Abstract
Nitrogen (N) is a key regulator in the growth of tea plants and the synthesis of amino acids (AAs) and related secondary metabolites, thus affecting the yield and quality of tea leaves. Increased N fertilization significantly improves the yield and quality of tea plants, but applying excess nitrogen wastes resources and causes pollution problems. Herein, we employed morphological, physiological, metabolomic, and RNA-seq methods to study the response of ‘Ziyan’ roots to high N. High N supply induced an increase in arginine (Arg), asparagine (Asn), and glutamine (Gln) in roots and simultaneously decreased sucrose, polyphenols, and caffeine contents. High N reduced the length, volume, number, and activity of the roots by 10.63%, 25.00%, 26.95%, and 14.50%, respectively, which inhibited ‘Ziyan’ root growth, probably by disturbing the regulation of carbon and nitrogen metabolism in the tea plant. According to the Kyoto Encyclopedia of Genes and Genome (KEGG) enrichment analysis, AAs, flavonoids, and flavonol-related pathways were relatively active after supplying high N. In addition, the transcriptome analysis identified NRT1/PTR and GOGAT as key genes, and the transcription factors (e.g., AP2/ERF, MYB, and WRKY) and auxins were actively involved in the high N stress response of ‘Ziyan’ roots. These findings will help us understand the adaptive mechanism of high N supply in tea tree roots and provide a reference for guiding the fertilization of ‘Ziyan’ tea plants.
1. Introduction
Tea (Camellia sinensis (L.) O. Kuntze) is grown as an important cash crop in over 60 countries. Tea is one of the most popular non-alcoholic beverages in the world, and the tea leaf industry is featured as one of the most important commercial crops in China and a pillar industry for farmers to increase their income. Nitrogen (N) is closely related to the growth, yield, and quality of tea plants [1]. Soil N deficiency significantly reduces tea leaf yield and quality [2], while applying N is beneficial for yield and quality [3,4]. However, applying excess N to increase yield has been commonly observed, particularly in Sichuan, China. Nevertheless, excess N application reduces tea leaf quality, wastes the resource, and causes pollution problems, seriously restricting the sustainable development of the tea industry [5,6]. Therefore, methods to increase the N absorption efficiency of tea plants for breeding new varieties are needed.
The roots of tea plants are the main N uptake organ and an important site for the synthesis of amino acids (AAs) and secondary metabolites. The morphological and physiological characteristics of the roots affect the growth and development of the aboveground parts of tea plants, thus affecting their growth and quality [7]. Tea plants have root morphology and root uptake systems that are strongly affected by the N level and prefer ammonium–nitrogen as the N source [8,9]. N deficiency promotes or inhibits root growth [10,11], and this difference may be related to the degree of N deficiency. Applying excess N does not continue to increase the number of adventitious roots, but it decreases root morphological parameters and eventually inhibits root growth [12]. The length and biomass of poplar roots increases with a drop in N effectiveness [13]. The most important regulatory factors in the root N uptake system include transcription factors (TFs), phytohormones, translocators, and AA metabolic enzymes, and phytohormones and AA metabolic enzymes are key factors regulating root growth [14,15,16]. Phytohormones are involved in regulating root growth and further enhance the resistance to stress. Glutamate (Glu) is a precursor of various AAs, which is involved in ammonium assimilation and affects the synthesis of secondary metabolites (e.g., polyphenols and alkaloids). Additionally, redistributed AAs alter the growth and development of plants [17,18].
‘Ziyan’ is a new variety of purple-leaved tea plant, which is rich in anthocyanins (3%) in its tender leaves, and it is very popular among consumers for its health benefits and unique flavor [19,20]. Interestingly, the aboveground and root growth of ‘Ziyan’ tea plants under high N supply is weaker than that of plants with a moderate N application. It has been demonstrated that gene expression and metabolite accumulation are related to the mechanisms regulating the root growth of tea plants under low N or N deficient conditions [8,14,21], whereas the regulatory mechanism of tea plant root growth under high N remains unclear. In this study, we integrated morphological, physiological, and non-targeted metabolomics analyses to investigate the correlation between a high N supply and the accumulation of AAs and secondary metabolites in roots. We also examined the regulatory mechanism of phytohormone and TFs in tea plant root growth. This study provides a reference for the precise fertilization of ‘Ziyan’ tea plants and provides insight into the mechanism of tea leaf growth under high N conditions.
2. Materials and Methods
2.1. Plant Materials and Treatment
One-year-old cuttings of the purple tea cultivar “Ziyan” were purchased from Sichuan Muchuan Yizhichun Tea Co., Ltd. (Leshan, China) and pre-cultivated in the Fifth Education Greenhouse at the Chengdu Campus of Sichuan Agricultural University (Chengdu, China). After 7 days of culture in pure water, a 1/4 Xiaoxi Maoyi nutrient solution (N is 1.427 mmol · L−1, the ratio of ammonium–nitrogen to nitrate–nitrogen was 3:1, pH = 5.0) was used to culture for 7 days, followed by a 1/2 tea tree nutrient solution [22]. The nutrient solution was replaced every 7 days until new roots grew. When the new roots were fully grown, the healthy tea seedlings with the same growing trend were selected and treated at three N levels: low nitrogen (0.3 mmol·L−1, LN), medium nitrogen (1.5 mmol·L−1, MN), and high nitrogen (4.5 mmol·L−1, HN), using 10 pots per treatment and 6 tea seedlings per pot. The tea plants were cultured in 2 L of nutrient solution, and the nutrient solution was exchanged every 7 d. An air pump was used for 24 h of continuous ventilation. The nutrient solution formula included macro-elements (P 0.1, K 1.0, Ca 0.8, Mg 0.4 mmol · L−1) and trace elements (EDTA Fe 6.3, Mn 1.5, Zn 1.0, Cu 0.2, B 10, Mo 0.5, and Mo 50 μmol·L−1). All experiments were conducted under light (25 ± 1 °C, 14 h)/dark (20 ± 1 °C, 10 h), 70% relative humidity, and a light intensity of 200 μmol ·m−2·s−1. The tea seedlings from each treatment (LN, MN, and HN) were taken after 20 days of treatment, and the root morphology and physiological indices were measured. Then, the white absorption roots were collected, immediately frozen in liquid nitrogen, and stored at −80 °C for the transcriptome sequencing analysis. Finally, the white absorption roots were collected and dried in an oven at 85 °C for the root metabolomics analysis.
2.2. Analysis of Root Architecture and Physiological Characteristics
The root system was carefully rinsed with running water. It was cut from the first root of the base, and the root morphology was recorded with an Epson 4990 scanner. The length, surface area, volume, diameter, and number of roots were analyzed using WinRINZO software [23]. The root activity was determined using the triphenyltetrazolium chloride (TTC) method [24]. At the same time, the activities of glutamine synthetase (GS), glutamate dehydrogenase (GDH), and glutamate synthase (GOGAT) were measured in fresh roots from each treatment. The activity of these synthases was determined with colorimetry kits from Nanjing Jiancheng Technology Co., Ltd. (Nanjing, China).
2.3. Ultra-Performance Liquid Chromatography to Quadrupole Time-of-Flight Mass Spectrometry (UPLC-Q-TOF/MS)-Based Metabolomics Analysis
The metabolites of the tea plant root samples were extracted as previously reported with minor modifications [25]. Briefly, root powder samples (100 mg) from each group were ground, followed by extracting the powder in 15 mL of 70% methanol solution (v/v) for 30 min at 70 °C. Then, the mixture was centrifuged at 5000 rpm for 10 min to collect the supernatant. Each sample was centrifuged 6 times. Three QC samples were set (all samples were mixed in equal amounts) to evaluate the stability and reproducibility of the metabolomics analysis. The ultrahigh performance liquid chromatography (UPLC)-Q-Extractive/mass spectroscopy (MS) instrument was used for the separation (Thermo Fisher Scientific, Waltham, MA, USA). Metabolites in water were loaded onto a T3 column (1.7 µm, 100 mm × 2.1 mm) and eluted with mobile phase A (aqueous solution containing 0.1% formic acid, v/v) and mobile phase B (aqueous solution containing 0.1% formic acid acetonitrile, v/v) at a flow rate of 0.4 mL/min, and an injection volume of 3 μL. The mobile phase elution gradient and LC-MS conditions were described by Li [26]. The original spectra obtained by the UHPLC-Q-Extractive/MS analysis were matched and extracted using Compound Discoverer 3.2 software. The structural identification of the metabolites was based on metabolomics-related literature [27,28], combined with HMDB, Kyoto Encyclopedia of Genes and Genome (KEGG), and a self-built local metabolite database. The first-level mass number retrieval accuracy was set to 5 ppm.
2.4. Transcriptome Analysis
Total RNA was extracted with Trizol reagent according to the manufacturer’s instructions (Invitrogen, Carlsbad, CA, USA). The quantity, purity, and integrity of the RNA were measured on the Bioanalyzer 2100 (Agilent Technologies, Palo Alto, CA, USA) using the NanoDrop ND-1000 (NanoDrop, Wilmington, DE, USA). Magnetic beads were attached with poly T oligonucleotides in two rounds from total RNA (5 μg), and the poly (A) RNA was purified. Next, divalent cations were used to break the mRNA fragments into smaller pieces under high temperature, and then the split RNA fragment was reverse-transcribed to create the final cDNA library (Illumina, San Diego, CA, USA) according to the protocol with the mRNAseq sample preparation kit. The cDNA library was constructed by Hangzhou Lianchuan Biotechnology Co., Ltd. (Hangzhou, China), sequenced on an Illumina Hiseq4000, and had a 150 bp paired-end reading segment. Root RNA sequencing was performed using three biological repeats.
Clean data were obtained using Cutadapt to trim the low-quality reads, including the adapter, ploy-N (N > 5%), and raw data. Hisa software was used to map valid data to the “Suchazao” genome [29]. Quality control of the sequencing data was performed using Fast QC (version 0.11.6). All assembled non-redundant transcripts were aligned with the common database to functionally annotate all single genes. The functions of different genes were identified by a KEGG signal pathway enrichment analysis (significance threshold: E ≤ 10). The mRNA expression levels were analyzed by calculating the number of fragments per thousand bases (FPKM) mapped read per million exon model [20]. The differentially expressed genes (DEGs) were screened with |log2fc| ≥ 1 and q < 0.05.
2.5. Quantitative Real-Time Polymerase Chain Reaction Analysis
Ten randomly selected genes were selected for gene expression verification by a quantitative real-time polymerase chain reaction (qRT-PCR) analysis. The mRNA was reverse-transcribed into cDNA using the PrimeScript™ RT kit with the gDNA eraser (Takara, Kyoto, Japan), and the NCBI was used to design gene-specific primers. The primers used are listed in Table S1. SYBR Premium (Takara) was used for the qRT-PCR. The PCR reaction system and procedure were carried out with reference to the method of Li [20]. GAPDH was used as the internal reference gene. All tested genes were tested with three biological replicates and three technical replicates. The relative expression of the genes was analyzed by the 2−ΔΔCt (Livak) method.
2.6. Statistical Analysis
The statistical analysis was conducted using SPSS Statistics 20 software (IBM Corp., Chicago, IL, USA). Differences were detected by a one-way analysis of variance and the least significant difference test. A p-value < 0.05 was considered significant.
3. Results
3.1. Morphological and Physiological Traits of ‘Ziyan’ Roots under Different Nitrogen Levels
As shown in Figure 1A,B, the length, surface area, diameter, volume, number, and activity of the roots in the HN group decreased by 10.63%, 7.85%, 10.23%, 25.00%, 26.95%, and 14.50%, respectively, compared with those in the MN group, while the surface area and diameter of the roots in the HN group were not significantly different from the MN group (p < 0.05). All of these indicators were maximized in the LN group, where the content from the surface area and the volume of the roots were 39.39% and 42.59% higher than those in the MN group, respectively (p < 0.05). These results demonstrate that the volume, surface area, length, and number of roots decreased in the HN group, which limited root growth, while root growth in the LN group was facilitated. Similarly, the activities of the roots and N metabolic enzymes (e.g., GS, GOGAT, and GDH) in roots supported these results (Figure 1C).
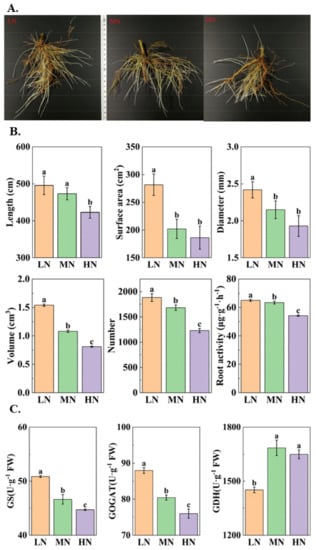
Figure 1.
Root growth (A), morphological characteristics (B) and nitrogen metabolic enzymes (C) under different nitrogen conditions.
3.2. Metabolites of ‘Ziyan’ Roots under Different Nitrogen Levels
The UPLC-Q-TOF/MS metabolomics analysis revealed that 98 of the 117 compounds were the reactants of eight key metabolites that exhibited significant differences (Figure 2A). As shown in Figure 2B, the extraction of total root metabolites and the metabolomics analysis showed good repeatability and stability. The metabolic pathway enrichment analysis of the differentially expressed metabolites revealed that the main metabolic pathways of the 98 compounds (p < 0.05) included aminoacyl-tRNA biosynthesis, flavone and flavonol biosynthesis, arginine biosynthesis, valine, leucine, and isoleucine biosynthesis; alanine, aspartate, and glutamate metabolism; glyoxylate and dicarboxylate metabolism, glycine, serine, and threonine metabolism, glycerophospholipid metabolism, and purine metabolism (Figure 2B). These results suggest that some pathways related to the biosynthesis of AAs, flavones, and flavonols were active in ‘Ziyan’ roots under a high N supply.
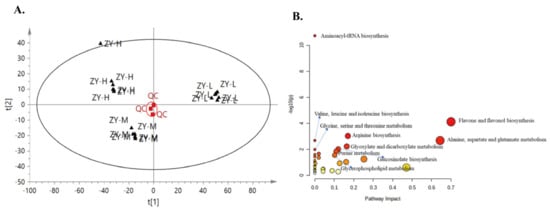
Figure 2.
Principal component analysis score of metabolomics (A) and metabolic pathway enrichment analysis of different compounds (B).
The different levels of AA in the roots directly revealed N assimilation under the different N levels. The main AA metabolic pathways included five AA families (Glu, Asp, Ala, Ser, and aromatic AAs), which are shown in different colors in Figure 3. Furthermore, 16 AAs were detected in our analysis (Figure 3; Table S2). Specifically, the expression of all 12 AAs was significantly upregulated in the HN group compared with the MN group, except isoleucine (Ile), leucine (Leu), phenylalanine (Phe), and N-methyllysine, whereas arginine (Arg), glutamine (Gln), asparagine (Asn), and theanine (Thea) were expressed at relatively higher levels. All 14 types of AAs were downregulated in the LN group, except proline (Pro) and N-methyllysine, whereas Arg, Gln, Thea, Asn, and Ser(Serine) decreased significantly. Additionally, Pro content increased slightly under low and high nitrogen conditions.
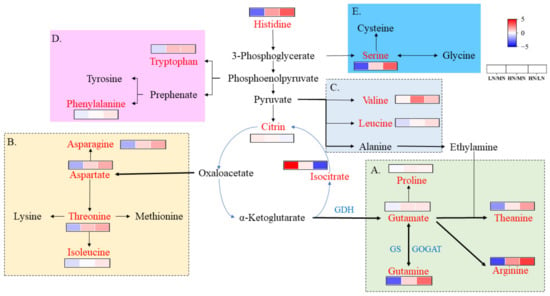
Figure 3.
Changes in free amino acids and organic acids in roots of ‘Ziyan’ under different nitrogen conditions. (A): Glutamate family (light-green frame); (B): aspartate family (yellow frame); (C): alanine family (light-gray frame); (D): aromatic AAs family (purple frame); (E): serine family (blue frame). The relative abundance of metabolites is illustrated on a red (high) to blue (low) scale.
A total of 31 tea polyphenols (Figure 4A,B and Table S2) were detected, including 22 flavonol/flavone glycosides, five flavanols, three dimeric catechins, and one phenolic acid. The contents of these substances increased in the LN group but decreased in the HN group, and only kaempferol-3-rutin, quercetin 3−O−rutinoside, EGCG, and methoxysalicylic acid had a higher content in the LN and HN groups. Thirteen alkaloids were detected (Figure 4C and Table S2). In the HN group, only choline, phosphocholine, guanine, and N-Lactitolamine were upregulated, whereas the contents of caffeine, guanine, and xanthine increased in the LN group. Additionally, 9 organic acids, 9 lipids, 10 aromatic substances, and 12 other substances were detected (Table S2). The contents of eight aromatic substances increased in the high nitrogen group, and piperidine-2,6-dicarboxylic acid, isocitrate acid, quinic acid, 2−(3,4,5−Trihydroxyphenyl) −3,4,5,7−tetrahydro−2H−1−benzopyran, and dihydroactinidiolide were upregulated in the roots from the HN group, while other components were not significantly different. In summary, the contents of most AAs increased as the N level increased, while the overall tea polyphenols decreased. Additionally, the contents of alkaloids and lipids decreased and the contents of aromatic substances increased in roots from the HN group.
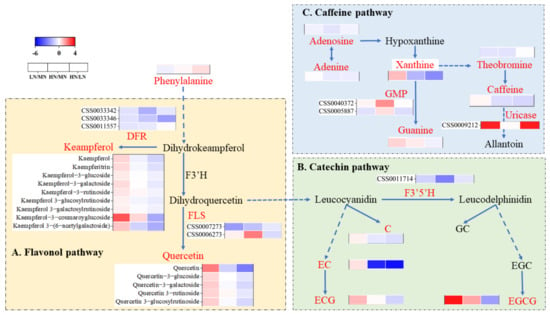
Figure 4.
Changes in secondary metabolites in roots of ‘Ziyan’ under different nitrogen conditions. (A): Flavonol pathway (yellow box); (B): catechin pathway (light-green box); (C): caffeine pathway (light-blue box).
3.3. Differentially Expressed Genes of the Roots under Different Nitrogen Levels
3.3.1. Analysis of Differentially Expressed Genes
High-throughput sequencing (RNA-seq) was conducted to clarify the molecular mechanism of ‘Ziyan’ root growth under the different N levels. The cDNA libraries of nine samples (three nitrogen levels and three biological repeats) were sequenced. As shown in Figure 5A–C, 635 DEGs were identified in the HN group (MNR vs. HNR; 230 upregulated and 405 downregulated). A total of 222 DEGs (110 upregulated and 112 downregulated) were discovered in the LN group (MNR vs. LNR). In summary, the MNR vs. HNR comparison of ‘Ziyan’ roots had more DEGs and more downregulated genes.
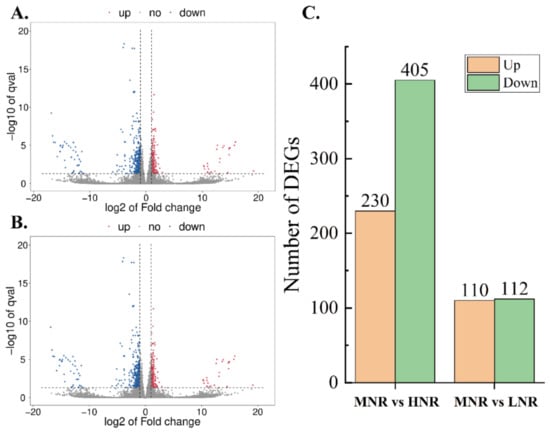
Figure 5.
Volcano map of expression genes (EDGs) ((A): MNR vs. HNR, (B): MNR vs. LNR) and number of EDGs (C).
3.3.2. Kyoto Encyclopedia of Genes and Genome Enrichment Analysis
A KEGG enrichment analysis was performed to better understand the functions of the DEGs. All DEGs in the MNR vs. HNR group and the MNR vs. LNR group were matched in the KEGG pathway database. Most DEGs in the MNR vs. HNR group were enriched in phytohormone signaling pathways, pentose and glucuronide interconversion, flavone biosynthesis, and monoterpene biosynthesis (Figure 6A). Twenty-eight pathways were included by the DEGs in the MNR vs. LNR group. The main pathways were phenylpropane synthesis, ABC translocator, isoflavone biosynthesis, and N metabolism (Figure 6B). Overall, the ‘Ziyan’ tea tree roots responded to N through phytohormone signaling, translocator, sugar metabolism, and nitrogen metabolism.
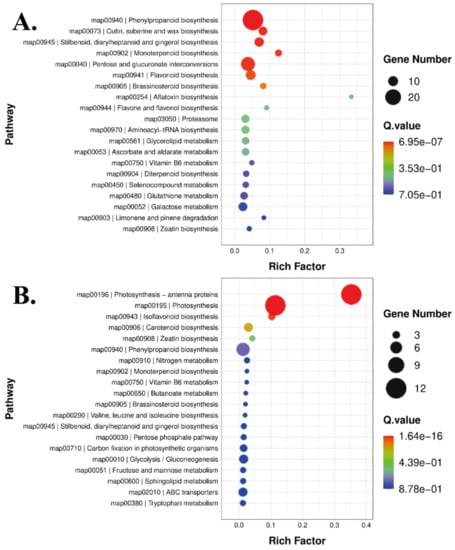
Figure 6.
KEGG enrichment analysis of differentially expressed genes ((A): MNR vs. HNR, (B): MNR vs. LNR).
3.3.3. Differentially Expressed Genes Analysis of Key Genes
Functional annotation of the DEGs showed that the TF, phytohormone, translocator, amino acid, and sugar metabolic genes related to root growth were differentially expressed in the LN and HN groups. Among the TFs (Figure 7A; Table S3), 43 DEGs were identified, of which 38 were concentrated in the ZY_HN_R vs. ZY_MN_R group. These DEGs were focused on the synthesis and signaling of 11 AP2/ERF, 7 MYB, 7 WRKY, 3 NAC, 3 bHLH, and 3 GATA. Compared with the MN group, AP2/ERF and GATA were upregulated in the HN group and most TFs were downregulated (e.g., MYB, WRKY, NAC) in roots. The HN group exhibited five DEGs for phytohormones (Figure 7B; Table S3), wherein the auxin-inducing protein (CSS0019669) and translocator (CSS0016609, CSS0030569) were upregulated, and cytokinin (CSS0009881) and gibberellin (CSS0019623) synthetase were downregulated. Additionally, cytokinin, ethylene, and abscisic acid-related genes were upregulated in the LN group (LNR/MNR). The HN group exhibited five amino acid translocators (Figure 7B; Table S3), wherein CSS0039117, CSS0021482, and CSS0001080 were downregulated. The HN group indicated upregulated the expression of nitrate translocators (CSS0005731, CSS0025443, CSS0027204) and downregulated the expression of sucrose (CSS0017942) and lipid (CSS0017942) translocators. NRT2.7 (CSS0001748) was upregulated in the LN group. Additionally, the HN group exhibited two amino acid metabolic enzymes, two sucrose, and four amylases (Figure 7B; Table S3). Herein, the HN group exhibited downregulation of GS (CSS0013068) and sucrose phosphate synthase (SPS) (CSS0009603) and upregulation of methionine γ-lyase (CSS0012078), β-amylase (CSS0043627, CSS0003801), and raffinose synthase 1 (CSS0001300).
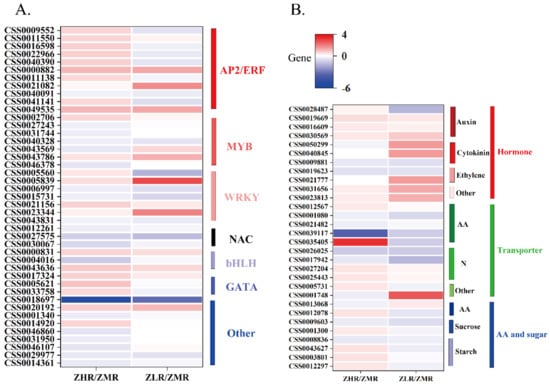
Figure 7.
Expression patterns of related genes under nitrogen treatments. (A): EDGs of transcription factor. (B): EDGs of hormone, transporter, AA (amino acid), and sugar metabolism. The scale represents the relative expression level of genes under low and medium nitrogen, high and medium nitrogen conditions. Expression differences of genes are represented by different colors ranging from low (blue) to high (red); the same below.
3.3.4. Verification of the Gene Expression Patterns by qRT-PCR
To further confirm the accuracy and reliability of the gene expression patterns in the transcriptome data, nine genes were selected, including two TFs (NAC and WRKY), three phytohormones (auxin transporter, GA2ox, and ERF), three amino acid synthetases (GOGAT, AQP, and MGL) and transport proteins, and one SPS. All results were verified by qRT-PCR. The results demonstrated that all the selected genes had similar regulatory patterns to those in the RNA-Seq data (Figure S1), demonstrating that the transcriptome data were reliable.
4. Discussion
4.1. Effect of High Nitrogen Supply on Amino Acids and Secondary Metabolites in ‘Ziyan’ Roots
AAs, polyphenols, and caffeine are three quality-related components in tea leaves. Herein, the differences in the metabolome were mapped to the KEGG pathway and were mainly enriched in the synthesis of AAs, polyphenols, and alkaloids. The differences in the genes of the enzymes and metabolites involved in their metabolic pathways were investigated. Theanine is mainly synthesized in roots and is transported to tea leaves through the vascular system [30,31]. Yang et al. [8] reported that Glu is the most abundant AA in the roots and the most active AA in response to N. Improving the N conditions helped increase the contents of The, Gln, Arg, Thr, Ser, and total AAs in the tea plant roots. In this study, the changes in the AAs induced by the high N supply were mainly involved in aspartate family and glutamate family metabolism, as well as histidine and serine metabolism (Figure 3). Interestingly, the expression of Phe, which is a major precursor of polyphenols, did not increase significantly in the HN group compared with the MN group of roots (Figure 3), suggesting that the high nitrogen supply only maintained the biosynthesis of Phe, thus enhancing the synthesis of Glu and aspartate. GS/GOGAT enzyme activity decreased under the high nitrogen supply, resulting in the greatest increase in Arg and suggesting that more N increased the Arg content through the urea cycle. Phenylalanine is a polyphenol precursor in tea, which synthesizes flavonoids and flavonoid glycosides through naringin, white anthocyanin, anthocyanin, and catechin (Figure 4A,B). In this study, the contents of nine kaempferols and their glycosides, and five quercetins and their glycosides, as well as C, EC, and ECG, decreased in the HN group; and the downregulation of dihydroflavonol-4-reductase (DFR), flavonoid synthase II (FLS), and flavonoid 3′,5′-hydroxylase (F3′5′H) expression (Figure 4A,B) supported this conclusion. Additionally, the EGCG content in roots from the HN group increased, which was different from previous studies [14,32]. As no significant increase in phenylalanine was detected in the roots under the high N treatment, the excess carbon resources were used to synthesize major AAs, which reduced carbon availability and inhibited the synthesis of polyphenolic substances. Moreover, the caffeine metabolic pathway consists of caffeine synthesis and caffeine degradation. The synthesis consists of adenosine converted to xanthine, theobromine, and finally to caffeine. In our study, adenosine, xanthine, adenine, theobromine, and caffeine decreased under the high nitrogen treatment, particularly xanthine and caffeine, while GMP and uricase accelerated the breakdown of these two substances. The caffeine content in the HN group decreased, possibly because the synthesis of AAs consumes excess carbon resources [33].
Most MYB, WRKY, and NAC TFs regulate the biosynthesis of catechins, anthocyanins, and caffeine. For example, CsMYB5b upregulates LAR and ANR in tea plant roots to increase the synthesis of catechins and proanthocyanidins [34,35]. MYB184 positively regulates TCS1 by increasing caffeine content in roots [36]. The upregulation of MdWRKY1 expression promotes the transcription of anthocyanin-related genes and the accumulation of anthocyanin [37]. CsNAC7 positively regulates the caffeine synthase gene yhNMT1 and promotes caffeine accumulation in tea plants [38]. In this study, most MYB, WRKY, and NAC TFs were downregulated under the high nitrogen supply (Figure 7A; Table S3), suggesting that the high nitrogen supply suppressed the synthesis of catechins, anthocyanins, and caffeine. Additionally, high N supplies more carbon skeletons for AA synthesis, which inhibits the synthesis and accumulation of sugars, lipids, and flavonoids in leaves [22,32,39,40].
In summary, the high nitrogen supply significantly enhanced the synthesis of AAs, while excess nitrogen intake inhibited carbon metabolism through the tricarboxylic acid cycle, leading to a decrease in sucrose, polyphenols, and caffeine. The balance between carbon and nitrogen metabolism in the tea plant was disrupted, which was detrimental to growth.
4.2. Morphological and Physiological Traits of ‘Ziyan’ Roots under Different Nitrogen Levels
High N cultivation is common in China’s tea plantations, as supplying N significantly increases the yield of tea. Nevertheless, applying excess N reduces the quality of tea leaves and causes ecological and environmental problems. China has promoted a policy of reducing N fertilizer and improving the efficiency of N fertilizer use in crops. In this case, measures to reduce N fertilizer applications have been focused on improving the N use efficiency in tea plants.
Phytohormones are influenced by N and are involved in the regulation of root growth [41]. Auxin is mainly synthesized at the stem tip, moving through the stem vascular tissue to the root tip, and through the peripheral root tissue to the base and the elongation zone [42]. Root apical auxin flux is mainly controlled by auxin efflux carriers, including PIN and LAX [43], and is negatively regulated by early auxin response genes (e.g., GH3, Aux/IAA, and SAUR). Additionally, cytokinin promotes the synthesis of auxin in roots and regulates the translocation of auxin [44]. It has been demonstrated that a low nitrogen supply increases the auxin content in roots and promotes root growth, whereas a high nitrogen supply achieves the opposite [45]. In this study, the indole acetic acid (IAA) content in roots from the HN group decreased after downregulating the Aux/IAA, PIN, and LAX genes, which suppressed the synthesis of cytokinin (cytokinin hydroxylase downregulated), thus, inhibiting root growth. In addition, the low nitrogen conditions promoted growth hormone (GH3.5 down-regulated) synthesis, thereby reducing auxin content in roots to promote root growth [41]. Auxin regulates lateral root formation by stimulating other endogenous phytohormones (e.g., cytokinin, gibberellin, abscisic acid, and ethylene) and environmental signals [46].
TFs regulate root growth in response to biotic and abiotic stressors, where MYB, AP2/ERF, GATA, and other TFs are involved in nitrogen stress [8,12,36]. The upregulation of MYB in plant roots enhances the expression of NRT, NAR, NIR, and GS, whereas the downregulation of MYB73/MYB77 directly inhibits the transcription of auxin-responsive genes, thereby suppressing lateral root development [47,48]. AP2/ERF-mediated gibberellin is involved in coordinating long-chain fatty acid biosynthesis during lateral root development, and overexpressing ERF13 inhibits lateral root growth and development [49]. In contrast, overexpressing PeGATA26 significantly inhibits primary root length and plant height [50]. In this study, AP2/ERF and GATA were upregulated and MYB was downregulated in the HN group, suggesting that a high nitrogen supply suppressed root growth (Figure 7A; Table S3).
The N/AA translocator and amino acid metabolic enzymes, which are related to root growth, are involved in the response to N stress through N signaling and affect root growth [51]. The NRT1/PTR family, also known as NPF, is involved in the absorption and transport of nitrate [52]. The overexpression of OsNRT1.1 in rice under a high nitrogen supply upregulates nitrate and ammonium transport-related genes and increases nitrogen absorption [53,54]. It has been demonstrated that AAP6 enhances N stress tolerance and source-to-sink transport and promotes the root absorption and translocation of AAs [55,56]. CsAAP1 expression at the roots is highly correlated with the translocation of Thea from the roots to the buds [57]. Raffinose synthase enhances drought tolerance in leaves by using sucrose to synthesize cottonseed sugars [58]. In this study, NRT1/PTR was mainly upregulated, while the AA transport genes (e.g., AVT 3, ProT 1, AAP 6) and saccharide translocator (SUT 3) were downregulated in the HN group (Figure 7B; Table S3). Glutamate synthetase (GOGAT 1) and SPS synthetic expression were downregulated, while methionine gamma lyase (MGL), β-amylase (starch hydrolase), and raffinose synthase (RAFS) were upregulated in the HN group. This result demonstrates that a high nitrogen supply enhances root nitrogen uptake and transport but inhibits sugar and lipid transport, affecting the balance of root carbon and N metabolism and inhibiting normal root growth [57,59,60]. A high N supply suppresses the translocation and biosynthesis of AAs in roots, which may result from a feedback mechanism of high AA content in the roots. The decrease in the sucrose content in the roots may have been attributed to the fact that the high nitrogen supply limited root carbon resources. Future studies should focus on how a high nitrogen supply affects AAs and secondary metabolites in roots.
In tea plants, there was an obvious genotype × N rate interaction effect on root architecture growth, as some tea genotypes were highly responsive to the N application while others were not [61]. Further, the structure of the root system directly affects the ability of the plant to obtain N. In our study, ‘Ziyan’ roots exhibited increased AA contents and significantly reduced the activity of carbon and N metabolic enzymes under high N conditions, resulting in a significant decrease in root length, volume, total root number, and root activity, and inhibiting root growth [62]. Ruan’s group compared root growth in two contrasting tea genotypes: Longjing43 (low-N tolerant) and Liyou002 (high-N tolerant), and they also found that high-N tolerant Liyou002 produced fewer lateral roots than Longjing43 [63]. Additionally, changes in ‘Ziyan’ root N metabolic enzymes (GS, GOGAT, and GDH) support this view. The content of N and AAs in the ‘Ziyan’ roots decreased significantly under the low N condition, but root surface area, volume, total root number, and root activity increased significantly. These results demonstrate that ‘Ziyan’ adopted a plant “foraging strategy”, by allocating more carbon to promote root growth to obtain more N. Therefore, applying moderate N can increase the activity of carbon and N metabolic enzymes in ‘Ziyan’ roots and significantly increase N and AA contents in the root system to promote root growth. A low level of N fertilization is recommended to manage ‘Ziyan’ tea plantations and promote the root growth of tea seedlings in a young tea plantation, while a combination of ‘nitrogen reduction and efficiency’ fertilization techniques is preferable in a production tea plantation.
5. Conclusions
In this study, global variations in the transcriptome and expression patterns of AAs, polyphenols, and caffeine-synthetic-related genes in response to different N levels were investigated in ‘Ziyan’ roots. The results demonstrated that a high nitrogen supply favored the synthesis of AAs in ‘Ziyan’ roots and inhibited the synthesis of polyphenols and caffeine. ‘Ziyan’ root growth was inhibited under the high nitrogen supply, which may be attributed to the cooperation of the entire regulatory network, including the overall regulatory mechanisms of carbon and N metabolism in tea plants, as well as the synergistic effects of various TFs, translocators, and phytohormones. This study will help to elucidate the regulatory mechanisms of root N in tea plants and provide useful information for guiding fertilization techniques in ‘Ziyan’ tea.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy13040968/s1, Figure S1: RT-qPCR validation of some genes under nitrogen treatment; Table S1: Primers used for qRT-PCR analysis; Table S2: Metabolite changes in root of ‘Ziyan’ under different nitrogen treatments; Table S3: Expression patterns of related genes under nitrogen treatments.
Author Contributions
W.X. and W.C. performed the experiments and data analyses. D.T. and Y.Y. contributed to plant management and phenotype data collection. X.T. participated in chart design and data analysis. W.X. and D.T. wrote the paper. Q.T. and L.T. coordinated the project. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Department of Science and Technology of Sichuan Province (2021YFN0004, 2022NSFSC0180) and the Opening Fund of Tea Refining and Innovation Key Laboratory of Sichuan Province (SCTOF202203).
Data Availability Statement
Data are contained within supplementary material.
Acknowledgments
Thank you for the research facilities provided by the public platform of the Tea Refining and Innovation Key Laboratory of Sichuan Province.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chandran, A.K.N.; Priatama, R.A.; Kumar, V.; Xuan, Y.; Je, B.I.; Kim, C.M.; Jung, K.-H.; Han, C.-D. Genome-wide transcriptome analysis of expression in rice seedling roots in response to supplemental nitrogen. J. Plant Physiol. 2016, 200, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Ruan, L.; Wei, K.; Wang, L.; Cheng, H.; Zhang, F.; Wu, L.; Bai, P.; Zhang, C. Characteristics of NH4+ and NO3− fluxes in tea (Camellia sinensis) roots measured by scanning ion-selective electrode technique. Sci. Rep. 2016, 6, 38370. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Y.; Cheng, Y.-H.; Chen, K.-E.; Tsay, Y.-F. Nitrate Transport, Signaling, and Use Efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-S.; Zhong, Q.-S.; Lin, Z.-H.; Yu, W.-Q.; Wang, M.-K.; Chen, Z.-H.; You, X.-M. Screening tea varieties for nitrogen efficiency. J. Plant Nutr. 2017, 40, 1797–1804. [Google Scholar] [CrossRef]
- Peng, Z.; Liu, Y.; Li, Y.; Abawi, Y.; Wang, Y.; Men, M.; An-Vo, D.-A. Responses of Nitrogen Utilization and Apparent Nitrogen Loss to Different Control Measures in the Wheat and Maize Rotation System. Front. Plant Sci. 2017, 8, 160. [Google Scholar] [CrossRef]
- Tayefeh, M.; Sadeghi, S.M.; Noorhosseini, S.A.; Bacenetti, J.; Damalas, C.A. Environmental impact of rice production based on nitrogen fertilizer use. Environ. Sci. Pollut. Res. 2018, 25, 15885–15895. [Google Scholar] [CrossRef]
- Hu, S.; Zhang, M.; Yang, Y.; Xuan, W.; Zou, Z.; Arkorful, E.; Chen, Y.; Ma, Q.; Jeyaraj, A.; Chen, X.; et al. A novel insight into nitrogen and auxin signaling in lateral root formation in tea plant [Camellia sinensis (L.) O. Kuntze]. BMC Plant Biol. 2020, 20, 232. [Google Scholar] [CrossRef]
- Yang, T.; Li, H.; Tai, Y.; Dong, C.; Cheng, X.; Xia, E.; Chen, Z.; Li, F.; Wan, X.; Zhang, Z. Transcriptional regulation of amino acid metabolism in response to nitrogen deficiency and nitrogen forms in tea plant root (Camellia sinensis L.). Sci. Rep. 2020, 10, 6868. [Google Scholar] [CrossRef]
- Liu, X.; Hu, B.; Chu, C. Nitrogen assimilation in plants: Current status and future prospects. J. Genet. Genom. 2022, 49, 394–404. [Google Scholar] [CrossRef]
- Sun, H.; Tao, J.; Liu, S.; Huang, S.; Chen, S.; Xie, X.; Yoneyama, K.; Zhang, Y.; Xu, G. Strigolactones are involved in phosphate- and nitrate-deficiency-induced root development and auxin transport in rice. J. Exp. Bot. 2014, 65, 6735–6746. [Google Scholar] [CrossRef]
- Gao, K.; Chen, F.; Yuan, L.; Zhang, F.; Mi, G. A comprehensive analysis of root morphological changes and nitrogen allocation in maize in response to low nitrogen stress. Plant Cell Environ. 2015, 38, 740–750. [Google Scholar] [CrossRef]
- Gaudinier, A.; Rodriguez-Medina, J.; Zhang, L.; Olson, A.; Liseron-Monfils, C.; Bågman, A.-M.; Foret, J.; Abbitt, S.; Tang, M.; Li, B.; et al. Transcriptional regulation of nitrogen-associated metabolism and growth. Nature 2018, 563, 259–264. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, J.; Li, H.; Shi, W.; Polle, A.; Lu, M.; Sun, X.; Luo, Z.-B. Global poplar root and leaf transcriptomes reveal links between growth and stress responses under nitrogen starvation and excess. Tree Physiol. 2015, 35, 1283–1302. [Google Scholar] [CrossRef]
- Xu, W.; Li, J.; Zhang, L.; Zhang, X.; Zhao, H.; Guo, F.; Wang, Y.; Wang, P.; Chen, Y.; Ni, D.; et al. Metabolome and RNA-seq Analysis of Responses to Nitrogen Deprivation and Resupply in Tea Plant (Camellia sinensis) Roots. Front. Plant Sci. 2022, 13, 932720. [Google Scholar] [CrossRef]
- Lin, Z.-H.; Chen, C.-S.; Zhong, Q.-S.; Ruan, Q.-C.; Chen, Z.-H.; You, X.-M.; Shan, R.-Y.; Li, X.-L. The GC-TOF/MS-based Metabolomic analysis reveals altered metabolic profiles in nitrogen-deficient leaves and roots of tea plants (Camellia sinensis). BMC Plant Biol. 2021, 21, 506. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, H.; Pilon-Smits, E.; Huang, W.; Wang, P.; Wang, M.; Guo, F.; Wang, Y.; Li, R.; Zhao, H.; et al. Transcriptome-Wide Analysis of Nitrogen-Regulated Genes in Tea Plant (Camellia sinensis L. O. Kuntze) and Characterization of Amino Acid Transporter CsCAT9.1. Plants 2020, 9, 1218. [Google Scholar] [CrossRef]
- Huang, H.; Yao, Q.; Xia, E.; Gao, L. Metabolomics and Transcriptomics Analyses Reveal Nitrogen Influences on the Accumulation of Flavonoids and Amino Acids in Young Shoots of Tea Plant (Camellia sinensis L.) Associated with Tea Flavor. J. Agric. Food Chem. 2018, 66, 9828–9838. [Google Scholar] [CrossRef]
- Ruan, L.; Wei, K.; Li, J.; He, M.; Wu, L.; Aktar, S.; Wang, L.; Cheng, H. Responses of tea plants (Camellia sinensis) with different low-nitrogen tolerances during recovery from nitrogen deficiency. J. Sci. Food Agric. 2022, 102, 1405–1414. [Google Scholar] [CrossRef]
- Tan, L.; Yang, C.; Zhou, B.; Wang, L.; Zou, Y.; Chen, W.; Xia, T.; Tang, Q. Inheritance and quantitative trait loci analyses of the anthocyanins and catechins of Camellia sinensis cultivar ‘Ziyan’ with dark-purple leaves. Physiol. Plant. 2020, 170, 109–119. [Google Scholar] [CrossRef]
- Li, W.; Tan, L.; Zou, Y.; Tan, X.; Huang, J.; Chen, W.; Tang, Q. The Effects of Ultraviolet A/B Treatments on Anthocyanin Accumulation and Gene Expression in Dark-Purple Tea Cultivar ‘Ziyan’ (Camellia sinensis). Molecules 2020, 25, 354. [Google Scholar] [CrossRef]
- Liu, Z.-W.; Li, H.; Liu, J.-X.; Wang, Y.; Zhuang, J. Integrative transcriptome, proteome, and microRNA analysis reveals the effects of nitrogen sufficiency and deficiency conditions on theanine metabolism in the tea plant (Camellia sinensis). Hortic. Res. 2020, 7, 65. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Haerdter, R.; Gerendás, J. Impact of nitrogen supply on carbon/nitrogen allocation: A case study on amino acids and catechins in green tea [Camellia sinensis (L.) O. Kuntze] plants. J. Plant Biol. 2010, 12, 724–734. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.; Zhou, D.; Song, J.; Gao, J. Nitrogen Uptake and Distribution in Different Chinese Cabbage Genotypes under Low Nitrogen Stress. Int. J. Mol. Sci. 2022, 23, 1573. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Li, L.; Meng, Y.; Liu, H.; Li, J.; Yu, Y.; Qian, C.; Wei, S.; Gu, W. Exogenous Spermidine Optimizes Nitrogen Metabolism and Improves Maize Yield under Drought Stress Conditions. Agriculture 2022, 12, 1270. [Google Scholar] [CrossRef]
- Yu, X.; Li, Y.; He, C.; Zhou, J.; Chen, Y.; Yu, Z.; Wang, P.; Ni, D. Nonvolatile metabolism in postharvest tea (Camellia sinensis L.) leaves: Effects of different withering treatments on nonvolatile metabolites, gene expression levels, and enzyme activity. Food Chem. 2020, 327, 126992. [Google Scholar] [CrossRef]
- Li, P.; Dai, W.; Lu, M.; Xie, D.; Tan, J.; Yang, C.; Zhu, Y.; Lv, H.; Peng, Q.; Zhang, Y.; et al. Metabolomic analysis reveals the composition differences in 13 Chinese tea cultivars of different manufacturing suitabilities. J. Sci. Food Agric. 2018, 98, 1153–1161. [Google Scholar] [CrossRef]
- Chen, D.; Sun, Z.; Gao, J.; Peng, J.; Wang, Z.; Zhao, Y.; Lin, Z.; Dai, W. Metabolomics combined with proteomics provides a novel interpretation of the compound differences among Chinese tea cultivars (Camellia sinensis var. sinensis) with different manufacturing suitabilities. Food Chem. 2021, 377, 131976. [Google Scholar] [CrossRef]
- Sun, Z.; Chen, D.; Zhu, L.; Zhao, Y.; Lin, Z.; Li, X.; Dai, W. A comprehensive study of the differences in protein expression and chemical constituents in tea leaves (Camellia sinensis var. sinensis) with different maturity using a combined proteomics and metabolomics method. Food Res. Int. 2022, 157, 111397. [Google Scholar] [CrossRef]
- Wei, C.; Yang, H.; Wang, S.; Zhao, J.; Liu, C.; Gao, L.; Xia, E.; Lu, Y.; Tai, Y.; She, G.; et al. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA 2018, 115, E4151–E4158. [Google Scholar] [CrossRef]
- Dong, F.; Hu, J.; Shi, Y.; Liu, M.; Zhang, Q.; Ruan, J. Effects of nitrogen supply on flavonol glycoside biosynthesis and accumulation in tea leaves (Camellia sinensis). Plant Physiol. Biochem. 2019, 138, 48–57. [Google Scholar] [CrossRef]
- Li, W.; Xiang, F.; Zhong, M.; Zhou, L.; Liu, H.; Li, S.; Wang, X. Transcriptome and metabolite analysis identifies nitrogen utilization genes in tea plant (Camellia sinensis). Sci. Rep. 2017, 7, 1693. [Google Scholar] [CrossRef]
- Liu, J.; Liu, M.; Fang, H.; Zhang, Q.; Ruan, J. Accumulation of Amino Acids and Flavonoids in Young Tea Shoots Is Highly Correlated With Carbon and Nitrogen Metabolism in Roots and Mature Leaves. Front. Plant Sci. 2021, 12, 756433. [Google Scholar] [CrossRef]
- Su, H.; Zhang, X.; He, Y.; Li, L.; Wang, Y.; Hong, G.; Xu, P. Transcriptomic Analysis Reveals the Molecular Adaptation of Three Major Secondary Metabolic Pathways to Multiple Macronutrient Starvation in Tea (Camellia sinensis). Genes 2020, 11, 241. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, L.; Jiang, X.; Dai, X.; Xu, L.; Li, T.; Xing, D.; Li, Y.; Li, M.; Gao, L.; et al. Evolutionary and functional characterization of leucoanthocyanidin reductases from Camellia sinensis. Planta 2017, 247, 139–154. [Google Scholar] [CrossRef]
- Wang, P.; Liu, Y.; Zhang, L.; Wang, W.; Hou, H.; Zhao, Y.; Jiang, X.; Yu, J.; Tan, H.; Wang, Y.; et al. Functional demonstration of plant flavonoid carbocations proposed to be involved in the biosynthesis of proanthocyanidins. Plant J. 2020, 101, 18–36. [Google Scholar] [CrossRef]
- Li, P.; Ye, Z.; Fu, J.; Xu, Y.; Shen, Y.; Zhang, Y.; Tang, D.; Li, P.; Zuo, H.; Tong, W.; et al. CsMYB184 regulates caffeine biosynthesis in tea plants. Plant Biotechnol. J. 2022, 20, 1012–1014. [Google Scholar] [CrossRef]
- Ma, H.; Yang, T.; Li, Y.; Zhang, J.; Wu, T.; Song, T.; Yao, Y.; Tian, J. The long noncoding RNA MdLNC499 bridges MdWRKY1 and MdERF109 function to regulate early-stage light-induced anthocyanin accumulation in apple fruit. Plant Cell 2021, 33, 3309–3330. [Google Scholar] [CrossRef]
- Ma, W.; Kang, X.; Liu, P.; She, K.; Zhang, Y.; Lin, X.; Li, B.; Chen, Z. The NAC-like transcription factor CsNAC7 positively regulates the caffeine biosynthesis-related gene yhNMT1 in Camellia sinensis. Hortic. Res. 2022, 9, uhab046. [Google Scholar] [CrossRef]
- Yang, Y.Y.; Li, X.H.; Ratcliffe, R.G.; Ruan, J.Y. Characterization of ammonium and nitrate uptake and assimilation in roots of tea plants. Russ. J. Plant Physiol. 2013, 60, 91–99. [Google Scholar] [CrossRef]
- Luo, D.; Gao, Y.; Wang, Y.; Shi, Y.; Chen, S.; Ding, Z.; Fan, K. Using UAV image data to monitor the effects of different nitrogen application rates on tea quality. J. Sci. Food Agric. 2021, 102, 1540–1549. [Google Scholar] [CrossRef]
- Lv, X.; Zhang, Y.; Hu, L.; Zhang, Y.; Zhang, B.; Xia, H.; Du, W.; Fan, S.; Kong, L. Low-Nitrogen Stress Stimulates Lateral Root Initiation and Nitrogen Assimilation in Wheat: Roles of Phytohormone Signaling. J. Plant Growth Regul. 2020, 40, 436–450. [Google Scholar] [CrossRef]
- Xie, X.; Wang, Y.; Datla, R.; Ren, M. Auxin and Target of Rapamycin Spatiotemporally Regulate Root Organogenesis. Int. J. Mol. Sci. 2021, 22, 11357. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Chen, H.; Wang, P.; Chen, F.; Yuan, L.; Mi, G. Low nitrogen induces root elongation via auxin-induced acid growth and auxin-regulated target of rapamycin (TOR) pathway in maize. J. Plant Physiol. 2020, 254, 153281. [Google Scholar] [CrossRef] [PubMed]
- Jing, H.; Strader, L.C. Interplay of Auxin and Cytokinin in Lateral Root Development. Int. J. Mol. Sci. 2019, 20, 486. [Google Scholar] [CrossRef]
- Jia, Z.; Giehl, R.F.; von Wirén, N. Nutrient–hormone relations: Driving root plasticity in plants. Mol. Plant 2022, 15, 86–103. [Google Scholar] [CrossRef]
- Xiao, G.; Zhang, Y. Adaptive Growth: Shaping Auxin-Mediated Root System Architecture. Trends Plant Sci. 2020, 25, 121–123. [Google Scholar] [CrossRef]
- Ma, N.; Dong, L.; Lü, W.; Lü, J.; Meng, Q.; Liu, P. Transcriptome analysis of maize seedling roots in response to nitrogen-, phosphorus-, and potassium deficiency. Plant Soil 2020, 447, 637–658. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, L.; Chen, P.; Liang, T.; Li, X.; Liu, H. UV-B photoreceptor UVR8 interacts with MYB73/MYB77 to regulate auxin responses and lateral root development. EMBO J. 2020, 39, e101928. [Google Scholar] [CrossRef]
- Guyomarc’H, S.; Boutté, Y.; Laplaze, L. AP2/ERF transcription factors orchestrate very long chain fatty acid biosynthesis during Arabidopsis lateral root development. Mol. Plant 2021, 14, 205–207. [Google Scholar] [CrossRef]
- Wang, T.; Yang, Y.; Lou, S.; Wei, W.; Zhao, Z.; Ren, Y.; Lin, C.; Ma, L. Genome-Wide Characterization and Gene Expression Analyses of GATA Transcription Factors in Moso Bamboo (Phyllostachys edulis). Int. J. Mol. Sci. 2019, 21, 14. [Google Scholar] [CrossRef]
- Kong, L.; Zhang, Y.; Du, W.; Xia, H.; Fan, S.; Zhang, B. Signaling Responses to N Starvation: Focusing on Wheat and Filling the Putative Gaps With Findings Obtained in Other Plants. A Review. Front. Plant Sci. 2021, 12, 656696. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xia, X.; Zeng, Y.; Nong, B.; Zhang, Z.; Wu, Y.; Tian, Q.; Zeng, W.; Gao, J.; Zhou, W.; et al. Genome-wide identification of the peptide transporter family in rice and analysis of the PTR expression modulation in two near-isogenic lines with different nitrogen use efficiency. BMC Plant Biol. 2020, 20, 193. [Google Scholar] [CrossRef]
- Fan, X.; Feng, H.; Tan, Y.; Xu, Y.; Miao, Q.; Xu, G. A putative 6-transmembrane nitrate transporter OsNRT1.1b plays a key role in rice under low nitrogen. J. Integr. Plant Biol. 2016, 58, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hu, B.; Yuan, D.; Liu, Y.; Che, R.; Hu, Y.; Ou, S.; Liu, Y.; Zhang, Z.; Wang, H.; et al. Expression of the Nitrate Transporter Gene OsNRT1.1A/OsNPF6.3 Confers High Yield and Early Maturation in Rice. Plant Cell 2018, 30, 638–651. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, D.; Mei, Y.; Xia, T.; Xu, W.; Zhang, Y.; You, X.; Zhang, X.; Li, L.; Wang, N.N. Overexpression of GmAAP6a enhances tolerance to low nitrogen and improves seed nitrogen status by optimizing amino acid partitioning in soybean. Plant Biotechnol. J. 2020, 18, 1749–1762. [Google Scholar] [CrossRef]
- Peng, B.; Kong, H.; Li, Y.; Wang, L.; Zhong, M.; Sun, L.; Gao, G.; Zhang, Q.; Luo, L.; Wang, G.; et al. OsAAP6 functions as an important regulator of grain protein content and nutritional quality in rice. Nat. Commun. 2014, 5, 4847. [Google Scholar] [CrossRef]
- Dong, C.; Li, F.; Yang, T.; Feng, L.; Zhang, S.; Li, F.; Li, W.; Xu, G.; Bao, S.; Wan, X.; et al. Theanine transporters identified in tea plants (Camellia sinensis L.). Plant J. 2020, 101, 57–70. [Google Scholar] [CrossRef]
- Li, T.; Zhang, Y.; Liu, Y.; Li, X.; Hao, G.; Han, Q.; Dirk, L.M.A.; Downie, A.B.; Ruan, Y.-L.; Wang, J.; et al. Raffinose synthase enhances drought tolerance through raffinose synthesis or galactinol hydrolysis in maize and Arabidopsis plants. J. Biol. Chem. 2020, 295, 8064–8077. [Google Scholar] [CrossRef]
- Gebauer, P.; Korn, M.; Engelsdorf, T.; Sonnewald, U.; Koch, C.; Voll, L.M. Sugar Accumulation in Leaves of Arabidopsis sweet11/sweet12 Double Mutants Enhances Priming of the Salicylic Acid-Mediated Defense Response. Front. Plant Sci. 2017, 8, 1378. [Google Scholar] [CrossRef]
- Xu, Y.; Ren, Y.; Li, J.; Li, L.; Chen, S.; Wang, Z.; Xin, Z.; Chen, F.; Lin, T.; Cui, D.; et al. Comparative Proteomic Analysis Provides New Insights Into Low Nitrogen-Promoted Primary Root Growth in Hexaploid Wheat. Front. Plant Sci. 2019, 10, 151. [Google Scholar] [CrossRef]
- Jagadish, R.; Shanmugaselvan, V.A. Influence of nitrogen and potassium on root nutrient and root CEC of different tea cultivars. (Camellia sinensis, C. assamica and C.assamica spp. Lasiocalyx). Rhizosphere 2016, 1, 36–44. [Google Scholar]
- Chen, J.; Liu, S.; Zhang, S.; Ge, C.; Shen, Q.; Ma, H.; Zhang, X.; Dong, H.; Zhao, X.; Pang, C. Nitrogen modulates cotton root morphology by affecting abscisic acid (ABA) and salicylic acid (SA) content. Arch. Agron. Soil Sci. 2020, 67, 1722–1738. [Google Scholar] [CrossRef]
- Ruan, L.; Wang, L.; Wei, K.; Cheng, H.; Li, H.; Shao, S.; Wu, L. Comparative analysis of nitrogen spatial heterogeneity responses in low nitrogen susceptible and tolerant tea plants (Camellia sinensis L.). Sci. Hortic. 2019, 246, 182–189. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).