
Different Temperature Regimes Influenced the Quality of Broccoli Seedlings, Which Caused a Change in the Chemical Composition of Mature Heads

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Field Experiments
2.3. Morphological Parameters of Broccoli Seedlings
2.4. Fresh Weight of Broccoli Seedlings
2.5. Dry Weight of Broccoli Seedlings and Mature Heads
2.6. Soluble Sugars and L-ascorbic Acid Determination of Broccoli Seedlings and Mature Heads
2.7. Photosynthetic Pigments Content of Mature Broccoli Heads
2.8. Mineral Composition of Mature Broccoli Heads
2.9. Statistical Analysis
3. Results
3.1. Morphological Parameters of Broccoli Seedlings
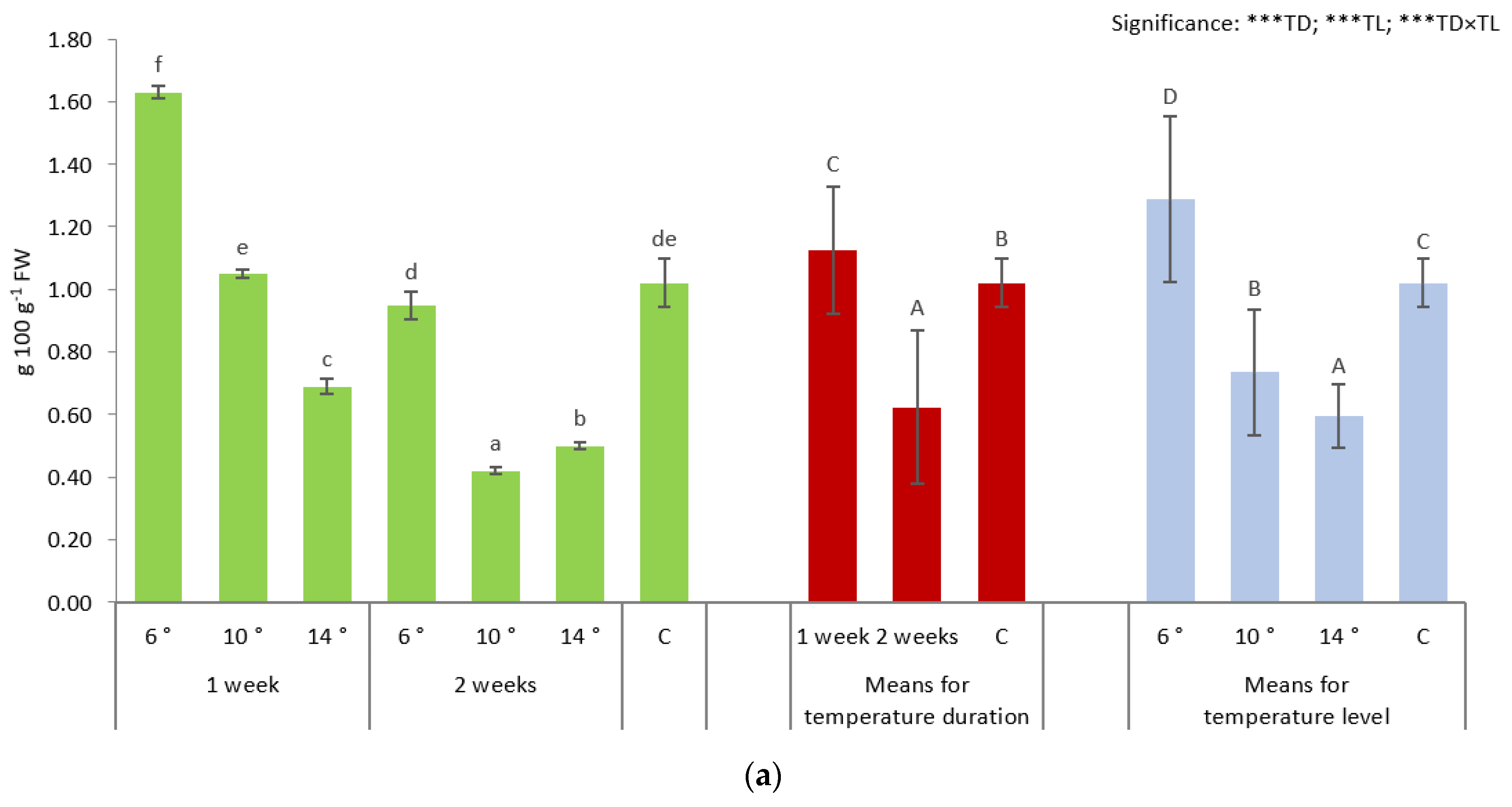
3.2. Fresh and Dry Weight of Broccoli Seedlings
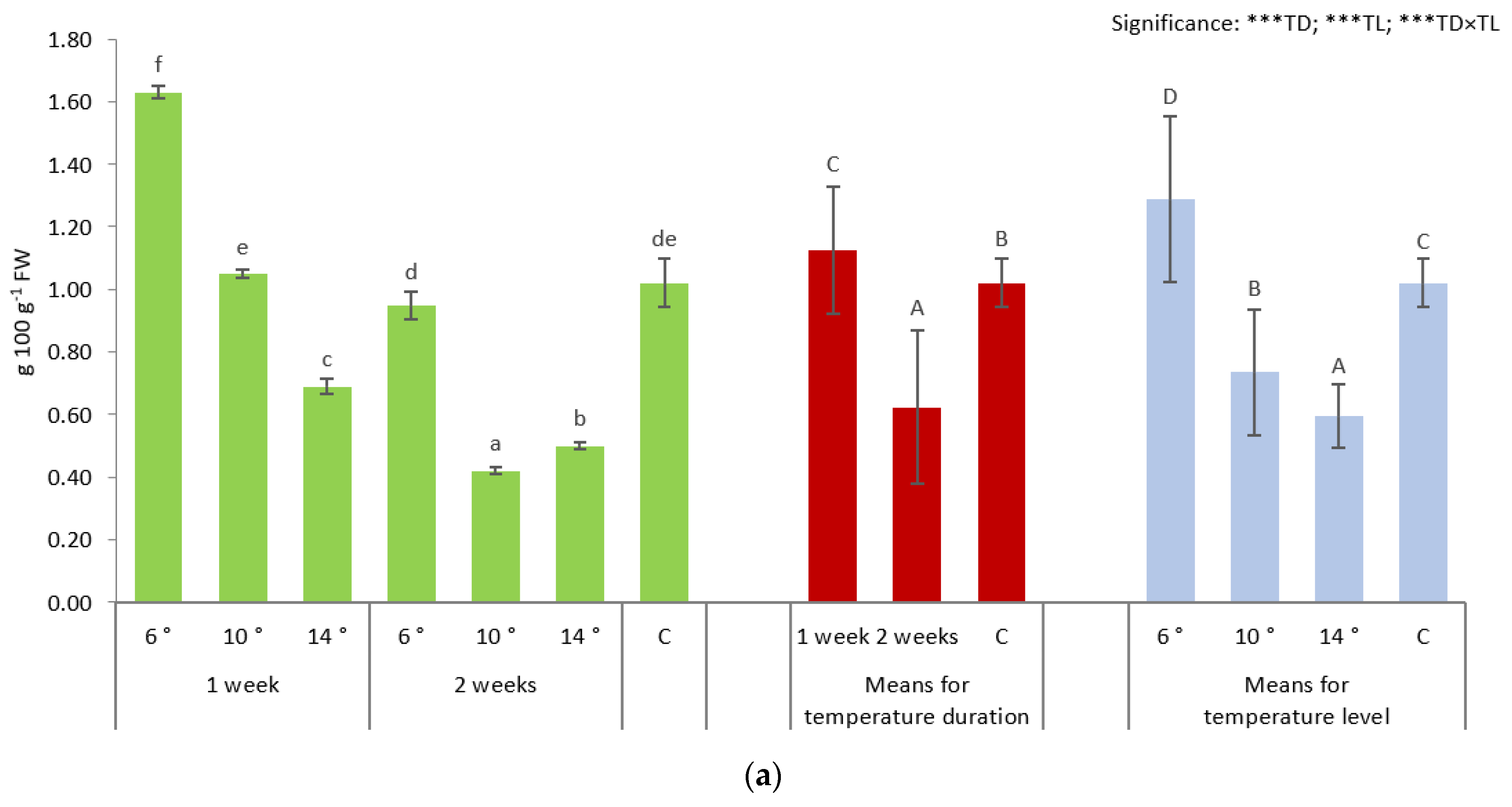
3.3. Soluble Sugars and L-Ascorbic Acid of Broccoli Seedlings and Mature Heads
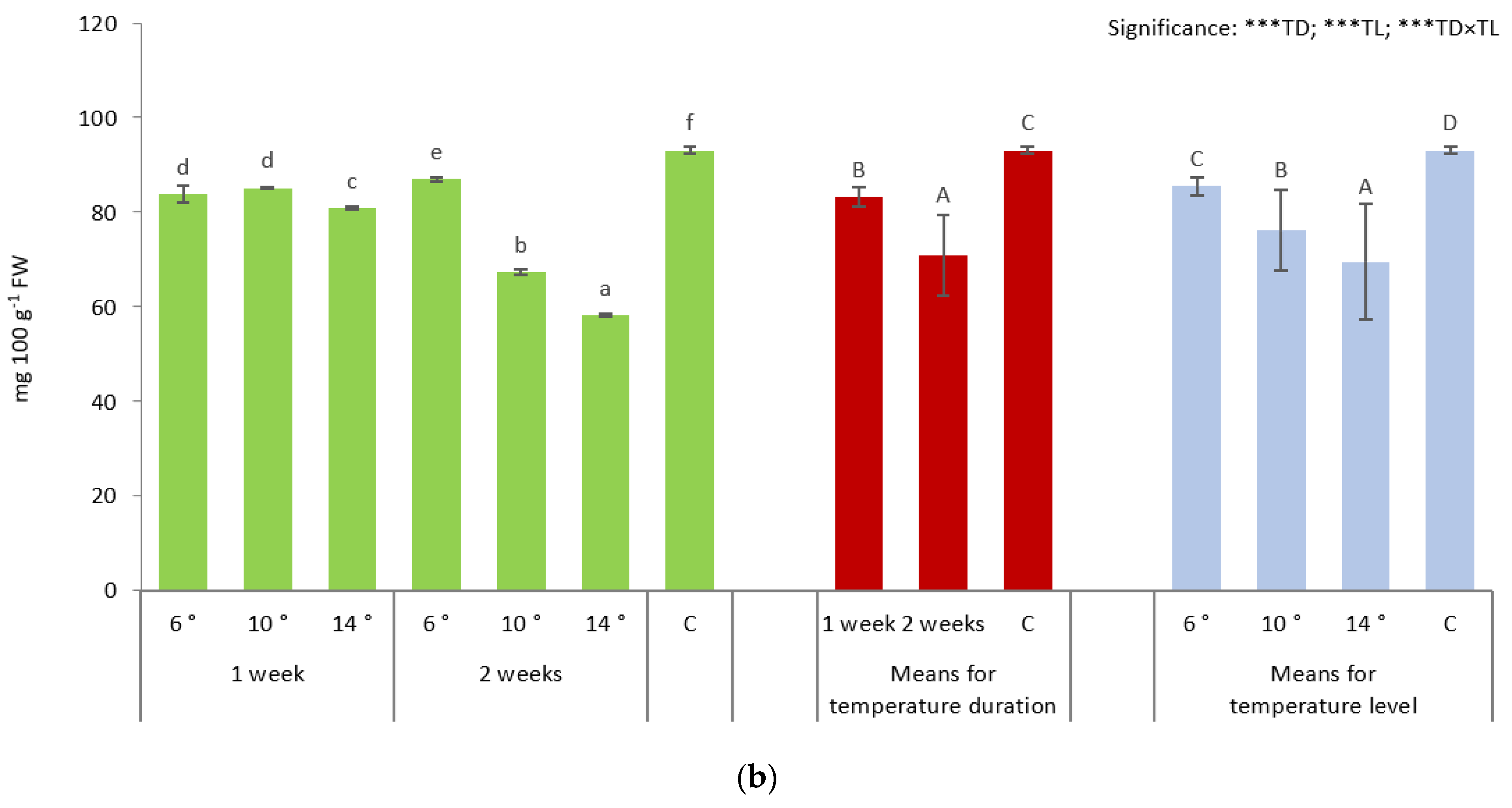
3.4. Dry Weight and Photosynthetic Pigments Content of Fully Mature Broccoli Heads
3.5. Mineral Composition of Fully Mature Broccoli Heads
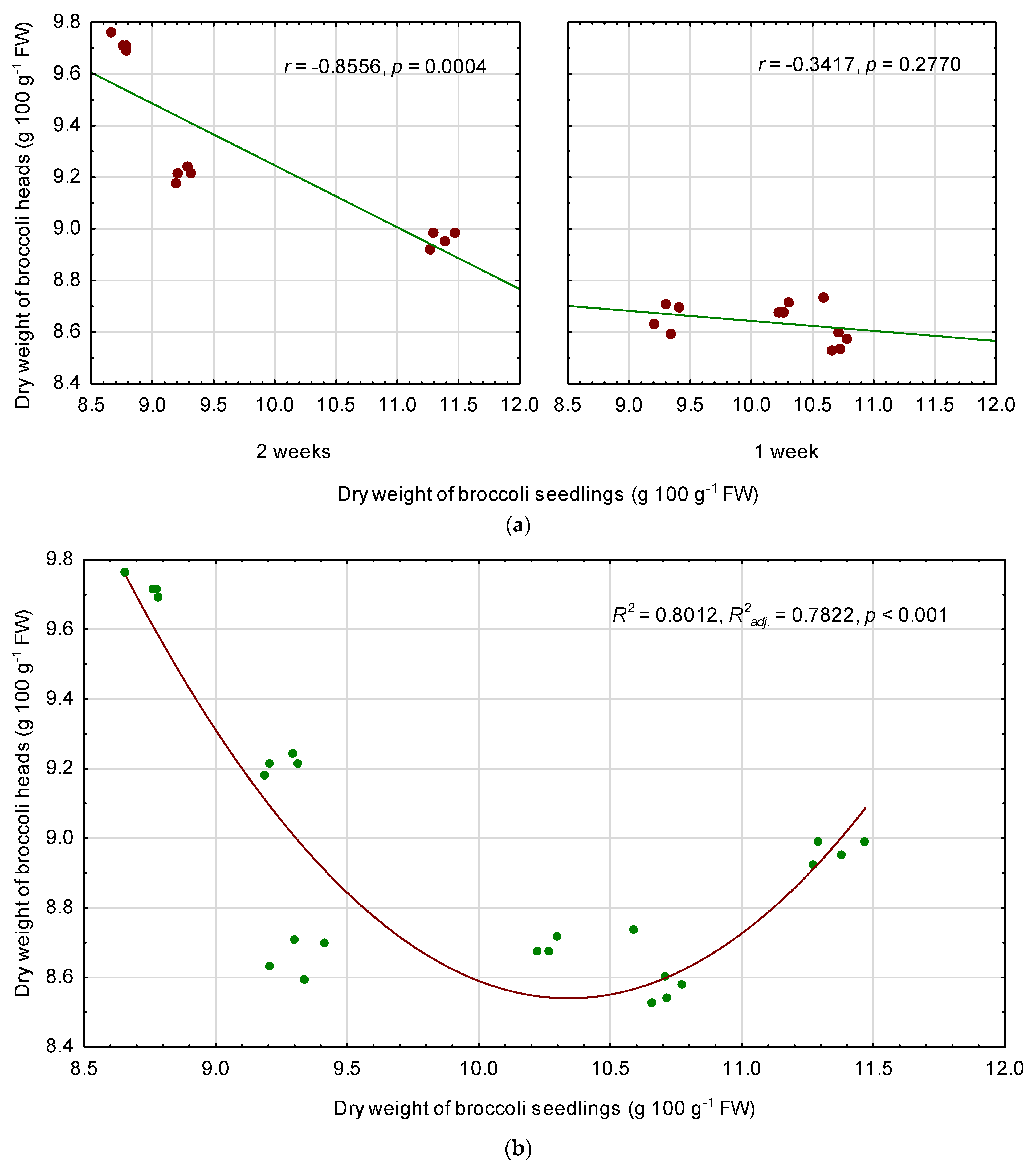
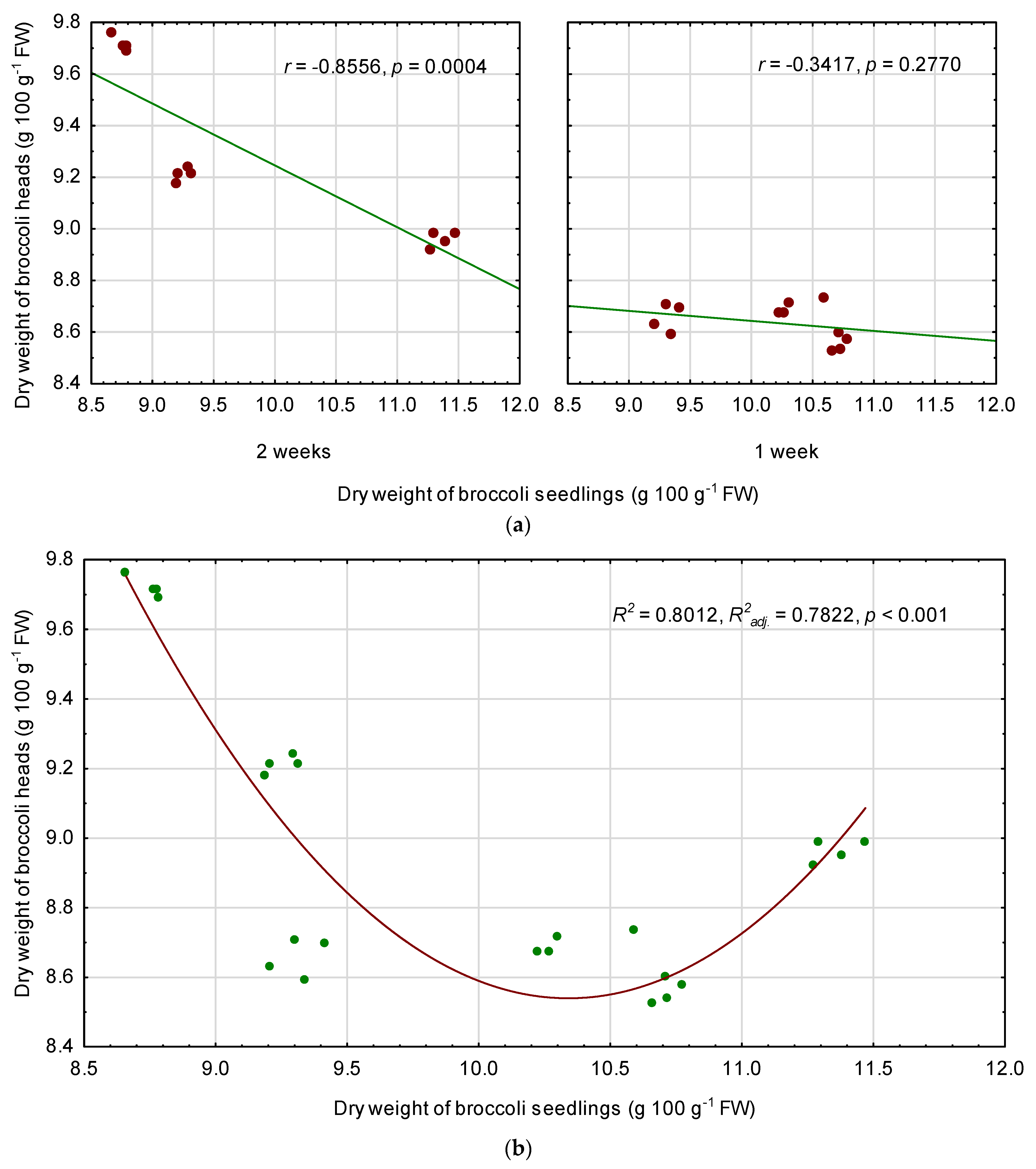
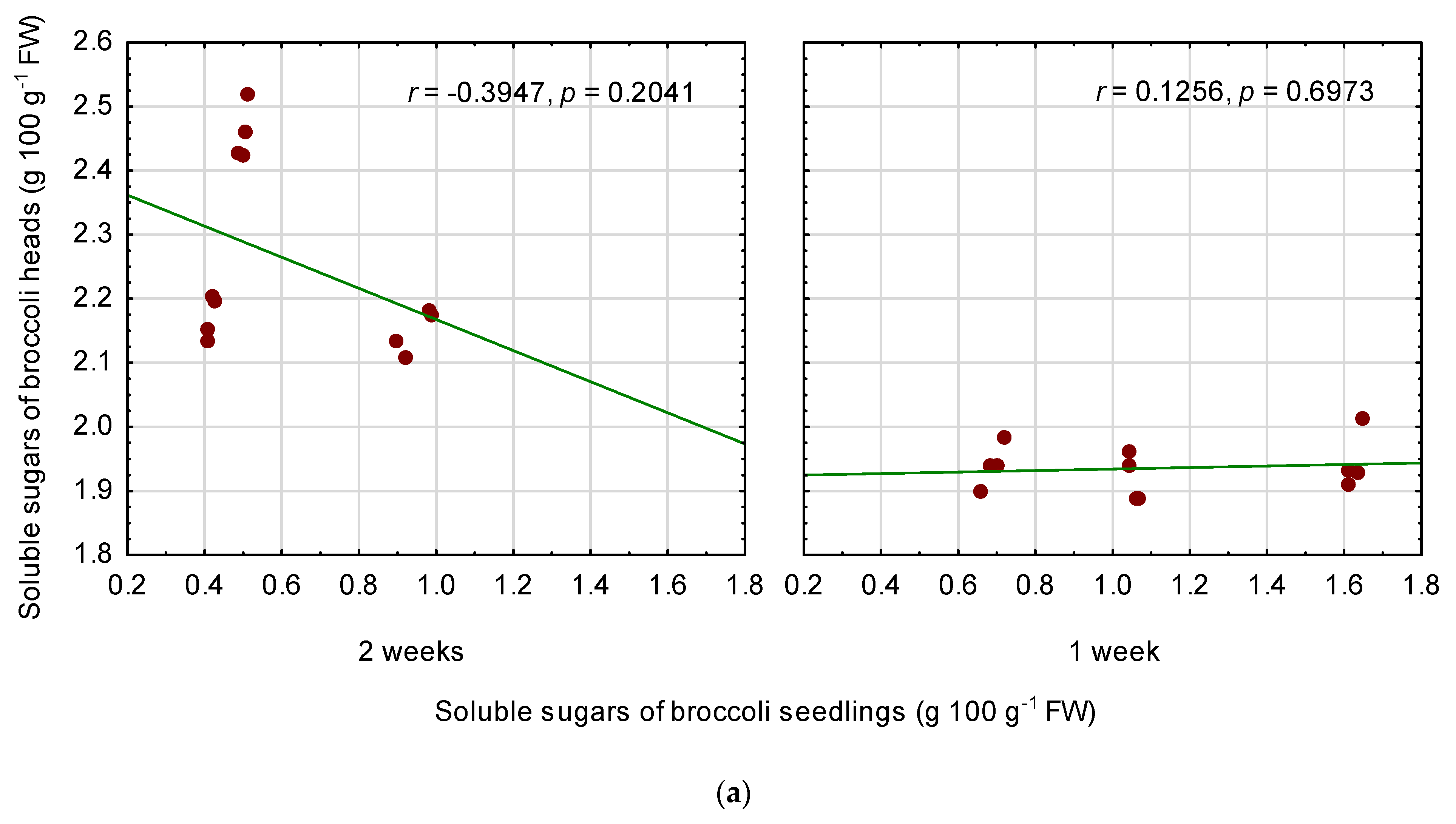
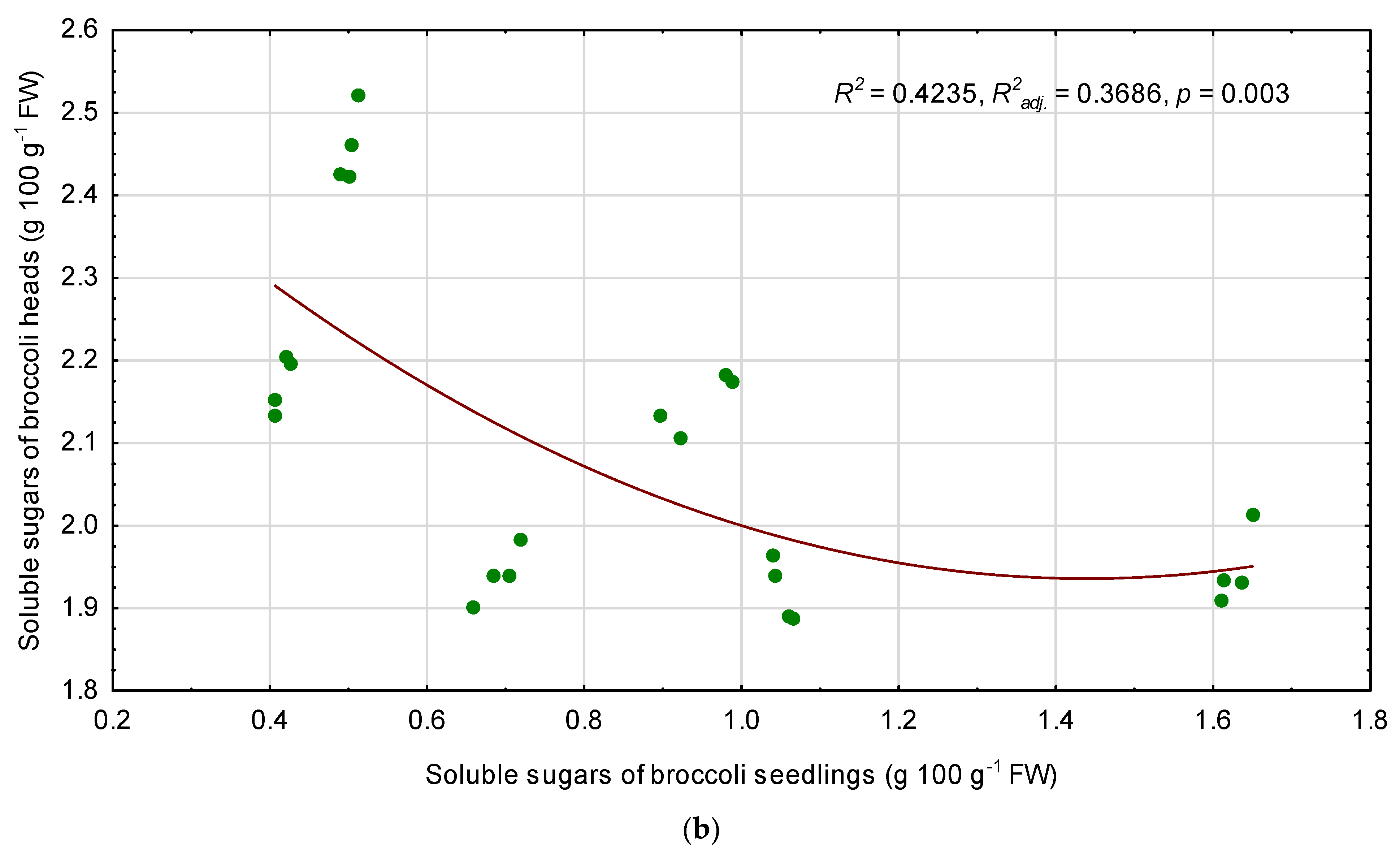
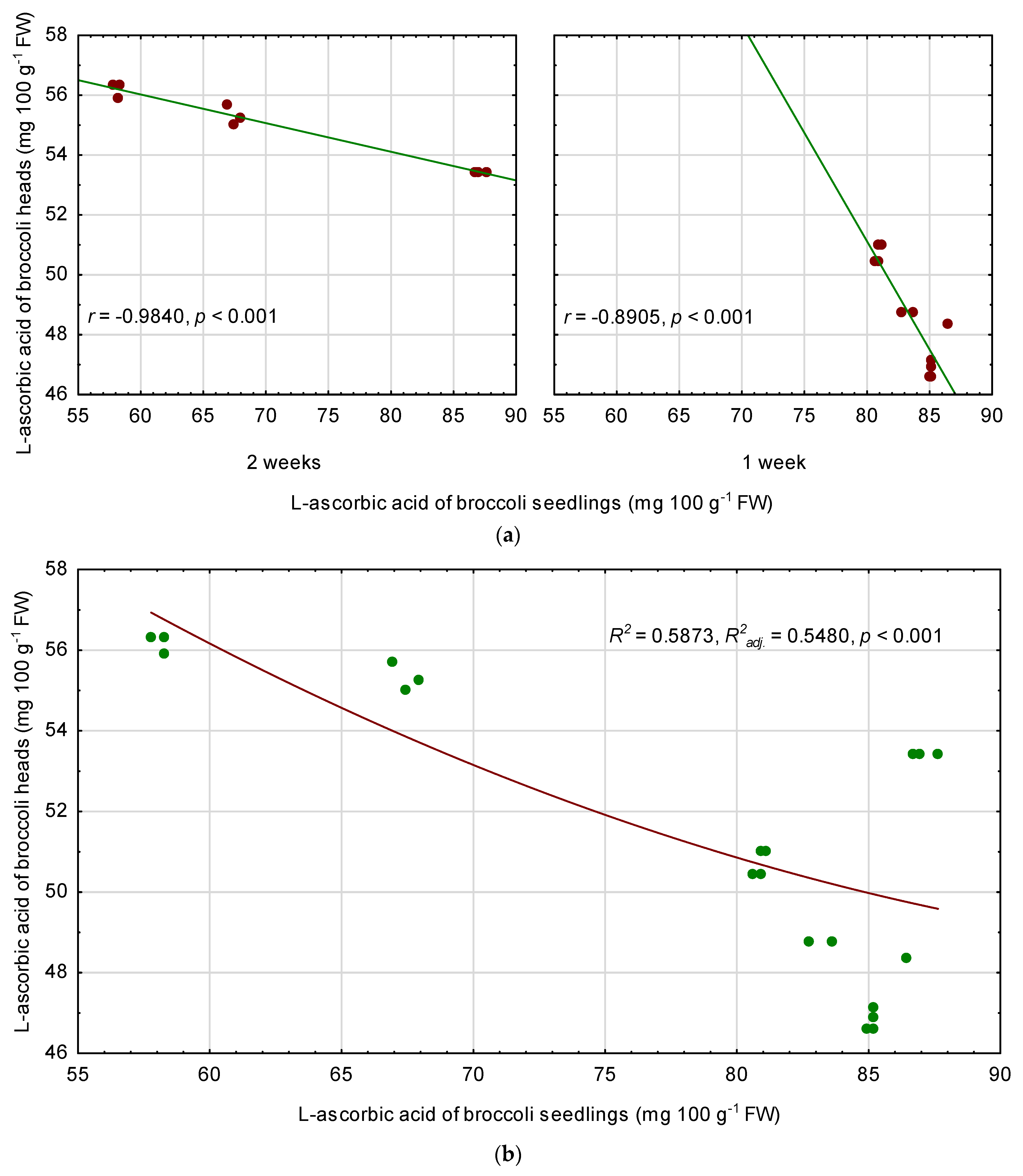
3.6. Relationships among Quality Parameters of Seedlings and Fully Mature Broccoli Heads
4. Discussion
4.1. Broccoli Seedlings Morphological Traits and Chemical Composition as Affected by Chilling
4.2. Broccoli Heads Chemical Composition as Affected by Seedlings Chilling
4.3. Relationships between the Seedling Traits and the Chemical Composition of Fully Mature Broccoli Heads
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moreno, D.A.; López-Berenguer, C.; Carvajal, M.; García-Viguera, C. Health benefits of broccoli. Influence of pre- and post-harvest factors on bioactive compounds. Food 2007, 1, 297–312. [Google Scholar]
- Podsędek, A. Natural antioxidants and antioxidant capacity of Brassica vegetables: A review. LWT Food Sci. Technol. 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Valverde, J.; Reilly, K.; Villacreces, S.; Gaffney, M.; Grant, J.; Brunton, N. Variation in bioactive content in broccoli (Brassica oleracea var. italica) grown under conventional and organic production systems. J. Sci. Food Agric. 2015, 95, 1163–1171. [Google Scholar] [CrossRef]
- Pathare, P.B.; Mohapatra, D. Bioactive compounds in broccoli: Extraction and processing. In Vegetable Processing and Bioactive Compounds; Kadam, D.M., Sharma, M., Kaur, D., Eds.; Studium Press (India) Pvt. Ltd.: New Delhi, India, 2017; pp. 420–450. [Google Scholar]
- Nuñez-Gómez, V.; Baenas, N.; Navarro-González, I.; García-Alonso, J.; Moreno, D.A.; González-Barrio, R.; Periago-Castón, M.J. Seasonal variation of health-promoting bioactives in broccoli and methyl-jasmonate pre-harvest treatments to enhance their contents. Foods 2020, 9, 1371. [Google Scholar] [CrossRef]
- Jahangir, M.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Health-affecting compounds in Brassicaceae. Compr. Rev. Food Sci. Food Saf. 2009, 8, 31–43. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Wang, W.S.; Zhang, F.; Zhang, T.; Zhao, W.; Fu, B.-Y.; Li, Z.-K. Temporal profiling of primary metabolites under chilling stress and its association with seedling chilling tolerance of rice (Oryza sativa L.). Rice 2013, 6, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Ullah, F.; Zhou, D.-X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Rivero, R.M.; Ruiz, J.M.; García, P.C.; López-Lefebre, L.R.; Sánchez, E.; Romero, R. Resistance to cold and heat stress: Accumulation of phenolic compounds in tomato and watermelon plants. Plant Sci. 2001, 160, 315–321. [Google Scholar] [CrossRef]
- Sivaci, A.; Kaya, A.; Duman, S. Effects of ascorbic acid on some physiological changes of pepino (Solanum muricatum Ait.) under chilling stress. Acta Biol. Hung. 2014, 65, 305–318. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, H.; Ichimura, K.; Imada, S.; Hamaki, S. Sucrose synthase and sucrosephosphate synthase, but not acid invertase, are regulated by cold acclimation and deacclimation in cabbage seedlings. J. Plant Physiol. 2001, 158, 847–852. [Google Scholar] [CrossRef]
- Gao, Q.H.; Xu, K.; Wang, X.F.; Wu, Y. Effect of grafting on cold tolerance in eggplant seedlings. Acta Hortic. 2008, 771, 167–174. [Google Scholar] [CrossRef]
- Długosz-Grochowska, O.; Leja, M.; Grabowska, A.; Kunicki, E. The effect of preliminary chilling of broccoli transplants on some antioxidative parameters. Folia Hort. 2012, 24, 131–139. [Google Scholar]
- Javanmardi, J.; Rahem, M.; Nasirzadeh, M. Physiological and reproductive responses of tomato and pepper transplants to low-temperature conditioning. Int. J. Veg. Sci. 2013, 19, 294–310. [Google Scholar] [CrossRef]
- Kalisz, A.; Sękara, A.; Smoleń, S.; Grabowska, A.; Gil, J.; Cebula, S. Mineral composition of cauliflowers with differently coloured curds modified by the chilling of juvenile plants. Sci. Hortic. 2018, 232, 216–225. [Google Scholar] [CrossRef]
- Kalisz, A.; Sękara, A.; Smoleń, S.; Grabowska, A.; Gil, J.; Komorowska, M.; Kunicki, E. Survey of 17 elements, including rare earth elements, in chilled and non-chilled cauliflower cultivars. Sci. Rep. 2019, 9, 5416. [Google Scholar] [CrossRef]
- FAO. World Reference Base for Soil Resources; FAO: Rome, Italy, 2014. [Google Scholar]
- Grabowska, A.; Kunicki, E. Brokuły od Wiosny do Jesieni [Broccoli from Spring to Fall]; Planpress: Kraków, Poland, 2013. [Google Scholar]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krełowska-Kułas, M. Badanie jakości produktów spożywczych. In The Study of Food Quality; PWN: Warszawa, Poland, 1993. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 603, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Ostrowska, A.; Gawliński, S.; Szczubiałka, Z. Metody Analizy i Oceny Właściwości Gleb i Roślin [The Methods of Analysis and Estimation of Soils and Plants Properties]; Instytut Ochrony Środowiska: Warszawa, Poland, 1991. [Google Scholar]
- Ruelland, E.; Vaultier, M.N.; Zachowski, A.; Hurry, V. Cold signalling and cold acclimation in plants. Adv. Bot. Res. 2009, 49, 35–150. [Google Scholar]
- Hussain, S.; Khan, F.; Cao, W.; Wu, L.; Geng, M. Seed priming alters the production and detoxification of reactive oxygen intermediates in rice seedlings grown under sub-optimal temperature and nutrient supply. Front. Plant Sci. 2016, 7, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalisz, A.; Cebula, S. The effect of temperature on growth and chemical composition of Chinese cabbage seedlings in spring period. Folia Hort. 2006, 18, 3–15. [Google Scholar]
- Rodríguez, V.M.; Soengas, P.; Alonso-Villaverde, V.; Sotelo, T.; Cartea, M.E.; Velasco, P. Effect of temperature stress on the early vegetative development of Brassica oleracea L. BMC Plant Biol. 2015, 15, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verheul, M.J.; Picatto, C.; Stamp, P. Growth and development of maize (Zea mays L.) seedlings under chilling conditions in the field. Eur. J. Agron. 1996, 5, 31–43. [Google Scholar] [CrossRef]
- Poiré, R.; Schneider, H.; Thorpe, M.R.; Kuhn, A.J.; Schurr, U.; Walter, A. Root cooling strongly affects diel leaf growth dynamics, water and carbohydrate relations in Ricinus communis. Plant Cell Environ. 2010, 33, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K. Cold stress tolerance mechanisms in plants. A review. Agron. Sust. Dev. 2010, 30, 515–527. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Malik, J.; Thakur, P.; Kaistha, S.; Sharma, D.; Upadhyaya, H.D.; Berger, J.D.; Nayyar, H. Growth and metabolic responses of contrasting chickpea (Cicer arietinum) genotypes to chilling stress at reproductive phase. Acta Physiol. Plant. 2011, 33, 779–787. [Google Scholar] [CrossRef]
- Nievola, C.C.; Carvalho, C.P.; Carvalho, V.; Rodrigues, E. Rapid responses of plants to temperature changes. Temperature 2017, 4, 371–405. [Google Scholar] [CrossRef] [PubMed]
- Crisp, P.A.; Ganguly, D.; Eichten, S.R.; Borevitz, J.O.; Pogson, B.J. Reconsidering plant memory: Intersections between stress recovery, RNA turnover, and epigenetics. Sci. Adv. 2016, 2, e1501340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, T.J.A.; Matthes, M.C.; Napier, J.A.; Pickett, J.A. Stressful “memories” of plants: Evidence and possible mechanisms. Plant Sci. 2007, 173, 603–608. [Google Scholar] [CrossRef]
- Lämke, J.; Bäurle, I. Epigenetic and chromatin-based mechanisms in environmental stress adaptation and stress memory in plants. Genome Biol. 2017, 18, 124. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.F.; Peng, X.F.; Han, X.; Ren, J.; Zhan, K.Y.; Zhu, M. The stress factor, exogenous ascorbic acid, affects plant growth and the antioxidant system in Arabidopsis thaliana. Russ. J. Plant Physiol. 2014, 61, 467–475. [Google Scholar] [CrossRef]
- Venkatesh, J.; Park, S.W. Role of L-ascorbate in alleviating abiotic stresses in crop plants. Bot. Stud. 2014, 55, 38. [Google Scholar] [CrossRef] [Green Version]
- Duan, M.; Ma, N.N.; Li, D.; Deng, Y.S.; Kong, F.Y.; Lv, W.; Meng, Q.W. Antisense-mediated sup-pression of tomato thylakoidal ascorbate peroxidase influences antioxidant network during chilling stress. Plant Physiol. Biochem. 2012, 58, 37–45. [Google Scholar] [CrossRef]
- Sękara, A.; Kalisz, A.; Bączek-Kwinta, R.; Gawęda, M.; Pohl, A.; Grabowska, A. The effect of abiotic stresses applied in the juvenile phase of eggplant ontogeny on chemical composition of seedlings and fruits. Agrochimica 2015, 59, 26–43. [Google Scholar]
- Kalisz, A.; Sękara, A.; Grabowska, A.; Cebula, S.; Kunicki, E. The effect of chilling stress at transplant stage on broccoli development and yield with elements of modeling. J. Plant Growth Regul. 2015, 34, 532–544. [Google Scholar] [CrossRef]
- Taşpınar, M.S.; Dumlupinar, R.; Demir, F.; Çakmak, T.; Gülen, Y. Determination of chilling temperature effects on nutrient elements composition and distribution in cole (Brassica oleracea L. cv. acephala) using the WDXRF spectroscopic technique. Afr. J. Biotechnol. 2009, 8, 6569–6575. [Google Scholar]
- Hussain, H.A.; Men, S.; Hussain, S.; Zhang, Q.; Ashraf, U.; Anjum, S.A.; Ali, I.; Wang, L. Maize tolerance against drought and chilling stresses varied with root morphology and antioxidative defense system. Plants 2020, 9, 720. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2011. [Google Scholar]
- Zengin, F.K. The effect of Co2+ and Zn2+ on the contents of protein, abscisic acid, proline and chlorophyll in bean (Phaseolus vulgaris cv. Strike) seedlings. J. Environ. Biol. 2006, 27, 441–448. [Google Scholar] [PubMed]
- Waraich, E.A.; Ahmad, R.; Halim, A.; Aziz, T. Alleviation of temperature stress by nutrient management in crop plants: A review. J. Soil Sci. Plant Nutr. 2012, 12, 221–244. [Google Scholar] [CrossRef] [Green Version]
- Cebula, S. The effect of pot size and transplant age on the yield and quality of white, green and romanesco cauliflower curds. Veg. Crops Res. Bull. 2009, 70, 101–110. [Google Scholar] [CrossRef]
- Grabowska, A.; Sękara, A.; Kalisz, A.; Kunicki, E.; Wojciechowska, R.; Kopta, T. Optimisation of transplant age in combination with dark-chilling to enhance the biological quality of broccoli cultivated in summer. Not. Bot. Horti. Agrobot. Cluj Napoca 2018, 46, 494–500. [Google Scholar] [CrossRef] [Green Version]
- Kalisz, A.; Sękara, A.; Cebula, S.; Grabowska, A.; Kunicki, E. Impact of low-temperature transplant treatment on yield and quality of cauliflower curds in late spring production. Sci. Hortic. 2014, 176, 134–142. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature Duration (Weeks) | Temperature Level (°C) | Height (cm) | Number of Leaves | Leaf Area (cm2) | Leaf Perimeter (cm) | Leaf Width (cm) | Leaf Length (cm) |
|---|---|---|---|---|---|---|---|
| 1 | 6 | 18.5 ± 0.33 c | 4.3 ± 0.06 b | 29.8 ± 0.89 d | 39.5 ± 1.18 b | 5.8 ± 0.10 d | 6.9 ± 0.14 a |
| 10 | 18.3 ± 0.27 c | 4.6 ± 0.11 c | 27.5 ± 0.47 c | 39.7 ± 0.43 b | 5.5 ± 0.05 cd | 7.1 ± 0.13 ab | |
| 14 | 19.5 ± 0.24 d | 4.8 ± 0.06 c | 28.4 ± 0.77 cd | 40.2 ± 0.90 b | 5.6 ± 0.16 d | 7.2 ± 0.18 ab | |
| 2 | 6 | 16.2 ± 0.27 a | 3.7 ± 0.14 a | 21.8 ± 0.46 a | 34.2 ± 0.52 a | 4.9 ± 0.12 a | 7.0 ± 0.08 ab |
| 10 | 17.0 ± 0.20 b | 4.2 ± 0.11 b | 23.7 ± 0.90 b | 35.8 ± 1.43 a | 5.1 ± 0.07 ab | 7.1 ± 0.24 ab | |
| 14 | 19.2 ± 0.09 d | 4.6 ± 0.08 c | 27.2 ± 0.38 c | 39.0 ± 0.32 b | 5.3 ± 0.03 bc | 7.4 ± 0.09 b | |
| Control | 18 | 23.3 ± 0.19 e | 5.1 ± 0.20 d | 35.7 ± 1.43 e | 44.2 ± 1.24 c | 6.2 ± 0.16 e | 7.2 ± 0.20 ab |
| Duration (TD) (weeks) | |||||||
| 1 | 18.8 ± 0.60 B | 4.6 ± 0.20 B | 28.6 ± 1.17 B | 39.8 ± 0.90 B | 5.6 ± 0.14 B | 7.1 ± 0.21 A | |
| 2 | 17.5 ± 1.31 A | 4.2 ± 0.41 A | 24.2 ± 2.42 A | 36.3 ± 2.26 A | 5.1 ± 0.21 A | 7.2 ± 0.20 A | |
| Control | 23.3 ± 0.19 C | 5.1 ± 0.20 C | 35.7 ± 1.43 C | 44.2 ± 1.24 C | 6.2 ± 0.16 C | 7.2 ± 0.20 A | |
| Temperature (TL) (°C) | |||||||
| 6 | 17.4 ± 1.23 A | 4.0 ± 0.36 A | 25.8 ± 4.35 A | 36.9 ± 2.97 A | 5.4 ± 0.49 A | 7.0 ± 0.14 A | |
| 10 | 17.7 ± 0.75 B | 4.4 ± 0.25 B | 25.6 ± 2.16 A | 37.8 ± 2.11 A | 5.3 ± 0.27 A | 7.1 ± 0.18 AB | |
| 14 | 19.4 ± 0.25 C | 4.7 ± 0.10 C | 27.8 ± 1.00 B | 39.6 ± 0.87 B | 5.5 ± 0.18 A | 7.3 ± 0.15 B | |
| Control | 23.3 ± 0.19 D | 5.1 ± 0.20 D | 35.7 ± 1.43 C | 44.2 ± 1.24 C | 6.2 ± 0.16 B | 7.2 ± 0.20 AB | |
| Significance | |||||||
| TD | *** | *** | *** | *** | *** | ns | |
| TL | *** | *** | *** | *** | *** | ** | |
| TD × TL | *** | *** | *** | *** | *** | ** | |
| Temperature Duration (Weeks) | Temperature Level (°C) | Shoot Fresh Weight (g per Plant) | Shoot Dry Weight (g 100 g−1 FW) | Total Shoot Dry Weight (g per Plant) | Root Fresh Weight (g per Plant) | Root Dry Weight (g 100 g−1 FW) | Total Root Dry Weight (g per Plant) |
|---|---|---|---|---|---|---|---|
| 1 | 6 | 3.77 ± 0.268 b | 10.72 ± 0.046 d | 0.404 ± 0.0271 c | 0.38 ± 0.025 c | 12.63 ± 0.640 bc | 0.048 ± 0.0055 b |
| 10 | 3.66 ± 0.521 b | 10.35 ± 0.166 c | 0.379 ± 0.0513 bc | 0.47 ± 0.043 d | 9.77 ± 0.170 a | 0.046 ± 0.0044 b | |
| 14 | 3.85 ± 0.145 b | 9.32 ± 0.085 b | 0.359 ± 0.0127 bc | 0.48 ± 0.022 d | 12.14 ± 0.203 b | 0.058 ± 0.0030 c | |
| 2 | 6 | 2.51 ± 0.183 a | 11.36 ± 0.090 e | 0.285 ± 0.0213 a | 0.22 ± 0.007 a | 13.95 ± 0.869 cd | 0.031 ± 0.0023 a |
| 10 | 2.97 ± 0.100 a | 9.25 ± 0.061 b | 0.275 ± 0.0100 a | 0.19 ± 0.017 a | 15.07 ± 0.926 d | 0.029 ± 0.0029 a | |
| 14 | 3.71 ± 0.178 b | 8.75 ± 0.059 a | 0.325 ± 0.0169 ab | 0.30 ± 0.028 b | 11.54 ± 0.472 b | 0.035 ± 0.0030 a | |
| Control | 18 | 4.90 ± 0.038 c | 10.15 ± 0.083 c | 0.497 ± 0.0012 d | 0.50 ± 0.032 d | 12.48 ± 0.378 b | 0.062 ± 0.0048 c |
| Duration (TD) (weeks) | |||||||
| 1 | 3.76 ± 0.326 B | 10.13 ± 0.627 B | 0.381 ± 0.0366 B | 0.44 ± 0.056 B | 11.51 ± 1.355 A | 0.051 ± 0.0069 B | |
| 2 | 3.06 ± 0.537 A | 9.79 ± 1.181 A | 0.295 ± 0.0271 A | 0.24 ± 0.052 A | 13.52 ± 1.694 C | 0.032 ± 0.0036 A | |
| Control | 4.90 ± 0.038 C | 10.15 ± 0.083 B | 0.497 ± 0.0012 C | 0.50 ± 0.032 C | 12.48 ± 0.378 B | 0.062 ± 0.0048 C | |
| Temperature (TL) (°C) | |||||||
| 6 | 3.14 ± 0.708 A | 11.04 ± 0.348 D | 0.346 ± 0.0676 A | 0.30 ± 0.087 A | 13.29 ± 0.997 B | 0.040 ± 0.0101 A | |
| 10 | 3.32 ± 0.508 A | 9.80 ± 0.597 B | 0.327 ± 0.0652 A | 0.33 ± 0.150 A | 12.42 ± 2.898 A | 0.038 ± 0.0096 A | |
| 14 | 3.78 ± 0.169 B | 9.04 ± 0.311 A | 0.342 ± 0.0230 A | 0.39 ± 0.097 B | 11.84 ± 0.467 A | 0.047 ± 0.0128 B | |
| Control | 4.90 ± 0.038 C | 10.15 ± 0.083 C | 0.497 ± 0.0012 B | 0.50 ± 0.032 C | 12.48 ± 0.378 AB | 0.062 ± 0.0048 C | |
| Significance | |||||||
| TD | *** | *** | *** | *** | *** | *** | |
| TL | *** | *** | *** | *** | *** | *** | |
| TD × TL | *** | *** | *** | *** | *** | *** | |
| Temperature Duration (Weeks) | Temperature Level (°C) | Soluble Sugars | L-Ascorbic Acid |
|---|---|---|---|
| 1 | 6 | 1.95 ± 0.046 a | 48.68 ± 0.200 b |
| 10 | 1.92 ± 0.038 a | 46.82 ± 0.257 a | |
| 14 | 1.94 ± 0.034 a | 50.74 ± 0.321 d | |
| 2 | 6 | 2.15 ± 0.036 b | 53.42 ± 0.006 e |
| 10 | 2.17 ± 0.034 b | 55.42 ± 0.349 f | |
| 14 | 2.46 ± 0.045 c | 56.13 ± 0.239 g | |
| Control | 18 | 1.99 ± 0.008 a | 49.51 ± 0.054 c |
| Duration (TD) (weeks) | |||
| 1 | 1.94 ± 0.038 A | 48.75 ± 1.687 A | |
| 2 | 2.26 ± 0.151 C | 54.99 ± 1.222 C | |
| Control | 1.99 ± 0.008 B | 49.51 ± 0.054 B | |
| Temperature (TL) (°C) | |||
| 6 | 2.05 ± 0.115 A | 51.05 ± 2.536 B | |
| 10 | 2.05 ± 0.139 A | 51.12 ± 4.604 B | |
| 14 | 2.20 ± 0.279 B | 53.44 ± 2.896 C | |
| Control | 1.99 ± 0.008 A | 49.51 ± 0.054 A | |
| Significance | |||
| TD | *** | *** | |
| TL | *** | *** | |
| TD × TL | *** | *** | |
| Temperature Duration (Weeks) | Temperature Level (°C) | Dry Weight | Chlorophyll a | Chlorophyll b | Carotenoids |
|---|---|---|---|---|---|
| 1 | 6 | 8.56 ± 0.034 a | 5.73 ± 0.319 b | 3.37 ± 0.126 b | 0.78 ± 0.136 a |
| 10 | 8.70 ± 0.031 b | 5.00 ± 0.032 a | 2.86 ± 0.049 a | 0.66 ± 0.023 a | |
| 14 | 8.66 ± 0.056 b | 5.34 ± 0.222 ab | 3.15 ± 0.125 b | 0.71 ± 0.102 a | |
| 2 | 6 | 8.96 ± 0.032 d | 5.46 ± 0.241 ab | 3.13 ± 0.144 ab | 0.77 ± 0.027 a |
| 10 | 9.21 ± 0.026 e | 5.79 ± 0.088 b | 3.26 ± 0.118 b | 0.80 ± 0.030 a | |
| 14 | 9.72 ± 0.031 f | 5.65 ± 0.238 b | 3.20 ± 0.112 b | 0.78 ± 0.095 a | |
| Control | 18 | 8.82 ± 0.021 c | 5.75 ± 0.255 b | 3.32 ± 0.176 b | 0.71 ± 0.033 a |
| Duration (TD) (weeks) | |||||
| 1 | 8.64 ± 0.072 A | 5.36 ± 0.371 A | 3.13 ± 0.238 A | 0.72 ± 0.103 A | |
| 2 | 9.30 ± 0.331 C | 5.63 ± 0.231 B | 3.20 ± 0.126 AB | 0.78 ± 0.056 A | |
| Control | 8.82 ± 0.021 B | 5.75 ± 0.255 B | 3.32 ± 0.176 B | 0.71 ± 0.033 A | |
| Temperature (TL) (°C) | |||||
| 6 | 8.76 ± 0.217 A | 5.60 ± 0.298 A | 3.25 ± 0.179 B | 0.78 ± 0.091 A | |
| 10 | 8.96 ± 0.275 B | 5.40 ± 0.427 A | 3.06 ± 0.230 A | 0.73 ± 0.078 A | |
| 14 | 9.19 ± 0.570 C | 5.50 ± 0.270 A | 3.18 ± 0.113 AB | 0.75 ± 0.100 A | |
| Control | 8.82 ± 0.021 A | 5.75 ± 0.255 A | 3.32 ± 0.176 B | 0.71 ± 0.033 A | |
| Significance | |||||
| TD | *** | * | * | ns | |
| TL | *** | ns | * | ns | |
| TD × TL | *** | *** | *** | ns | |
| Temperature Duration (Weeks) | Temperature Level (°C) | P | K | Mg | Ca | Fe | Cu | Mn | Zn |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 6 | 5784 ± 136.8 d | 29916 ± 989 ab | 1314 ± 38.4 d | 4698 ± 655.6 b | 79.65 ± 2.710 bc | 6.542 ± 0.4539 d | 22.25 ± 0.908 b | 62.71 ± 1.971 c |
| 10 | 5461 ± 130.6 abc | 30652 ± 528 bc | 1322 ± 16.9 d | 4282 ± 105.0 b | 82.71 ± 4.121 c | 5.563 ± 0.1049 bc | 20.40 ± 1.048 ab | 59.48 ± 1.827 ab | |
| 14 | 5602 ± 54.7 cd | 30877 ± 79 bc | 1333 ± 18.2 d | 4476 ± 312.8 b | 101.65 ± 1.028 d | 5.542 ± 0.2097 bc | 22.69 ± 0.661 b | 62.83 ± 0.365 c | |
| 2 | 6 | 5508 ± 191.6 bc | 29108 ± 504 a | 1206 ± 14.2 ab | 4372 ± 33.9 b | 75.54 ± 2.150 b | 5.438 ± 0.0798 bc | 20.48 ± 2.643 ab | 60.04 ± 0.867 b |
| 10 | 5571 ± 138.2 cd | 31287 ± 452 c | 1248 ± 14.0 bc | 4329 ± 84.2 b | 101.77 ± 2.100 d | 5.979 ± 0.6884 cd | 20.23 ± 0.774 ab | 62.73 ± 0.605 c | |
| 14 | 5239 ± 65.4 ab | 28923 ± 334 a | 1159 ± 17.4 a | 3606 ± 143.0 a | 61.40 ± 2.312 a | 4.625 ± 0.1443 a | 18.79 ± 0.689 a | 56.95 ± 0.313 a | |
| Control | 18 | 5214 ± 41.5 a | 29421 ± 167 a | 1290 ± 20.4 cd | 4208 ± 153.5 ab | 80.33 ± 5.001 bc | 5.104 ± 0.1577 ab | 18.65 ± 1.710 a | 57.65 ± 0.733 ab |
| Duration (TD) (weeks) | |||||||||
| 1 | 5616 ± 172.3 C | 30482 ± 727 B | 1323 ± 25.1 C | 4485 ± 422.3 B | 88.00 ± 10.498 B | 5.882 ± 0.5556 B | 21.78 ± 1.312 B | 61.67 ± 2.152 C | |
| 2 | 5439 ± 197.6 B | 29773 ± 1188 A | 1204 ± 40.6 A | 4102 ± 377.4 A | 79.57 ± 17.583 A | 5.347 ± 0.6890 A | 19.83 ± 1.673 A | 59.91 ± 2.534 B | |
| Control | 5214 ± 41.5 A | 29421 ± 167 A | 1290 ± 20.4 B | 4208 ± 153.5 AB | 80.33 ± 5.001 A | 5.104 ± 0.1577 A | 18.65 ± 1.710 A | 57.65 ± 0.733 A | |
| Temperature (TL) (°C) | |||||||||
| 6 | 5646 ± 213.3 C | 29512 ± 845 A | 1260 ± 63.8 AB | 4535 ± 463.7 B | 77.60 ± 3.153 A | 5.990 ± 0.6628 B | 21.37 ± 2.060 B | 61.38 ± 2.006 B | |
| 10 | 5516 ± 137.6 BC | 30970 ± 567 B | 1285 ± 42.1 B | 4306 ± 91.6 AB | 92.24 ± 10.630 B | 5.771 ± 0.5074 B | 20.32 ± 0.858 AB | 61.11 ± 2.144 B | |
| 14 | 5421 ± 202.0 B | 29900 ± 1068 A | 1246 ± 94.3 A | 4041 ± 516.6 A | 81.53 ± 21.578 A | 5.084 ± 0.5180 A | 20.74 ± 2.174 AB | 59.89 ± 3.163 B | |
| Control | 5214 ± 41.5 A | 29421 ± 167 A | 1290 ± 20.4 B | 4208 ± 153.5 AB | 80.33 ± 5.001 A | 5.104 ± 0.1577 A | 18.65 ± 1.710 A | 57.65 ± 0.733 A | |
| Significance | |||||||||
| TD | *** | ** | *** | * | *** | ** | ** | *** | |
| TL | *** | *** | ** | * | *** | *** | * | *** | |
| TD × TL | *** | *** | *** | *** | *** | *** | ** | *** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalisz, A.; Gil, J.; Kunicki, E.; Sękara, A.; Sałata, A.; Caruso, G. Different Temperature Regimes Influenced the Quality of Broccoli Seedlings, Which Caused a Change in the Chemical Composition of Mature Heads. Agronomy 2021, 11, 1806. https://doi.org/10.3390/agronomy11091806
Kalisz A, Gil J, Kunicki E, Sękara A, Sałata A, Caruso G. Different Temperature Regimes Influenced the Quality of Broccoli Seedlings, Which Caused a Change in the Chemical Composition of Mature Heads. Agronomy. 2021; 11(9):1806. https://doi.org/10.3390/agronomy11091806
Chicago/Turabian StyleKalisz, Andrzej, Joanna Gil, Edward Kunicki, Agnieszka Sękara, Andrzej Sałata, and Gianluca Caruso. 2021. "Different Temperature Regimes Influenced the Quality of Broccoli Seedlings, Which Caused a Change in the Chemical Composition of Mature Heads" Agronomy 11, no. 9: 1806. https://doi.org/10.3390/agronomy11091806
APA StyleKalisz, A., Gil, J., Kunicki, E., Sękara, A., Sałata, A., & Caruso, G. (2021). Different Temperature Regimes Influenced the Quality of Broccoli Seedlings, Which Caused a Change in the Chemical Composition of Mature Heads. Agronomy, 11(9), 1806. https://doi.org/10.3390/agronomy11091806