
Coupling between the Basic Replicon and the Kis-Kid Maintenance System of Plasmid R1: Modulation by Kis Antitoxin Levels and Involvement in Control of Plasmid Replication

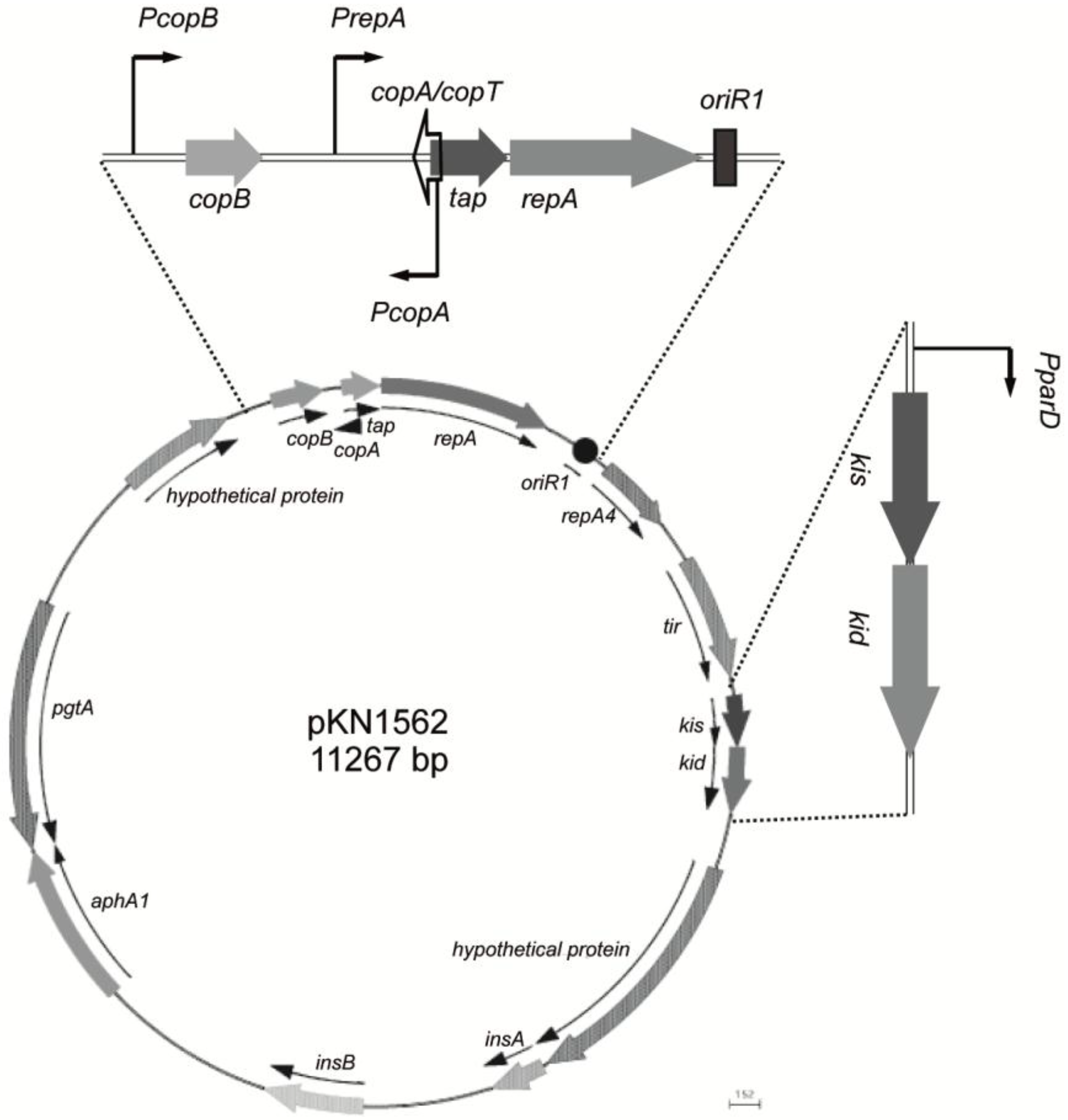{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results
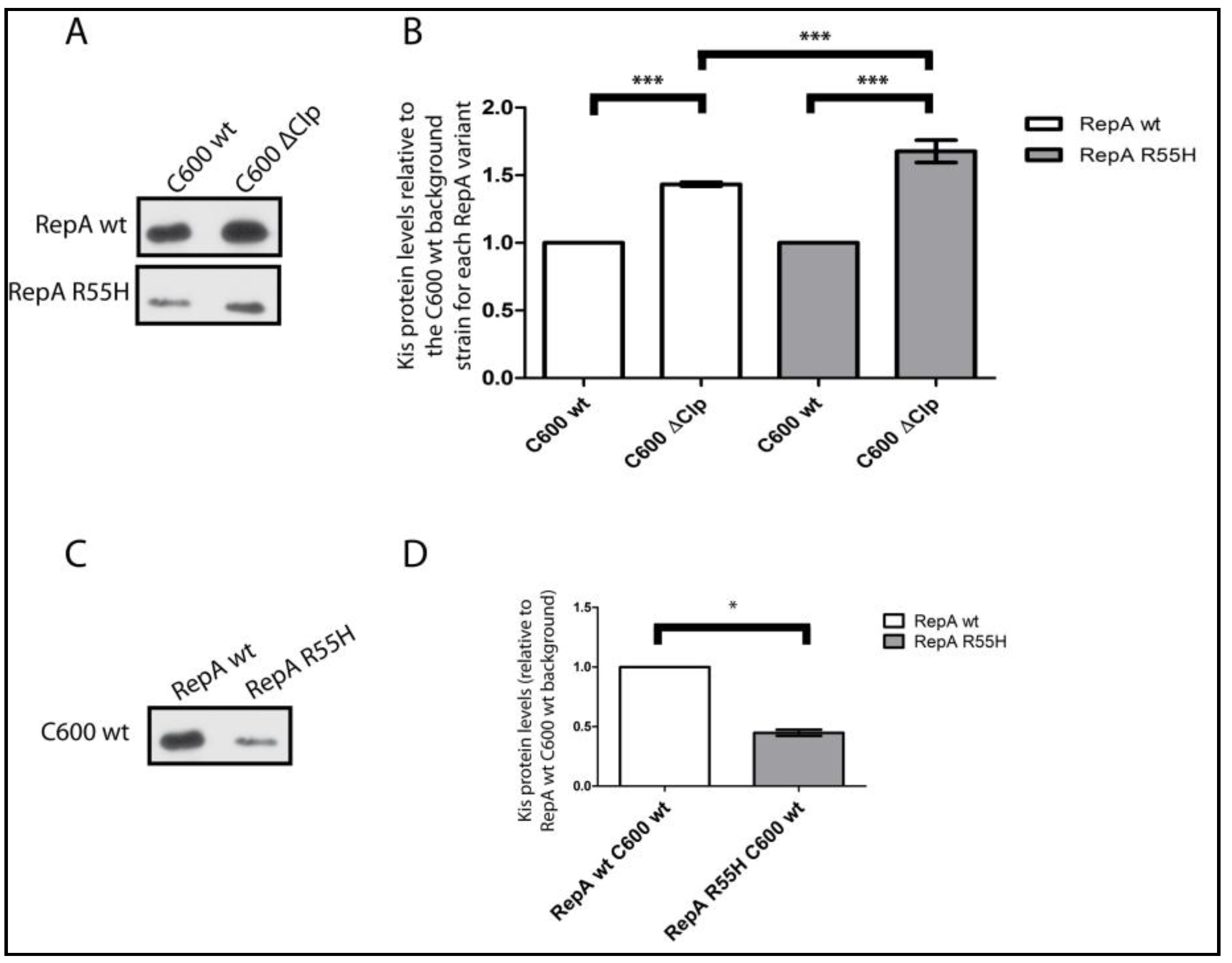
2.1. Kis Antitoxin Levels Decrease in Cells Containing a pKN1562 Replication Mutant and Increase in a clpP− Background
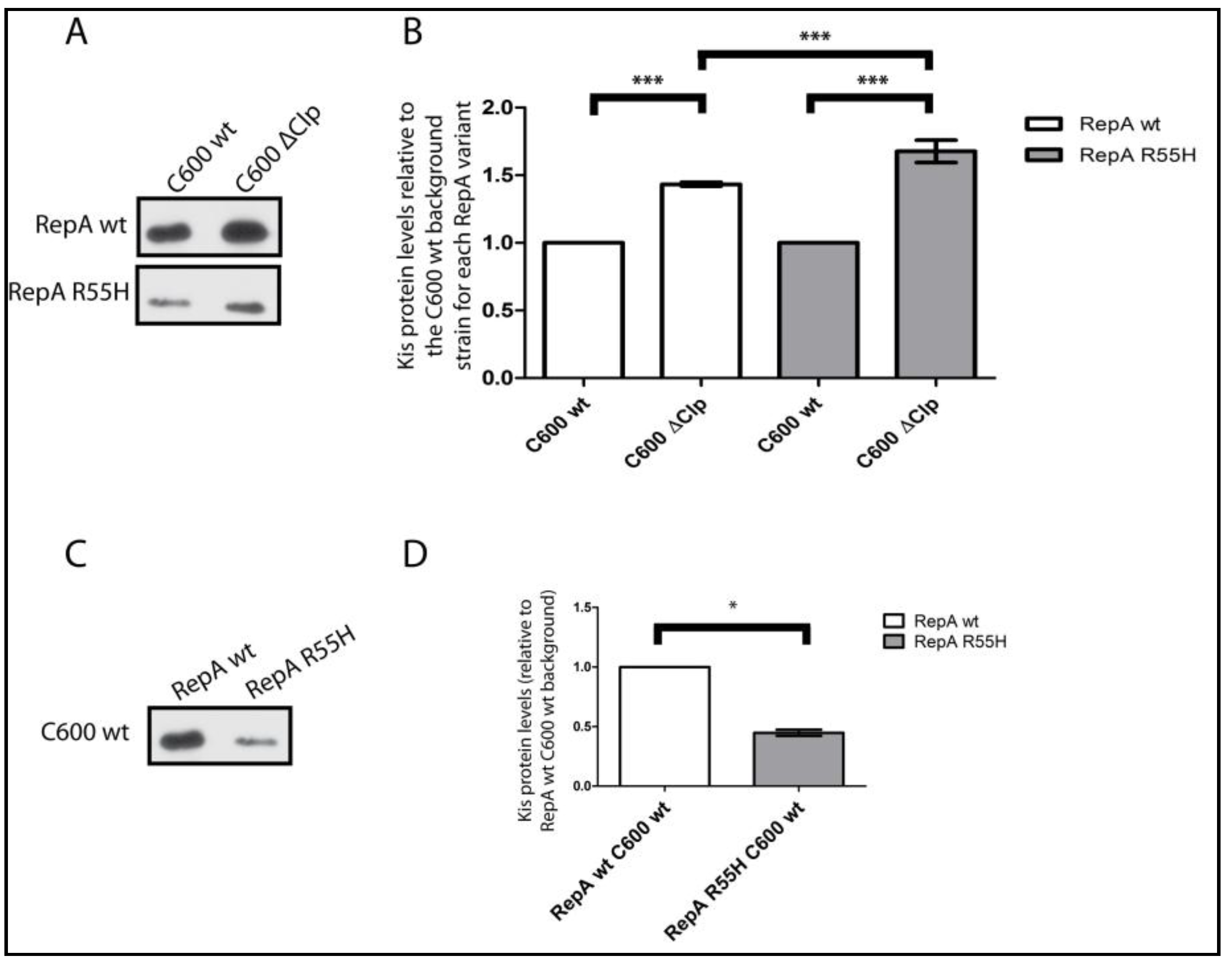
2.2. Effects of Increasing the Stability of the Kis Antitoxin on the Modular Coupling

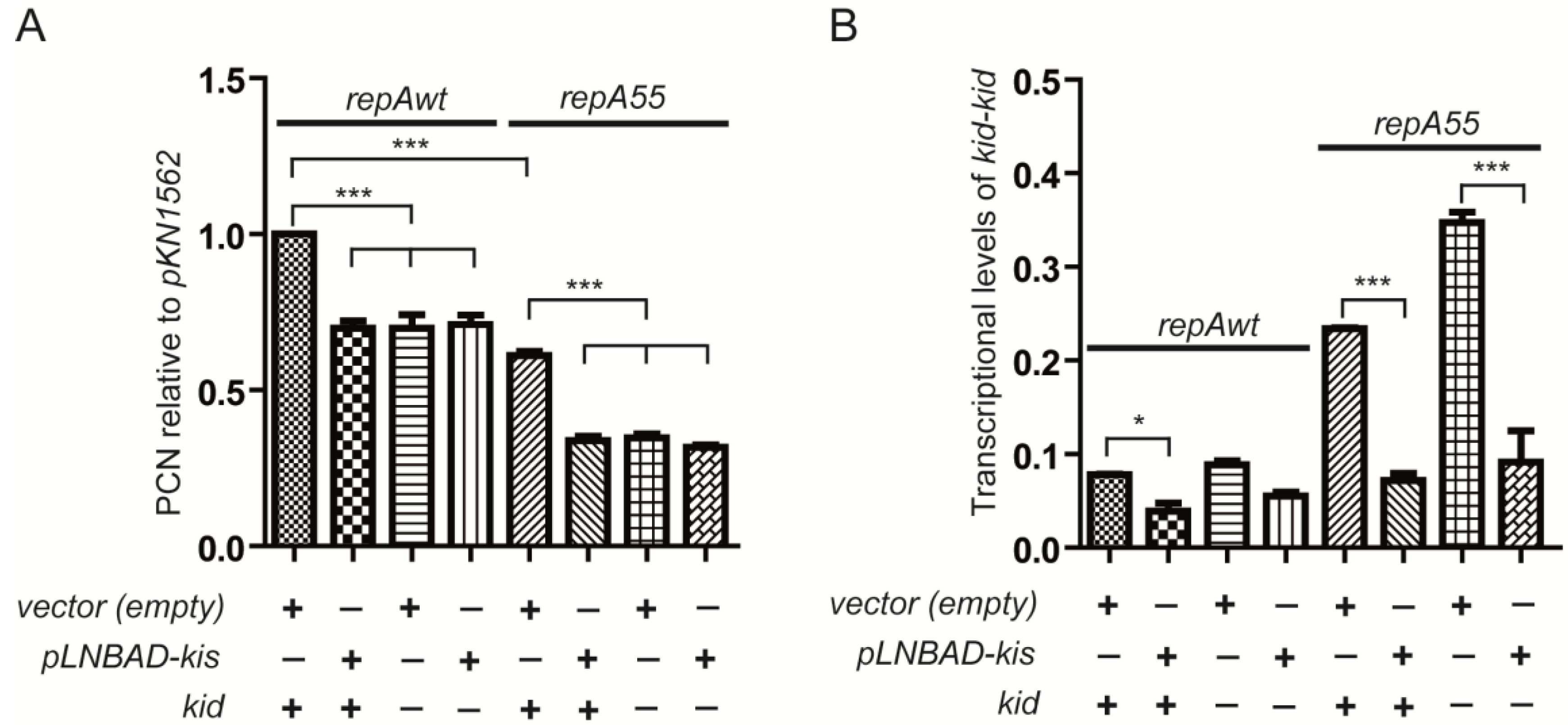
2.3. Effects of Overproducing the Kis Antitoxin on the Modular Coupling
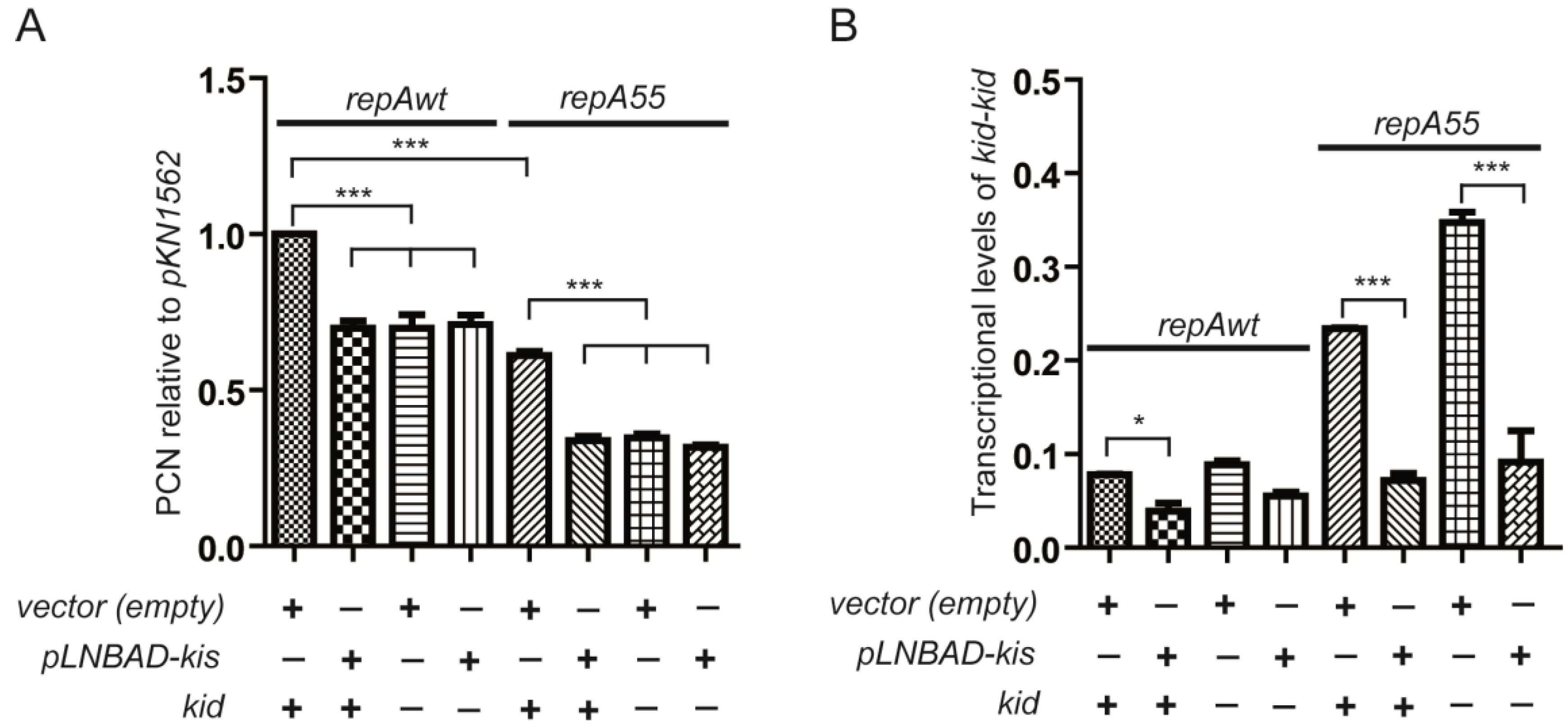
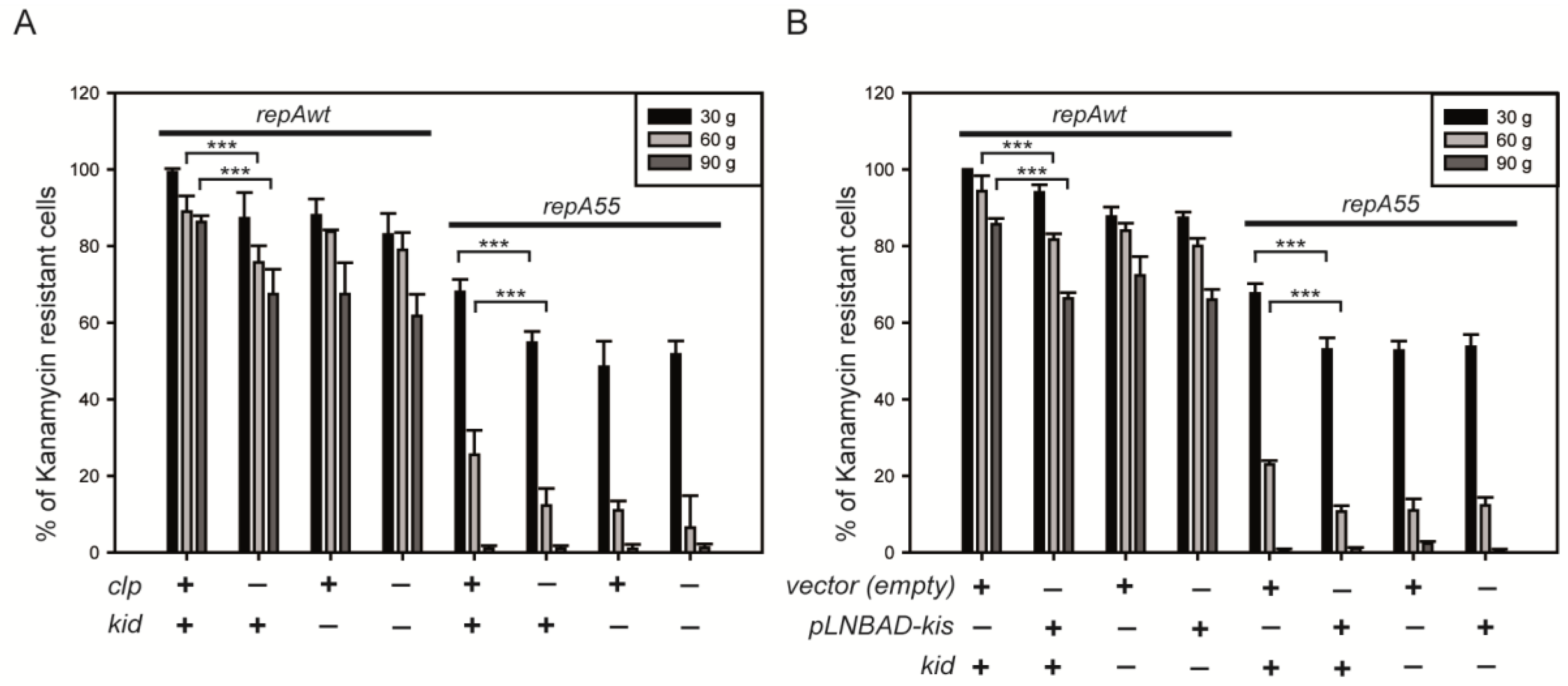
2.4. Coupling between Maintenance Modules and Plasmid Stability
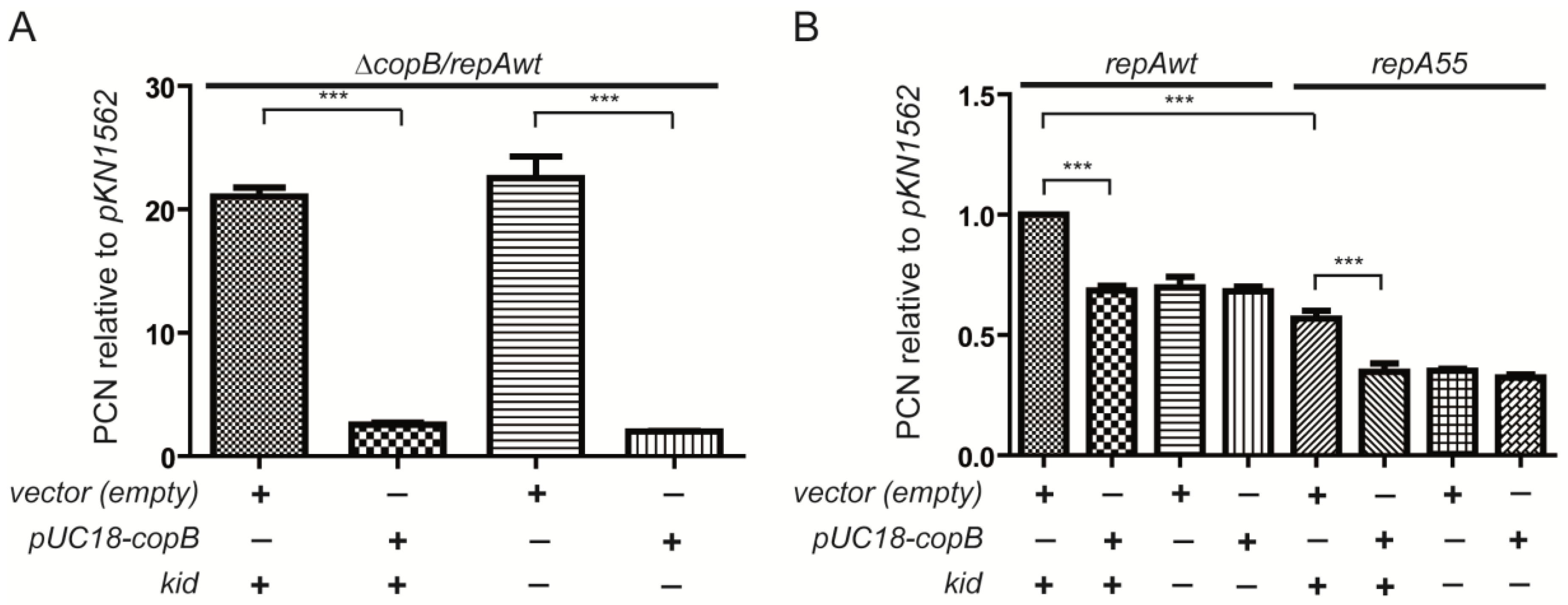
2.5. Excess of CopB Abolishes Replication Rescue
3. Discussion
3.1. Modular Coupling between Maintenance Modules and Efficiency of Plasmid wt Replication
3.2. The Pathway: Kis Antitoxin Acts as the Switch Connecting Plasmid Replication and Toxin-Antitoxin Modules
3.3. On the Inhibition of the Replication Rescue by Excess of CopB
3.4. Coordination of Plasmid Maintenance Functions in Different Systems
4. Experimental Section
4.1. Cell Cultures, Strains and Plasmids
4.2. Replication Interference Phenotype
4.3. Plasmid Stability and Statistical Analysis
4.4. Plasmid Copy Number Determinations
4.5. Quantification of KIS-KID MRNA LEVELS
4.6. Determination of Kis Antitoxin Levels by Western Blotting-3XFLAG
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nordström, K. Plasmid R1—replication and its control. Plasmid 2006, 55, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, R.; Diaz, R. Differential binding of wild-type and a mutant RepA protein to oriR sequence suggests a model for the initiation of plasmid R1 replication. J. Mol. Biol. 1992, 228, 787–802. [Google Scholar] [CrossRef] [PubMed]
- Nordström, K.; Molin, S.; Light, J. Control of replication of bacterial plasmids: Genetics, molecular biology, and physiology of the plasmid R1 system. Plasmid 1984, 12, 71–90. [Google Scholar] [CrossRef] [PubMed]
- Nordström, K.; Wagner, E.G. Kinetic aspects of control of plasmid replication by antisense RNA. Trends Biochem. Sci. 1994, 19, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Uhlin, B.E.; Molin, S.; Gustafsson, P.; Nordström, K. Plasmids with temperature-dependent copy number for amplification of cloned genes and their products. Gene 1979, 6, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Riise, E.; Stougaard, P.; Bindslev, B.; Nordström, K.; Molin, S. Molecular cloning and functional characterization of a copy number control gene (copB) of plasmid R1. J. Bacteriol. 1982, 151, 1136–1145. [Google Scholar] [PubMed]
- Riise, E.; Molin, S. Purification and characterization of the CopB replication control protein, and precise mapping of its target site in the R1 plasmid. Plasmid 1986, 15, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Light, J.; Riise, E.; Molin, S. Transcription and its regulation in the basic replicon region of plasmid R1. Mol. Gen. Genet. 1985, 198, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Olsson, J.A.; Paulsson, J.; Nordström, K. Effect of the copB auxiliary replication control system on stability of maintenance of par (+) plasmid R1. J. Bacteriol. 2004, 186, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Molin, S.; Stougaard, P.; Uhlin, B.E.; Gustafsson, P.; Nordström, K. Clustering of genes involved in replication, copy number control, incompatibility, and stable maintenance of the resistance plasmid R1drd-19. J. Bacteriol. 1979, 138, 70–79. [Google Scholar] [PubMed]
- Bravo, A.; de Torrontegui, G.; Diaz, R. Identification of components of a new stability system of plasmid R1, parD, that is close to the origin of replication of this plasmid. Mol. Gen. Genet. 1987, 210, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Diago-Navarro, E.; Hernandez-Arriaga, A.M.; Lopez-Villarejo, J.; Muñoz-Gomez, A.J.; Kamphuis, M.B.; Boelens, R.; Lemonnier, M.; Diaz-Orejas, R. parD toxin-antitoxin system of plasmid R1—Basic contributions, biotechnological applications and relationships with closely-related toxin-antitoxin systems. FEBS J. 2010, 277, 3097–3117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, Y.; Zhu, L.; Suzuki, M.; Inouye, M. Interference of mRNA function by sequence-specific endoribonuclease PemK. J. Biol. Chem. 2004, 279, 20678–20684. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Gomez, A.J.; Lemonnier, M.; Santos-Sierra, S.; Berzal-Herranz, A.; Diaz-Orejas, R. RNase/anti-RNase activities of the bacterial parD toxin-antitoxin system. J. Bacteriol 2005, 187, 3151–3157. [Google Scholar] [CrossRef] [PubMed]
- Kamphuis, M.B.; Bonvin, A.M.; Monti, M.C.; Lemonnier, M.; Munoz-Gomez, A.; van den Heuvel, R.H.; Diaz-Orejas, R.; Boelens, R. Model for RNA binding and the catalytic site of the RNase kid of the bacterial parD toxin-antitoxin system. J. Mol. Biol. 2006, 357, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Echevarria, M.J.; Berzal-Herranz, A.; Gerdes, K.; Diaz-Orejas, R. The kis and kid genes of the parD maintenance system of plasmid R1 form an operon that is autoregulated at the level of transcription by the co-ordinated action of the Kis and Kid proteins. Mol. Microbiol. 1991, 5, 2685–2693. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.C.; Hernandez-Arriaga, A.M.; Kamphuis, M.B.; Lopez-Villarejo, J.; Heck, A.J.; Boelens, R.; Diaz-Orejas, R.; van den Heuvel, R.H. Interactions of Kid-Kis toxin-antitoxin complexes with the parD operator-promoter region of plasmid R1 are piloted by the Kis antitoxin and tuned by the stoichiometry of Kid-Kis oligomers. Nucleic Acids Res. 2007, 35, 1737–1749. [Google Scholar] [CrossRef] [PubMed]
- Ortega, S.; de Torrontegui, G.; Diaz, R. Isolation and characterization of a conditional replication mutant of the antibiotic resistance factor R1 affected in the gene of the replication protein RepA. Mol. Gen. Genet. 1989, 217, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Echevarria, M.J.; de-la-Torre, M.A.; Diaz-Orejas, R. A mutation that decreases the efficiency of plasmid R1 replication leads to the activation of parD, a killer stability system of the plasmid. FEMS Microbiol. Lett. 1995, 130, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, B.; Madine, M.A.; de la Cueva-Mendez, G. Kid cleaves specific mRNAs at UUACU sites to rescue the copy number of plasmid R1. EMBO J. 2005, 24, 3459–3469. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, B.; Nair, R.; Bermejo-Rodriguez, C.; Preston, M.A.; Agu, C.A.; Wang, X.; Bernal, J.A.; Sherratt, D.J.; de la Cueva-Mendez, G. Toxin Kid uncouples DNA replication and cell division to enforce retention of plasmid R1 in Escherichia coli cells. Proc. Natl. Acad. Sci. USA 2014, 111, 2734–2739. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Villarejo, J.; Diago-Navarro, E.; Hernandez-Arriaga, A.M.; Diaz-Orejas, R. Kis antitoxin couples plasmid R1 replication and parD (kis,kid) maintenance modules. Plasmid 2012, 67, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Lioy, V.S.; Pratto, F.; de la Hoz, A.B.; Ayora, S.; Alonso, J.C. Plasmid pSM19035, a model to study stable maintenance in firmicutes. Plasmid 2010, 64, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.M.; Smith, C.A.; Shingler, V.; Cross, M.A.; Hussain, A.A.; Pinkney, M. Regulation of replication and maintenance functions of broad host-range plasmid RK2. Basic Life Sci. 1985, 30, 261–276. [Google Scholar] [PubMed]
- Cevallos, M.A.; Cervantes-Rivera, R.; Gutierrez-Rios, R.M. The repABC plasmid family. Plasmid 2008, 60, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Field, C.M.; Summers, D.K. Multicopy plasmid stability: Revisiting the dimer catastrophe. J. Theor. Biol. 2011, 291, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Diago-Navarro, E.; Hernandez-Arriaga, A.M.; Kubik, S.; Konieczny, I.; Diaz-Orejas, R. Cleavage of the antitoxin of the parD toxin-antitoxin system is determined by the ClpAP protease and is modulated by the relative ratio of the toxin and the antitoxin. Plasmid 2013, 70, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.T.; Malkus, P.; Paulsson, J. New quantitative methods for measuring plasmid loss rates reveal unexpected stability. Plasmid 2013, 70, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Kolatka, K.; Kubik, S.; Rajewska, M.; Konieczny, I. Replication and partitioning of the broad host-range plasmid RK2. Plasmid 2010, 64, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Summers, D.K.; Sherratt, D.J. Multimerization of high copy number plasmids causes instability: Co1E1 encodes a determinant essential for plasmid monomerization and stability. Cell 1984, 36, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Field, C.M.; Summers, D.K. Indole inhibition of ColE1 replication contributes to stable plasmid maintenance. Plasmid 2012, 67, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Maniatis, T.; Fritsch, E.F.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbour Laboratory: Cold Spring Harbour, NY, USA, 1982. [Google Scholar]
- Kandror, O.; Busconi, L.; Sherman, M.; Goldberg, A.L. Rapid degradation of an abnormal protein in Escherichia coli involves the chaperones GroEl and GroES. J. Biol. Chem. 1994, 269, 23575–23582. [Google Scholar] [PubMed]
- Appleyard, R.K. Segregation of new lysogenic types during growth of a doubly lysogenic strain derived from Escherichia coli K12. Genetics 1954, 39, 440–452. [Google Scholar] [PubMed]
- Diago-Navarro, E.; Kamphuis, M.B.; Boelens, R.; Barendregt, A.; Heck, A.J.; van den Heuvel, R.H.; Diaz-Orejas, R. A mutagenic analysis of the RNase mechanism of the bacterial Kid toxin by mass spectrometry. FEBS J. 2009, 276, 4973–4986. [Google Scholar] [CrossRef] [PubMed]
- Lemonnier, M.; Santos-Sierra, S.; Pardo-Abarrio, C.; Diaz-Orejas, R. Identification of residues of the Kid toxin involved in autoregulation of the parD system. J. Bacteriol. 2004, 186, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fristsch, E.F.; Maniatis, T. Molecular Cloning; Cold Spring Harbour Laboratory: Cold Spring Harbour, NY, USA, 1989. [Google Scholar]
- Hashimoto, T.; Sekiguchi, M. Isolation of temperature-sensitive mutants of R plasmid by in vitro mutagenesis with hydroxylamine. J. Bacteriol. 1976, 127, 1561–1563. [Google Scholar] [PubMed]
- Lee, C.; Kim, J.; Shin, S.G.; Hwang, S. Absolute and relative qPCR quantification of plasmid copy number in Escherichia coli. J. Biotechnol. 2006, 123, 273–280. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Villarejo, J.; Lobato-Márquez, D.; Díaz-Orejas, R. Coupling between the Basic Replicon and the Kis-Kid Maintenance System of Plasmid R1: Modulation by Kis Antitoxin Levels and Involvement in Control of Plasmid Replication. Toxins 2015, 7, 478-492. https://doi.org/10.3390/toxins7020478
López-Villarejo J, Lobato-Márquez D, Díaz-Orejas R. Coupling between the Basic Replicon and the Kis-Kid Maintenance System of Plasmid R1: Modulation by Kis Antitoxin Levels and Involvement in Control of Plasmid Replication. Toxins. 2015; 7(2):478-492. https://doi.org/10.3390/toxins7020478
Chicago/Turabian StyleLópez-Villarejo, Juan, Damián Lobato-Márquez, and Ramón Díaz-Orejas. 2015. "Coupling between the Basic Replicon and the Kis-Kid Maintenance System of Plasmid R1: Modulation by Kis Antitoxin Levels and Involvement in Control of Plasmid Replication" Toxins 7, no. 2: 478-492. https://doi.org/10.3390/toxins7020478
APA StyleLópez-Villarejo, J., Lobato-Márquez, D., & Díaz-Orejas, R. (2015). Coupling between the Basic Replicon and the Kis-Kid Maintenance System of Plasmid R1: Modulation by Kis Antitoxin Levels and Involvement in Control of Plasmid Replication. Toxins, 7(2), 478-492. https://doi.org/10.3390/toxins7020478