, 333–359. [Google Scholar] [CrossRef] [PubMed]
- Sivaramakrishnan, V.; Fountain, S.J. Evidence for extracellular ATP as a stress signal in a single-celled organism. Eukaryot. Cell 2015, 14, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Ghazanfari, N.; Mueller, S.N.; Heath, W.R. Cerebral malaria in mouse and man. Front. Immunol. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Cowman, A.F.; Tonkin, C.J.; Tham, W.H.; Duraisingh, M.T. The molecular basis of erythrocyte invasion by malaria parasites. Cell Host Microbe 2017, 22, 232–245. [Google Scholar] [CrossRef]
- Ashley, E.A.; Phyo, P.; Woodrow, C.J. Seminar Malaria. Lancet 2018, 6736, 1–14. [Google Scholar]
- Gardner, M.J.; Hall, N.; Fung, E.; White, O.; Berriman, M.; Hyman, R.W.; Carlton, J.M.; Pain, A.; Nelson, K.E.; Bowman, S.; et al. Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 2002, 419, 498–511. [Google Scholar] [CrossRef]
- Frech, C.; Chen, N. Genome comparison of human and non-human malaria parasites reveals species subset-specific genes potentially linked to human disease. PLoS Comput. Biol. 2011, 7, e1002320. [Google Scholar] [CrossRef]
- Budu, A.; Garcia, C.R.S. Generation of second messengers in Plasmodium. Microbes Infect. 2012, 14, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Maier, A.G.; Cooke, B.M.; Cowman, A.F.; Tilley, L. Malaria parasite proteins that remodel the host erythrocyte. Nat. Rev. Microbiol. 2009, 7, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Tilley, L.; Sougrat, R.; Lithgow, T.; Hanssen, E. The twists and turns of Maurer’s cleft trafficking in P. falciparum-infected erythrocytes. Traffic 2008, 9, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.A. Why do malaria parasites increase host erythrocyte permeability? Trends Parasitol. 2014, 30, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Balabaskaran-nina, P.; Nguitragool, W.; Saggu, G.S.; Schureck, M.A.; Desai, A. CLAG3 Self-associates in malaria parasites and quantitatively determines nutrient uptake channels at the host membrane. Am. Soc. Microbiol. 2018, 9, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Chalapareddy, S.; Desai, S.A. Malaria parasite proteins involved in nutrient channels at the host erythrocyte membrane: Advances and questions for future research. Int. J. Curr. Multidiscip. Stud. 2017, 3, 619–623. [Google Scholar] [PubMed]
- Marginedas-Freixa, I.; Hattab, C.; Bouyer, G.; Halle, F.; Chene, A.; Lefevre, S.D.; Cambot, M.; Cueff, A.; Schmitt, M.; Gamain, B.; et al. TSPO ligands stimulate ZnPPIX transport and ROS accumulation leading to the inhibition of P. falciparum growth in human blood. Sci. Rep. 2016, 6, 33516. [Google Scholar] [CrossRef]
- Brochet, M.; Billker, O. Calcium signalling in malaria parasites. Mol. Microbiol. 2016, 100, 397–408. [Google Scholar] [CrossRef]
- Zipprer, E.M.; Neggers, M.; Kushwaha, A.; Rayavara, K.; Desai, S.A. A kinetic fluorescence assay reveals unusual features of Ca++ uptake in Plasmodium falciparum-infected erythrocytes. Lipids Health Dis. 2014, 13, 184. [Google Scholar] [CrossRef]
- Wasserman, M.; Alarcón, C.; Mendoza, P.M. Effects of Ca++ depletion on the asexual cell cycle of Plasmodium falciparum. Am. J. Trop. Med. Hyg. 1982, 31, 711–717. [Google Scholar] [CrossRef]
- Cruz, L.; Juliano, M.; Budu, A.; Juliano, L.; Holder, A.A.; Blackman, M.J.; Garcia, C.R. Extracellular ATP triggers proteolysis and cytosolic Ca2+ rise in Plasmodium berghei and Plasmodium yoelii malaria parasites. Malar. J. 2012, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Kanaani, J.; Ginsburg, H. Compartment analysis of ATP in malaria-infected erythrocytes. Biochem. Int. 1988, 17, 451–459. [Google Scholar] [PubMed]
- Gorman, M.W.; Feigl, E.O.; Buffington, C.W. Human plasma ATP concentration. Clin. Chem. 2007, 53, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Gazarini, M.L.; Thomas, A.P.; Pozzan, T.; Garcia, C.R.S. Calcium signaling in a low calcium environment: How the intracellular malaria parasite solves the problem. J. Cell Biol. 2003, 161, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Adovelande, J.; Bastide, B.; Délèze, J.; Schrével, J. Cytosolic free calcium in Plasmodium falciparum-infected erythrocytes and the effect of verapamil: A cytofluorimetric study. Exp. Parasitol. 1993, 76, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.R.; Dluzewski, A.R.; Catalani, L.H.; Burting, R.; Hoyland, J.; Mason, W.T. Calcium homeostasis in intraerythrocytic malaria parasites. Eur. J. Cell Biol. 1996, 71, 409–413. [Google Scholar] [PubMed]
- Kushwaha, A.K.; Apolis, L.; Ito, D.; Desai, S.A. Increased Ca++ uptake by erythrocytes infected with malaria parasites: Evidence for exported proteins and novel inhibitors. Cell. Microbiol. 2018, 20, e12853. [Google Scholar] [CrossRef]
- Schlesinger, P.H.; Krogstad, D.J.; Desai, S.A.; McCleskey, E.W. A novel pathway for Ca++ entry into Plasmodium falciparum-infected blood cells. Am. J. Trop. Med. Hyg. 1996, 54, 464–470. [Google Scholar]
- Miller, C.M.; Boulter, N.R.; Fuller, S.J.; Zakrzewski, A.M.; Lees, M.P.; Saunders, B.M.; Wiley, J.S.; Smith, N.C. The role of the P2X7 receptor in infectious diseases. PLoS Pathog. 2011, 7, e1002212. [Google Scholar] [CrossRef]
- Sibley, L.D. Toxoplasma gondii: Perfecting an intracellular life style. Traffic 2003, 4, 581–586. [Google Scholar] [CrossRef]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Krug, U.; Zebisch, M.; Krauss, M.; Sträter, N. Structural insight into activation mechanism of Toxoplasma gondii nucleoside triphosphate diphosphohydrolases by disulfide reduction. J. Biol. Chem. 2012, 287, 3051–3066. [Google Scholar] [CrossRef] [PubMed]
- Mercier, C.; Dubremetz, J.-F.; Rauscher, B.; Lecordier, L.; Sibley, L.D.; Cesbron-Delauw, M.-F. Biogenesis of nanotubular network in Toxoplasma parasitophorous vacuole induced by parasite proteins. Mol. Biol. Cell 2002, 13, 2397–2409. [Google Scholar] [CrossRef] [PubMed]
- Laliberté, J.; Carruthers, V.B. Host cell manipulation by the human pathogen Toxoplasma gondii. Cell Mol Life Sci. 2008, 65, 1900–1915. [Google Scholar]
- Corrêa, G.; Marques da Silva, C.; de Abreu Moreira-Souza, A.C.; Vommaro, R.C.; Coutinho-Silva, R. Activation of the P2X7 receptor triggers the elimination of Toxoplasma gondii tachyzoites from infected macrophages. Microbes Infect. 2010, 12, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Lees, M.P.; Fuller, S.J.; Mcleod, R.; Boulter, N.R.; Catherine, M.; Zakrzewski, A.M.; Mui, E.J.; Witola, W.H.; Coyne, J.J.; Hargrave, C.; et al. P2X7 receptor-mediated killing of an intracellular parasite, Toxoplasma gondii, by human and murine macrophages1. J Immunol. 2010, 184, 7040–7046. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Souza, A.C.A.; Almeida-da-Silva, C.L.C.; Rangel, T.P.; Rocha, G.D.C.; Bellio, M.; Zamboni, D.S.; Vommaro, R.C.; Coutinho-Silva, R. The P2X7 receptor mediates Toxoplasma gondii control in macrophages through canonical NLRP3 inflammasome activation and reactive oxygen species production. Front. Immunol. 2017, 8, 1257. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, S.E.; Peixoto-Rangel, A.L.; Hargrave, A.C.; Roubaix, L.A.D.; Mui, E.J.; Boulter, N.R.; Miller, E.N.; Fuller, S.J.; Wiley, J.S.; Castellucci, L.; et al. Evidence for associations between the purinergic receptor P2X7 (P2RX7) and toxoplasmosis. Genes Immun. 2010, 11, 374–383. [Google Scholar] [CrossRef]
- Melo, M.B.; Jensen, K.D.C.; Saeij, J.P.J. Toxoplasma gondii effectors are master regulators of the inflammatory response. Trends Parasitol. 2011, 27, 487–495. [Google Scholar] [CrossRef]
- Sansom, F.M. The role of the NTPDase enzyme family in parasites: What do we know, and where to from here? Parasitology 2012, 139, 963–980. [Google Scholar] [CrossRef]
- Asai, T.; Miura, S.; Sibley, L.D.; Okabayashi, H.; Takeuchi, T. Biochemical and molecular characterization of nucleoside triphosphate hydrolase isozymes from the parasitic protozoan Toxoplasma gondii. J. Exp. Psychol. Gen. 1995, 270, 11391–11397. [Google Scholar] [CrossRef]
- Coutinho-Silva, R.; Morandini, A.; Savio, L.B. The role of P2X7 receptor in infectious inflammatory diseases and the influence of ectonucleotidases. Biomed. J. 2014, 37, 169. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.S.; Costales, M.G.; Cavanaugh, C.; Williams, K. Extracellular adenosine generation in the regulation of pro-inflammatory responses and pathogen colonization. Biomolecules 2015, 5, 775–792. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, A.L.; Sarti, A.C.; Di Virgilio, F. Extracellular nucleotides and nucleosides as signalling molecules. Immunol. Lett. 2018. [Google Scholar] [CrossRef] [PubMed]
- Dunay, I.R.; Sibley, L.D. Monocytes mediate mucosal immunity to Toxoplasma gondii. Curr. Opin. Immunol. 2010, 22, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Säve, S.; Mohlin, C.; Vumma, R.; Persson, K. Activation of adenosine A2Areceptors inhibits neutrophil transuroepithelial migration. Infect. Immun. 2011, 79, 3431–3437. [Google Scholar] [CrossRef] [PubMed]
- Mahamed, D.A.; Toussaint, L.E.; Bynoe, M.S. CD73-generated adenosine is critical for immune regulation during Toxoplasma gondii infection. Infect. Immun. 2015, 83, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Denkers, E.Y.; Butcher, B.A. Sabotage and exploitation in macrophages parasitized by intracellular protozoans. Trends Parasitol. 2005, 21, 35–41. [Google Scholar] [CrossRef]
- Chaves, S.P.; Torres-Santos, E.C.; Marques, C.; Figliuolo, V.R.; Persechini, P.M.; Coutinho-Silva, R.; Rossi-Bergmann, B. Modulation of P2X7purinergic receptor in macrophages by Leishmania amazonensis and its role in parasite elimination. Microbes Infect. 2009, 11, 842–849. [Google Scholar] [CrossRef]
- Chaves, M.M.; Marques-da-Silva, C.; Monteiro, A.P.T.; Canetti, C.; Coutinho-Silva, R. Leukotriene B4 modulates P2X7 receptor-mediated Leishmania amazonensis elimination in murine macrophages. J. Immunol. 2014, 192, 4765–4773. [Google Scholar] [CrossRef]
- Jacobson, K.A.; Balasubramanian, R.; Deflorian, F.; Gao, Z.-G. G protein-coupled adenosine (P1) and P2Y receptors: Ligand design and receptor interactions. Purinergic Signal. 2012, 8, 419–436. [Google Scholar] [CrossRef] [PubMed]
- Marques-da-Silva, C.; Chaves, M.M.; Chaves, S.P.; Figliuolo, V.R.; Meyer-Fernandes, J.R.; Corte-Real, S.; Lameu, C.; Ulrich, H.; Ojcius, D.M.; Rossi-Bergmann, B.; et al. Infection with Leishmania amazonensis upregulates purinergic receptor expression and induces host-cell susceptibility to UTP-mediated apoptosis. Cell. Microbiol. 2011, 13, 1410–1428. [Google Scholar] [CrossRef] [PubMed]
- Arora, K.; Rai, A.K. Dependence of Leishmania parasite on host derived ATP: An overview of extracellular nucleotide metabolism in parasite. J. Parasit. Dis. 2018. [Google Scholar] [CrossRef]
- Rai, A.K.; Thakur, C.P.; Velpandian, T.; Sharma, S.K.; Ghosh, B.; Mitra, D.K. High concentration of adenosine in human visceral leishmaniasis despite increased ADA and decreased CD73. Parasite Immunol. 2011, 33, 632–636. [Google Scholar] [CrossRef] [PubMed]
- De Almeida Marques-da-Silva, E.; de Oliveira, J.C.; Figueiredo, A.B.; de Souza Lima Júnior, D.; Carneiro, C.M.; Rangel Fietto, J.L.; Crocco Afonso, L.C. Extracellular nucleotide metabolism in Leishmania: Influence of adenosine in the establishment of infection. Microbes Infect. 2008, 10, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.H.F.; Sacramento, L.A.; Quirino, G.F.S.; Ferreira, M.D.; Benevides, L.; Santana, A.K.M.; Cunha, F.Q.; Almeida, R.P.; Silva, J.S.; Carregaro, V. Leishmania infantum parasites subvert the host inflammatory response through the adenosine A2A receptor to promote the establishment of infection. Front. Immunol. 2017, 8, 815. [Google Scholar] [CrossRef] [PubMed]
- Mahaut-smith, M.P.; Jones, S.; Evans, R.J. The P2X1 receptor and platelet function. Purinergic Signal. 2011, 7, 341–356. [Google Scholar] [CrossRef]
- Angchaisuksiri, P. Coagulopathy in malaria. Thromb. Res. 2014, 133, 5–9. [Google Scholar] [CrossRef]
- Idzko, M.; Ferrari, D.; Eltzschig, H.K. Nucleotide signalling during inflammation. NIH Public Access 2014, 509, 310–317. [Google Scholar] [CrossRef]
- Manohar, M.; Hirsh, M.I.; Chen, Y.; Woehrle, T.; Karande, A.A.; Junger, W.G. ATP release and autocrine signaling through P2X4 receptors regulate T cell activation. J. Leukoc. Biol. 2012, 92, 787–794. [Google Scholar] [CrossRef]
- Lazarowski, E.R. Vesicular and conductive mechanisms of nucleotide release. Purinergic Signal. 2012, 8, 359–373. [Google Scholar] [CrossRef] [PubMed]
- Yegutkin, G.G. Nucleotide- and nucleoside-converting ectoenzymes: Important modulators of purinergic signalling cascade. Biochim. Biophys. Acta Mol. Cell Res. 2008, 1783, 673–694. [Google Scholar] [CrossRef] [PubMed]
- Praetorius, H.A.; Leipziger, J. ATP release from non-excitable cells. Purinergic Signal. 2009, 5, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Sabirov, R.Z.; Merzlyak, P.G. Plasmalemmal VDAC controversies and maxi-anion channel puzzle. Biochim. Biophys. Acta Biomembr. 2012, 1818, 1570–1580. [Google Scholar] [CrossRef] [PubMed]
- Dahl, G. ATP release through pannexon channels. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370, 20140191. [Google Scholar] [CrossRef] [PubMed]
- Huber, S.M. Purinoceptor signaling in malaria-infected erythrocytes. Microbes Infect. 2012, 14, 779–786. [Google Scholar] [CrossRef]
- Sabirov, R.Z.; Okada, Y. ATP release via anion channels. Purinergic Signal. 2005, 1, 311–328. [Google Scholar] [CrossRef]
- Marginedas-Freixa, I.; Alvarez, C.L.; Moras, M.; Leal Denis, M.F.; Hattab, C.; Halle, F.; Bihel, F.; Mouro-Chanteloup, I.; Lefevre, S.D.; Le Van Kim, C.; et al. Human erythrocytes release ATP by a novel pathway involving VDAC oligomerization independent of pannexin-1. Sci. Rep. 2018, 8, 11384. [Google Scholar] [CrossRef]
- Capella-Gutierrez, S.; Marcet-Houben, M.; Gabaldón, T. Phylogenomics supports microsporidia as the earliest diverging clade of sequenced fungi. BMC Biol. 2012, 10, 47. [Google Scholar] [CrossRef]
- Didier, E.S.; Weiss, L.M. Microsporidiosis: Not just in AIDS patients. Curr. Opin. Infect. Dis. 2011, 24, 490–495. [Google Scholar] [CrossRef]
- Nakjang, S.; Williams, T.A.; Heinz, E.; Watson, A.K.; Foster, P.G.; Sendra, K.M.; Heaps, S.E.; Hirt, R.P.; Embley, T.M. Reduction and expansion inmicrosporidian genome evolution: New insights from comparative genomics. Genome Biol. Evol. 2013, 5, 2285–2303. [Google Scholar] [CrossRef] [PubMed]
- Dean, P.; Sendra, K.M.; Williams, T.A.; Watson, A.K.; Major, P.; Nakjang, S.; Kozhevnikova, E.; Goldberg, A.V.; Kunji, E.R.S.; Hirt, R.P.; et al. Transporter gene acquisition and innovation in the evolution of Microsporidia intracellular parasites. Nat. Commun. 2018, 9, 1709. [Google Scholar] [CrossRef] [PubMed]
- Heinz, E.; Hacker, C.; Dean, P.; Mifsud, J.; Goldberg, A.V.; Williams, T.A.; Nakjang, S.; Gregory, A.; Hirt, R.P.; Lucocq, J.M.; et al. Plasma membrane-located purine nucleotide transport proteins are key components for host exploitation by microsporidian intracellular parasites. PLoS Pathog. 2014, 10, e1004547. [Google Scholar] [CrossRef] [PubMed]
- Schmitz-Esser, S.; Linka, N.; Collingro, A.; Beier, C.L.; Neuhaus, H.E.; Wagner, M.; Horn, M. ATP/ADP translocases: A common feature of obligate intracellular amoebal symbionts related to Chlamydiae and Rickettsiae. J. Bacteriol. 2004, 186, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Bouyer, G.; Cueff, A.; Egée, S.; Kmiecik, J.; Maksimova, Y.; Glogowska, E.; Gallagher, P.G.; Thomas, S.L.Y. Erythrocyte peripheral type benzodiazepine receptor/voltage-dependent anion channels are upregulated by Plasmodium falciparum. Blood 2011, 118, 2305–2312. [Google Scholar] [CrossRef] [PubMed]
- Marginedas-Freixa, I.; Alvarez, C.L.; Moras, M.; Hattab, C.; Bouyer, G.; Chene, A.; Lefevre, S.D.; Le Van Kim, C.; Bihel, F.; Schwarzbaum, P.J.; et al. Induction of ATP Release, PPIX Transport, and cholesterol uptake by human red blood cells using a new family of TSPO ligands. Int. J. Mol. Sci. 2018, 19, 3098. [Google Scholar] [CrossRef] [PubMed]
- Lutz, P.L.; Kabler, S. Release of adenosine and ATP in the brain of the freshwater turtle (Trachemys scripta) during long-term anoxia. Brain Res. 1997, 769, 281–286. [Google Scholar] [CrossRef]
- Clarke, T.C.; Williams, O.J.S.; Martin, P.E.M.; Evans, W.H. ATP release by cardiac myocytes in a simulated ischaemia model. Inhibition by a connexin mimetic and enhancement by an antiarrhythmic peptide. Eur. J. Pharmacol. 2009, 605, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Lim To, W.K.; Kumar, P.; Marshall, J.M. Hypoxia is an effective stimulus for vesicular release of ATP from human umbilical vein endothelial cells. Placenta 2015, 36, 759–766. [Google Scholar] [CrossRef]
- Rajani, V.; Zhang, Y.; Jalubula, V.; Rancic, V.; SheikhBahaei, S.; Zwicker, J.D.; Pagliardini, S.; Dickson, C.T.; Ballanyi, K.; Kasparov, S.; et al. Release of ATP by pre-Bötzinger complex astrocytes contributes to the hypoxic ventilatory response via a Ca2+-dependent P2Y1 receptor mechanism. J. Physiol. 2018, 596, 3245–3269. [Google Scholar] [CrossRef]
- Sridharan, M.; Adderley, S.P.; Bowles, E.A.; Egan, T.M.; Stephenson, A.H.; Ellsworth, M.L.; Sprague, R.S. Pannexin 1 is the conduit for low oxygen tension-induced ATP release from human erythrocytes. AJP Hear. Circ. Physiol. 2010, 299, H1146–H1152. [Google Scholar] [CrossRef] [PubMed]
- Maguire, M.H.; Lukas, M.C.; Rettie, J.F. Adenine Nucleotide salvage synthesis pathways of adenosine salvage in the rat heart. Biochim. Biophys. Acta 1972, 262, 108–115. [Google Scholar] [CrossRef]
- Paes-Vieira, L.; Gomes-Vieira, A.L.; Meyer-Fernandes, J.R. NTPDase activities: Possible roles on Leishmania spp. infectivity and virulence. Cell Biol. Int. 2018, 42, 670–682. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Saha, A.K.; Mukhopadhyay, N.K.; Glew, R.H. A cyclic nucleotide-independent protein kinase in Leishmania donovani. Biochem. J. 1986, 240, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Robson, S.C.; Sévigny, J.; Zimmermann, H. The E-NTPDase family of ectonucleotidases: Structure function relationships and pathophysiological significance. Purinergic Signal. 2006, 2, 409–430. [Google Scholar] [CrossRef] [PubMed]
- Franco, R.; Pacheco, R.; Gatell, J.M.; Gallart, T.; Lluis, C. Enzymatic and extraenzymatic role of adenosine deaminase 1 in T-cell-dendritic cell contacts and in alterations of the immune function. Crit. Rev. Immunol. 2007, 27, 495–509. [Google Scholar] [CrossRef] [PubMed]
- Yegutkin, G.G.; Samburski, S.S.; Jalkanen, S. Soluble purine-converting enzymes circulate in human blood and regulate extracellular ATP level via counteracting pyrophosphatase and phosphotransfer reactions. FASEB J. 2003, 17, 1328–1330. [Google Scholar] [CrossRef]
- Boison, D. Adenosine kinase: Exploitation for therapeutic gain. Pharmacol. Rev. 2013, 65, 906–943. [Google Scholar] [CrossRef]
- Virtanen, S.S.; Kukkonen-Macchi, A.; Vainio, M.; Elima, K.; Harkonen, P.L.; Jalkanen, S.; Yegutkin, G.G. Adenosine Inhibits tumor cell invasion via receptor-independent mechanisms. Mol. Cancer Res. 2014, 12, 1863–1874. [Google Scholar] [CrossRef]
- Plesner, L. Ecto-ATPases: Identities and functions. Int. Rev. Cytol. 1995, 158, 141–214. [Google Scholar]
- Kukulski, F.; Levesque, S.A.; Lavoie, E.G.; Lecka, J.; Bigonnesse, F.; Knowles, A.F.; Robson, S.C.; Kirley, T.L.; Sevigny, J. Erratum: Comparative hrydrolysis of P2 receptor agonists by NTPDases 1, 2, 3 and 8. (Purinergic Signalling (2005) vol. 1 (2) (193-204)). Purinergic Signal. 2005, 1, 293. [Google Scholar] [CrossRef]
- Handa, M.; Guidotti, G. Purification and cloning of a soluble ATP-diphosphohydrolase (Apyrase) from potato tubers (Solanum tuberosum). Biochem. Biophys. Res. Commun. 1996, 218, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Schulte Am Esch, J.; Sévigny, J.; Kaczmarek, E.; Siegel, J.B.; Imai, M.; Koziak, K.; Beaudoin, A.R.; Robson, S.C. Structural elements and limited proteolysis of CD39 influence ATP diphosphohydrolase activity. Biochemistry 1999, 38, 2248–2258. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.J.; Choi, L.E.; Guidotti, G. N-linked oligosaccharides affect the enzymatic activity of CD39: Diverse interactions between seven N-linked glycosylation sites. Mol. Biol. Cell 2005, 16, 1661–1672. [Google Scholar] [CrossRef] [PubMed]
- Knowles, A.F. The GDA1_CD39 superfamily: NTPDases with diverse functions. Purinergic Signal. 2011, 7, 21–45. [Google Scholar] [CrossRef] [PubMed]
- Kirley, T.L.; Crawford, P.A.; Smith, T.M. The structure of the nucleoside triphosphate diphosphohydrolases (NTPDases) as revealed by mutagenic and computational modeling analyses. Purinergic Signal. 2006, 2, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Mandapathil, M.; Hilldorfer, B.; Szczepanski, M.J.; Czystowska, M.; Szajnik, M.; Ren, J.; Lang, S.; Jackson, E.K.; Gorelik, E.; Whiteside, T.L. Generation and accumulation of immunosuppressive adenosine by human CD4+CD25highFOXP3+ regulatory T cells. J. Biol. Chem. 2010, 285, 7176–7186. [Google Scholar] [CrossRef]
- Heine, P.; Braun, N.; Sévigny, J.; Robson, S.C.; Servos, J.; Zimmermann, H. The C-terminal cysteine-rich region dictates specific catalytic properties in chimeras of the ectonucleotidases NTPDase1 and NTPDase2. Eur. J. Biochem. 2001, 268, 364–373. [Google Scholar] [CrossRef]
- Zimmermann, H. Ectonucleotidases: Some recent developments and a note on nomenclature. Drug Dev. Res. 2001, 52, 44–56. [Google Scholar] [CrossRef]
- EMBL. SMART-Simple Modular Architecture Research Tool; 1996–2014 Health Sciences Library System, University of Pittsburgh: Pittsburgh, PA, USA.
- Nakaar, V.; Samuel, B.U.; Ngo, E.O.; Joiner, K.A. Targeted reduction of nucleoside triphosphate hydrolase by antisense RNA inhibits Toxoplasma gondii proliferation. J. Biol. Chem. 1999, 274, 5083–5087. [Google Scholar] [CrossRef]
- Sansom, F.M.; Robson, S.C.; Hartland, E.L. Possible effects of microbial ecto-nucleoside triphosphate diphosphohydrolases on host-pathogen interactions. Microbiol. Mol. Biol. Rev. 2008, 72, 765–781. [Google Scholar] [CrossRef] [PubMed]
- Berrêdo-Pinho, M.; Peres-Sampaio, C.E.; Chrispim, P.P.M.; Belmont-Firpo, R.; Lemos, A.P.; Martiny, A.; Vannier-Santos, M.A.; Meyer-Fernandes, J.R. A Mg-dependent ecto-ATPase in Leishmania amazonensis and its possible role in adenosine acquisition and virulence. Arch. Biochem. Biophys. 2001, 391, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, C.M.; Martins-Duarte, E.S.; Ferraro, R.B.; Fonseca de Souza, A.L.; Gomes, M.T.; Lopes, A.H.C.S.; Vannier-Santos, M.A.; Santos, A.L.S.; Meyer-Fernandes, J.R. Leishmania amazonensis: Biological and biochemical characterization of ecto-nucleoside triphosphate diphosphohydrolase activities. Exp. Parasitol. 2006, 114, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Sansom, F.M.; Newton, H.J.; Crikis, S.; Cianciotto, N.P.; Cowan, P.J.; d’Apice, A.J.F.; Hartland, E.L. A bacterial ecto-triphosphate diphosphohydrolase similar to human CD39 is essential for intracellular multiplication of Legionella pneumophila. Cell. Microbiol. 2007, 9, 1922–1935. [Google Scholar] [CrossRef] [PubMed]
- Nakaar, V.; Beckers, C.J.M.; Polotsky, V.; Joiner, K.A. Basis for substrate specificity of the Toxoplasma gondii nucleoside triphosphate hydrolase. Mol. Biochem. Parasitol. 1998, 97, 209–220. [Google Scholar] [CrossRef]
- Silverman, J.A.; Qi, H.; Riehl, A.; Beckers, C.; Nakaar, V.; Joiner, K.A. Induced activation of the Toxoplasma gondii nucleoside triphosphate hydrolase leads to depletion of host cell ATP levels and rapid exit of intracellular parasites from infected cells. J. Biol. Chem. 1998, 273, 12352–12359. [Google Scholar] [CrossRef] [PubMed]
- Bermudes, D.; Peck, K.R.; Afifi, M.A.; Beckers, C.J.M.; Joiner, K.A. Tandemly repeated genes encode nucleoside triphosphate hydrolase isoforms secreted into the parasitophorous vacuole of Toxoplasma gondii. J. Biol. Chem. 1994, 269, 29252–29260. [Google Scholar] [PubMed]
- Sibley, L.D.; Niesman, I.R.; Asai, T.; Takeuchi, T. Toxoplasma gondii: Secretion of a potent nucleoside triphosphate hydrolase into the parasitophorous vacuole. Exp. Parasitol. 1994, 79, 301–311. [Google Scholar] [CrossRef]
- Kikuchi, T.; Furuta, T.; Kojima, S. Membrane localization and demonstration of isoforms of nucleoside triphosphate hydrolase from Toxoplasma gondii. Parasitology 2001, 122, S0031182000007101. [Google Scholar] [CrossRef]
- Montalbetti, N.; Leal Denis, M.F.; Pignataros, O.P.; Kobatake, E.; Lazarowski, E.R.; Schwarzbaum, P.J. Homeostasis of extracellular ATP in human erythrocytes. J. Biol. Chem. 2011, 286, 38397–38407. [Google Scholar] [CrossRef]
- Pastor-Fernández, I.; Regidor-Cerrillo, J.; Álvarez-García, G.; Marugán-Hernández, V.; García-Lunar, P.; Hemphill, A.; Ortega-Mora, L.M. The tandemly repeated NTPase (NTPDase) from Neospora caninum is a canonical dense granule protein whose RNA expression, protein secretion and phosphorylation coincides with the tachyzoite egress. Parasites Vectors 2016, 9, 352. [Google Scholar] [CrossRef] [PubMed]
- Nagajyothi, F.; Machado, F.S.; Burleigh, B.A.; Jelicks, L.A.; Scherer, E.; Mukherjee, S.; Lisanti, M.P.; Weiss, L.M.; Garg, N.J.; Tanowitz, H.B. Mechanisms of Trypanosoma cruzi persistence in Chagas disease. Cell Microbiol. 2013, 14, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.F.; Pôssa, M.A.S.; Bastos, M.S.; Guedes, P.M.M.; Almeida, M.R.; DeMarco, R.; Verjovski-Almeida, S.; Bahia, M.T.; Fietto, J.L.R. Influence of ecto-nucleoside triphosphate diphosphohydrolase activity on Trypanosoma cruzi infectivity and virulence. PLoS Negl. Trop. Dis. 2009, 3, e387. [Google Scholar] [CrossRef] [PubMed]
- WHO Chagas Disease. American Trypanosomiasis. Available online: https://www.who.int/chagas/en/ (accessed on 19 December 2018).
- Fietto, J.L.R.; DeMarco, R.; Nascimento, I.P.; Castro, I.M.; Carvalho, T.M.U.; De Souza, W.; Bahia, M.T.; Alves, M.J.M.; Verjovski-Almeida, S. Characterization and immunolocalization of an NTP diphosphohydrolase of Trypanosoma cruzi. Biochem. Biophys. Res. Commun. 2004, 316, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Schnurr, M.; Then, F.; Galambos, P.; Scholz, C.; Siegmund, B.; Endres, S.; Eigler, A. Extracellular ATP and TNF- synergize in the activation and maturation of human dendritic cells. J. Immunol. 2000, 165, 4704–4709. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, H. Extracellular metabolism of ATP and other nucleotides. Naunyn. Schmiedebergs. Arch. Pharmacol. 2000, 362, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.C.; Novaes, R.D.; Cupertino, M.C.; Bastos, D.S.S.; Klein, R.C.; Silva, E.A.M.; Fietto, J.L.R.; Talvani, A.; Bahia, M.T.; Oliveira, L.L. Concomitant benznidazole and suramin chemotherapy in mice infected with a virulent strain of Trypanosoma cruzi. Antimicrob. Agents Chemother. 2015, 59, 5999–6006. [Google Scholar] [CrossRef] [PubMed]
- Bisaggio, D.F.R.; Peres-Sampaio, C.E.; Meyer-Fernandes, J.R.; Souto-Padrón, T. Ecto-ATPase activity on the surface of Trypanosoma cruzi and its possible role in the parasite-host cell interaction. Parasitol. Res. 2003, 91, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Mariotini-Moura, C.; Bastos, M.S. e.; de Castro, F.F.; Trindade, M.L.; de Souza Vasconcellos, R.; Neves-do-Valle, M.A.A.; Moreira, B.P.; de Freitas Santos, R.; de Oliveira, C.M.; Cunha, L.C.S.; et al. Trypanosoma cruzi nucleoside triphosphate diphosphohydrolase 1 (TcNTPDase-1) biochemical characterization, immunolocalization and possible role in host cell adhesion. Acta Trop. 2014, 130, 140–147. [Google Scholar] [CrossRef]
- de Souza Leite, M.; Thomaz, R.; Fonseca, F.V.; Panizzutti, R.; Vercesi, A.E.; Meyer-Fernandes, J.R. Trypanosoma brucei brucei: Biochemical characterization of ecto-nucleoside triphosphate diphosphohydrolase activities. Exp. Parasitol. 2007, 115, 315–323. [Google Scholar] [CrossRef]
- Matthews, K.R. The developmental cell biology of Trypanosoma brucei. J. Cell Sci. 2005, 118, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Berriman, M.; Ghedin, E.; Hertz-Fowler, C.; Blandin, G.; Renauld, H.; Bartholomeu, D.C.; Lennard, N.J.; Caler, E.; Hamlin, N.E.; Haas, B.; et al. The genome of the African trypanosome Trypanosoma brucei. Science 2005, 309, 416–422. [Google Scholar] [CrossRef]
- Balaña-Fouce, R.; Reguera, R.M. RNA interference in Trypanosoma brucei: A high-throughput engine for functional genomics in trypanosomatids? TRENDS Parasitol. 2007, 23, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Peacock, C.S.; Seeger, K.; Harris, D.; Murphy, L.; Ruiz, J.C.; Quail, M.A.; Peters, N.; Adlem, E.; Tivey, A.; Aslett, M.; et al. 2008 Comparative genomic analysis of three Leishmania species that cause diverse human disease. Nat. Genet. 2008, 39, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Sansom, F.M.; Ralton, J.E.; Sernee, M.F.; Cohen, A.M.; Hooker, D.J.; Hartland, E.L.; Naderer, T.; McConville, M.J. Golgi-located NTPDase1 of Leishmania major is required for lipophosphoglycan elongation and normal lesion development whereas secreted NTPDase2 is dispensable for virulence. PLoS Negl. Trop. Dis. 2014, 8, e3402. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Fernandes, J.R.; Dutra, P.M.L.; Rodrigues, C.O.; Saad-Nehme, J.; Lopes, A.H.C.S. Mg-dependent ecto-ATPase activity in Leishmania tropica. Arch. Biochem. Biophys. 1997, 341, 40–46. [Google Scholar] [CrossRef]
- Borges-Pereira, L.; Meissner, K.A.; Wrenger, C.; Garcia, C.R.S. Plasmodium falciparum GFP-E-NTPDase expression at the intraerythrocytic stages and its inhibition blocks the development of the human malaria parasite. Purinergic Signal. 2017, 13, 267–277. [Google Scholar] [CrossRef]
- Dos Santos-Rodrigues, A.; Grañé-Boladeras, N.; Bicket, A.; Coe, I.R. Nucleoside transporters in the purinome. Neurochem. Int. 2014, 73, 229–237. [Google Scholar] [CrossRef]
- Landfear, S.M.; Ullman, B.; Carter, N.S.; Sanchez, M.A. Nucleoside and nucleobase transporters in parasitic protozoa. Eukaryot. Cell 2004, 3, 245–254. [Google Scholar] [CrossRef]
- Yao, S.Y.M.; Ng, A.M.L.; Cass, C.E.; Baldwin, S.A.; Young, J.D. Nucleobase transport by human equilibrative nucleoside transporter 1 (hENT1). J. Biol. Chem. 2011, 286, 32552–32562. [Google Scholar] [CrossRef]
- Landfear, S.M. Nutrient transport and pathogenesis in selected parasitic protozoa. Eukaryot. Cell 2011, 10, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Boswell-Casteel, R.C.; Hays, F.A. Equilibrative nucleoside transporters—A review. Nucleosides Nucleotides Nucleic Acids 2016, 36, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Löffler, M.; Morote-Garcia, J.C.; Eltzschig, S.A.; Coe, I.R.; Eltzschig, H.K. Physiological roles of vascular nucleoside transporters. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Hyde, R.J.; Cass, C.E.; Young, J.D.; Baldwin, S.A. The ENT family of eukaryote nucleoside and nucleobase transporters: Recent advances in the investigation of structure/function relationships and the identification of novel isoforms. Mol. Membr. Biol. 2001, 18, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Dean, P.; Major, P.; Nakjang, S.; Hirt, R.P.; Embley, T.M. Transport proteins of parasitic protists and their role in nutrient salvage. Front. Plant Sci. 2014, 5, 153. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.A.; Tryon, R.; Green, J.; Boor, I.; Landfear, S.M. Six related nucleoside/nucleobase transporters from Trypanosoma brucei exhibit distinct biochemical functions. J. Biol. Chem. 2002, 277, 21499–21504. [Google Scholar] [CrossRef] [PubMed]
- Mäser, P.; Sütterlin, C.; Kralli, A.; Kaminsky, R. A nucleoside transporter from Trypanosoma brucei involved in drug resistance. Science 1999, 285, 242–244. [Google Scholar]
- Carter, N.S.; Mamoun, C.B.; Liu, W.; Silva, E.O.; Landfear, S.M.; Goldberg, D.E.; Ullman, B. Isolation and functional characterization of the PfNT1 nucleoside transporter gene from Plasmodium falciparum. J. Biol. Chem. 2000, 275, 10683–10691. [Google Scholar] [CrossRef]
- Carter, N.S.; Landfear, S.M.; Ullman, B. Nucleoside transporters of parasitic protozoa. Trends Parasitol. 2001, 17, 142–145. [Google Scholar] [CrossRef]
- Berg, M.; Van der Veken, P.; Goeminne, A.; Haemers, A.; Augustyns, K. Inhibitors of the purine salvage pathway: A valuable approach for antiprotozoal chemotherapy? Curr. Med. Chem. 2010, 17, 2456–2481. [Google Scholar] [CrossRef]
- Cassera, M.B.; Zhang, Y.; Hazleton, K.Z.; Schramm, V.L. Purine and pyrimidine pathways as targets in Plasmodium falciparum. Curr. Top. Med. Chem. 2011, 4511, 2103–2115. [Google Scholar] [CrossRef]
- Hassan, H.F.; Coombs, G.H. Purine and pyrimidine metabolism in parasitic protozoa. FEMS Microbiol. Lett. 1988, 54, 47–83. [Google Scholar]
- Riegelhaupt, P.M.; Cassera, M.B.; Fröhlich, R.F.G.; Hazleton, K.Z.; Hefter, J.J.; Schramm, V.L.; Akabas, M.H. Transport of purines and purine salvage pathway inhibitors by the Plasmodium falciparum equilibrative nucleoside transporter PfENT1. Mol. Biochem. Parasitol. 2010, 169, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Schwab, J.C.; Beckers, C.J.M.; Joiner, K.A. The parasitophorous vacuole membrane surrounding intracellular Toxoplasma gondii funtions as a molecular sieve. Cell Biol. 1994, 91, 509–513. [Google Scholar]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lauri, N.; Bazzi, Z.; Alvarez, C.L.; Leal Denis, M.F.; Schachter, J.; Herlax, V.; Ostuni, M.A.; Schwarzbaum, P.J. ATPe Dynamics in Protozoan Parasites. Adapt or Perish. Genes 2019, 10, 16. https://doi.org/10.3390/genes10010016
Lauri N, Bazzi Z, Alvarez CL, Leal Denis MF, Schachter J, Herlax V, Ostuni MA, Schwarzbaum PJ. ATPe Dynamics in Protozoan Parasites. Adapt or Perish. Genes. 2019; 10(1):16. https://doi.org/10.3390/genes10010016
Chicago/Turabian StyleLauri, Natalia, Zaher Bazzi, Cora L. Alvarez, María F. Leal Denis, Julieta Schachter, Vanesa Herlax, Mariano A. Ostuni, and Pablo J. Schwarzbaum. 2019. "ATPe Dynamics in Protozoan Parasites. Adapt or Perish" Genes 10, no. 1: 16. https://doi.org/10.3390/genes10010016
APA StyleLauri, N., Bazzi, Z., Alvarez, C. L., Leal Denis, M. F., Schachter, J., Herlax, V., Ostuni, M. A., & Schwarzbaum, P. J. (2019). ATPe Dynamics in Protozoan Parasites. Adapt or Perish. Genes, 10(1), 16. https://doi.org/10.3390/genes10010016