Biological and Chemical Management of Aspergillus carbonarius and Ochratoxin A in Vineyards


Abstract
1. Introduction
2. Results
2.1. Evaluation of Plant Protection Products in Controlling Aspergillus Bunch Sour Rot in Vineyards
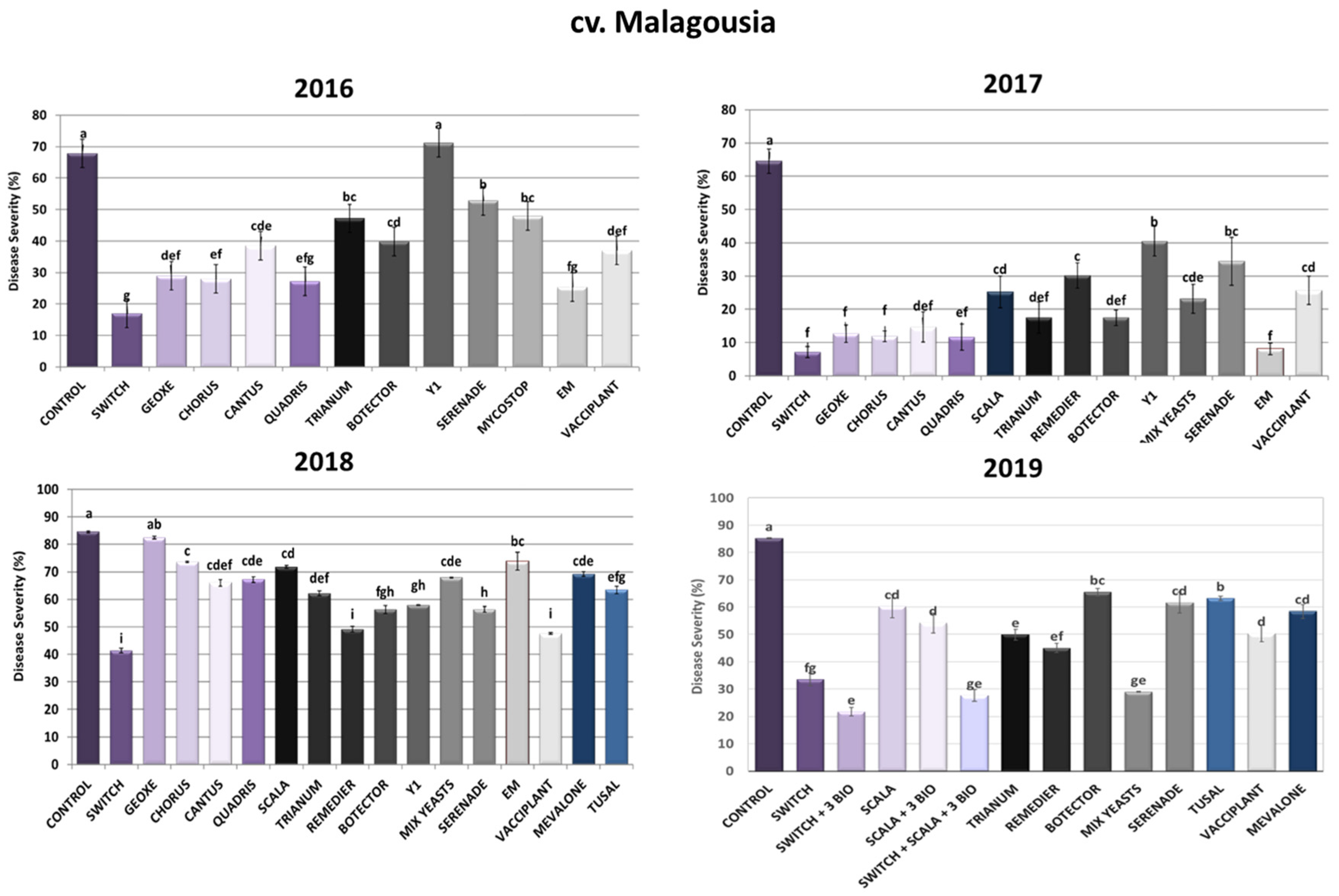
2.1.1. cv. Malagousia Disease Severity Assessment
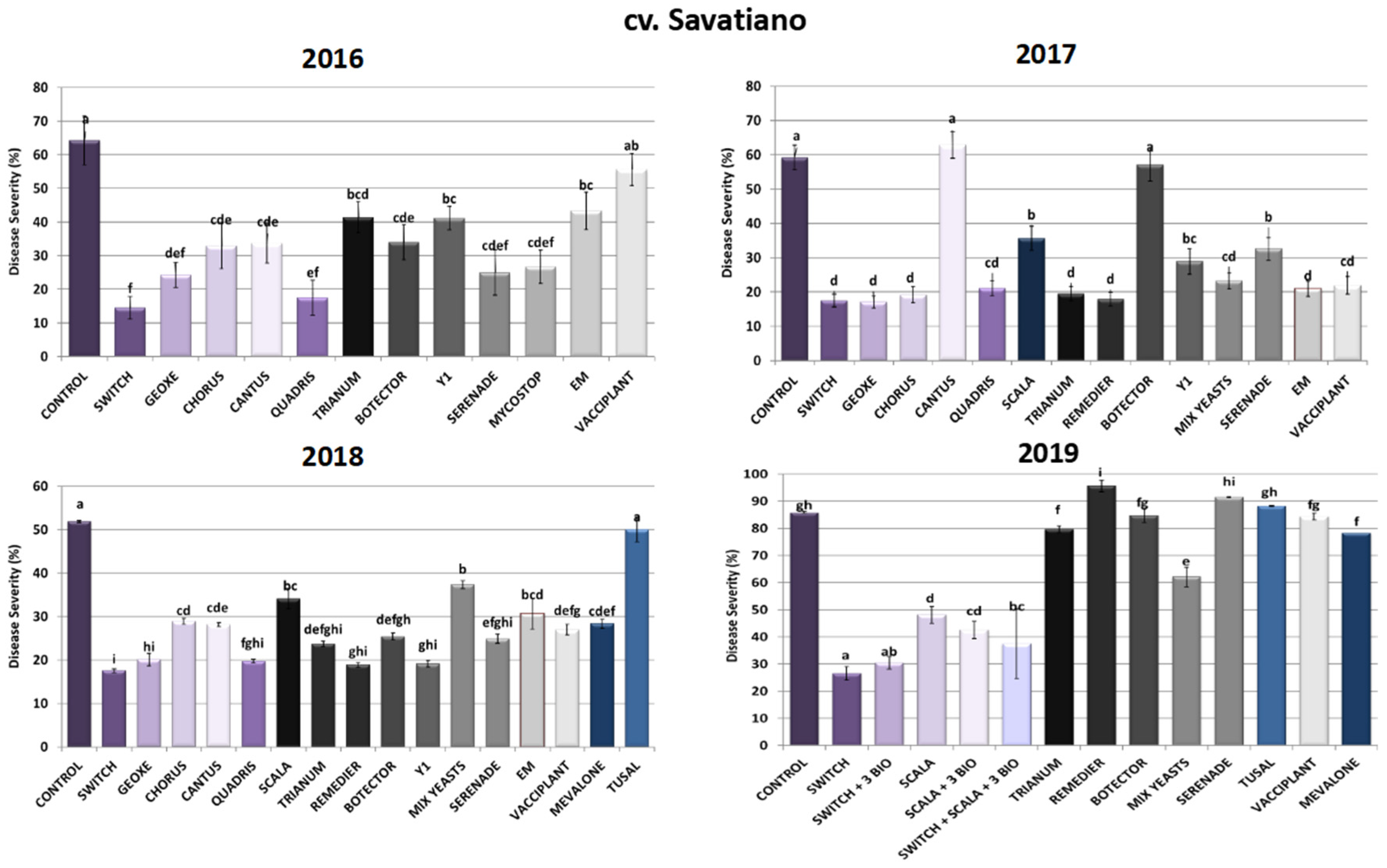
2.1.2. cv. Savatiano Disease Severity Assessment
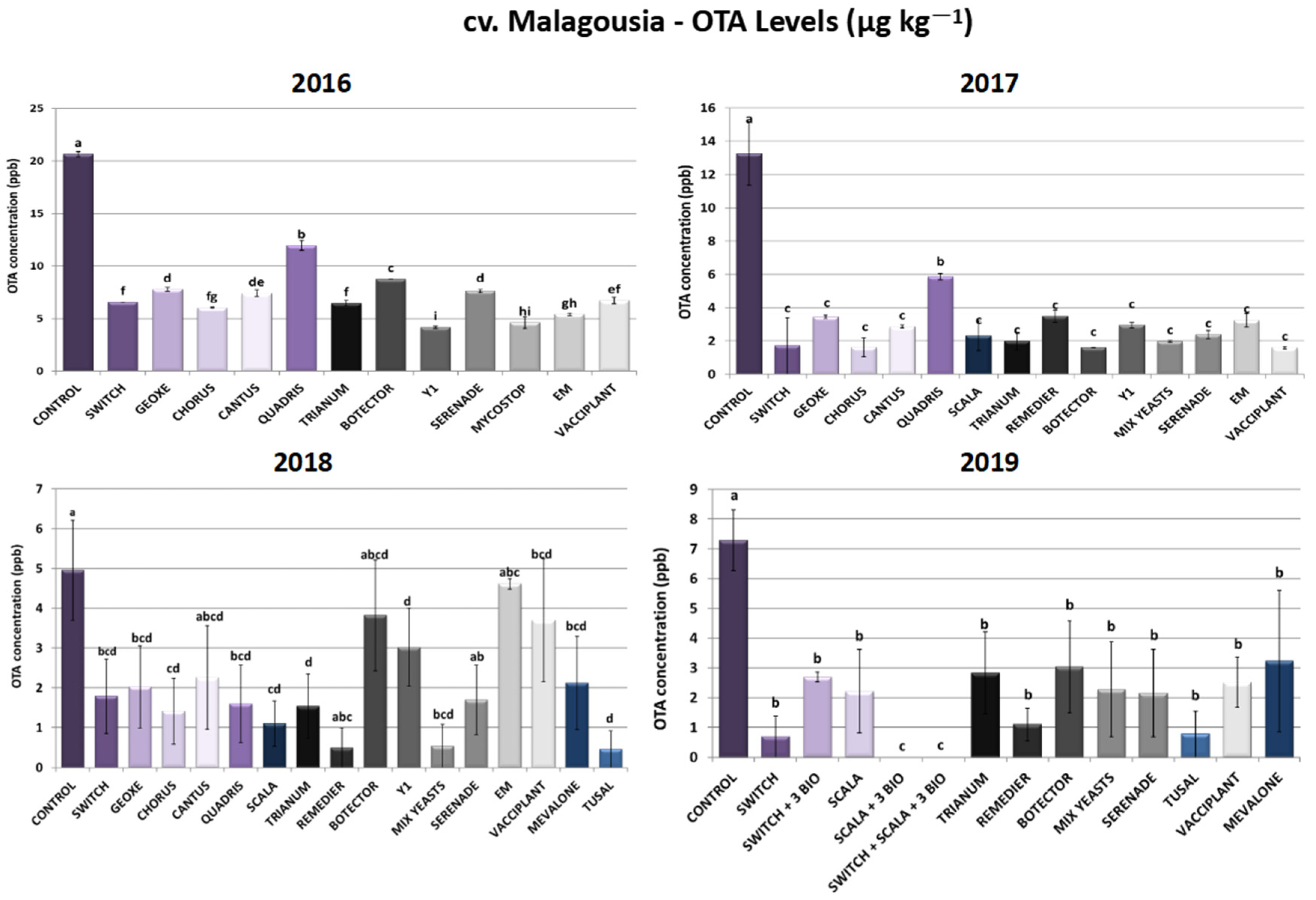
2.1.3. cv. Malagousia—Ochratoxin A Levels
2.1.4. cv. Savatiano—Ochratoxin A Levels
2.2. In Vitro Assessment of A. carbonarius’ Sensitivity to Fungicides
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Field Trials
5.1.1. Experimental Design—Plant Protection Products
5.1.2. Disease Assessment
5.1.3. Ochratoxin A (OTA) Quantification
5.2. Sensitivity Tests on Active Substances
5.2.1. Mycelial Growth Inhibition
5.2.2. Conidial Germination Inhibition
5.3. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ortiz-Villeda, B.; Lobos, O.; Aguilar-Zuniga, K.; Carrasco-Sánchez, V. Ochratoxins in Wines: A Review of Their Occurrence in the Last Decade, Toxicity, and Exposure Risk in Humans. Toxins 2021, 13, 478. [Google Scholar] [CrossRef] [PubMed]
- Some Naturally Occurring Substances. In IARC Monographs on the Evaluation of Carcinogenic Risk of Chemicals to Man; World Health Organization, International Agency for Research on Cancer: Lyon, France, 1976; Volume 10, pp. 191–197.
- The European Food Safety Authority. Opinion of the Scientific Panel on contaminants in the Food Chain of the EFSA on a request from the Commission related to ochratoxin A in food. EFSA J. 2006, 365, 1–56. [Google Scholar]
- Visconti, A.; Perrone, G.; Cozzi, G.; Solfrizzo, M. Managing ochratoxin A risk in the grape-wine food chain. Food Addit. Contam. Part A 2008, 25, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Ben Miri, Y.; Benabdallah, A.; Chentir, I.; Djenane, D.; Luvisi, A.; De Bellis, L. Comprehensive Insights into Ochratoxin A: Occurrence, Analysis, and Control Strategies. Foods 2024, 13, 1184. [Google Scholar] [CrossRef]
- Amézqueta, S.; Schorr-Galindo, S.; Murillo-Arbizu, M.; González-Peñas, E.; López de Cerain, A.; Guiraud, J. OTA-producing fungi in foodstuffs: A review. Food Control 2012, 26, 259–268. [Google Scholar] [CrossRef]
- Alshannaq, A.; Yu, J.-H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef]
- Freire, L.; Braga, P.A.C.; Furtado, M.M.; Delafiori, J.; Dias-Audibert, F.L.; Pereira, G.E.; Reyes, F.G.; Catharino, R.R.; Sant’Ana, A.S. From grape to wine: Fate of ochratoxin A during red, rose, and white winemaking process and the presence of ochratoxin derivatives in the final products. Food Control 2020, 113, 107167. [Google Scholar] [CrossRef]
- Leong, S.L.; Hocking, A.D.; Pitt, J.I.; Kazi, B.A.; Emmett, R.W.; Scott, E.S. Australian research on ochratoxigenic fungi and ochratoxin A. Int. J. Food Microbiol. 2006, 111, S10–S17. [Google Scholar] [CrossRef]
- Cervini, C.; Gallo, A.; Piemontese, L.; Magistà, D.; Logrieco, A.F.; Ferrara, M.; Solfrizzo, M.; Perrone, G. Effects of temperature and water activity change on ecophysiology of ochratoxigenic Aspergillus carbonarius in field-simulating conditions. Int. J. Food Microbiol. 2020, 315, 108420. [Google Scholar] [CrossRef]
- Wang, G.; Li, E.; Gallo, A.; Perrone, G.; Varga, E.; Ma, J.; Yang, B.; Tai, B.; Xing, F. Impact of environmental factors on ochratoxin A: From natural occurrence to control strategy. Environ. Pollut. 2023, 317, 120767. [Google Scholar] [CrossRef]
- Passamani, F.R.F.; Hernandes, T.; Lopes, N.A.; Bastos, S.C.; Santiago, W.D.; Cardoso, M.D.G.; Batist, L.R. Effect of Temperature, Water Activity, and pH on Growth and Production of Ochratoxin A by Aspergillus niger and Aspergillus carbonarius from Brazilian Grapes. J. Food Prot. 2014, 77, 1947–1952. [Google Scholar] [CrossRef] [PubMed]
- Kontaxakis, E.; Fysarakis, I.; Lydakis, D.; Magan, N. Farming System Effect on the Incidence of Aspergillus carbonarius on Kotsifali Grapes and Ochratoxin A Occurrence in Wines of Crete. J. Food Prot. 2020, 83, 1796–1800. [Google Scholar] [CrossRef]
- Testempasis, S.I.; Kamou, N.N.; Papadakis, E.-N.; Menkissoglu-Spiroudi, U.; Karaoglanidis, G.S. Conventional vs. organic vineyards: Black Aspergilli population structure, mycotoxigenic capacity and mycotoxin contamination assessment in wines, using a new Q-TOF MS-MS detection method. Food Control 2022, 136, 108860. [Google Scholar] [CrossRef]
- Cervini, C. Aspergillus from Grapes: Ochratoxin A Risk in Relation to Climate Change and New Strategies for Reducing Contamination. Ph.D. Thesis, Università degli studi di Foggia, Foggia, Italy, 2020. [Google Scholar]
- Mondani, L.; Palumbo, R.; Tsitsigiannis, D.; Perdikis, D.; Mazzoni, E.; Battilani, P. Pest Management and Ochratoxin A Contamination in Grapes: A Review. Toxins 2020, 12, 303. [Google Scholar] [CrossRef] [PubMed]
- Tjamos, S.E.; Antoniou, P.P.; Kazantzidou, A.; Antonopoulos, D.F.; Papageorgiou, I.; Tjamos, E.C. Aspergillus niger and Aspergillus carbonarius in Corinth Raisin and Wine-Producing Vineyards in Greece: Population Composition, Ochratoxin A Production and Chemical Control. J. Phytopathol. 2004, 152, 250–255. [Google Scholar] [CrossRef]
- Bellí, N.; Marín, S.; Argilés, E.; Ramos, A.J.; Sanchis, V. Effect of Chemical Treatments on Ochratoxigenic Fungi and Common Mycobiota of Grapes (Vitis vinifera). J. Food Prot. 2007, 70, 157–163. [Google Scholar] [CrossRef]
- Farbo, M.G.; Urgeghe, P.P.; Fiori, S.; Marcello, A.; Oggiano, S.; Balmas, V.; Hassan, Z.U.; Jaoua, S.; Migheli, Q. Effect of yeast volatile organic compounds on ochratoxin A-producing Aspergillus carbonarius and A. ochraceus. Int. J. Food Microbiol. 2018, 284, 1–10. [Google Scholar] [CrossRef]
- Oufensou, S.; Ul Hassan, Z.; Balmas, V.; Jaoua, S.; Migheli, Q. Perfume Guns: Potential of Yeast Volatile Organic Compounds in the Biological Control of Mycotoxin-Producing Fungi. Toxins 2023, 15, 45. [Google Scholar] [CrossRef]
- Jiang, C.; Shi, J.; Liu, Y.; Zhu, C. Inhibition of Aspergillus carbonarius and fungal contamination in table grapes using Bacillus subtilis. Food Control 2014, 35, 41–48. [Google Scholar] [CrossRef]
- Jiang, C.; Li, Z.; Shi, Y.; Guo, D.; Pang, B.; Chen, X.; Shao, D.; Liu, Y.; Shi, J. Bacillus subtilis inhibits Aspergillus carbonarius by producing iturin A, which disturbs the transport, energy metabolism, and osmotic pressure of fungal cells as revealed by transcriptomics analysis. Int. J. Food Microbiol. 2020, 330, 108783. [Google Scholar] [CrossRef]
- Higazy, N.S.; Saleh, A.E.; Hassan, Z.U.; Al Thani, R.; Migheli, Q.; Jaoua, S. Investigation and application of Bacillus pumilus QBP344-3 in the control of Aspergillus carbonarius and ochratoxin A contamination. Food Control 2021, 119, 107464. [Google Scholar] [CrossRef]
- Silveira, R.D.; Veras, F.F.; Bach, E.; Manfroi, V.; Brandelli, A.; Welke, J.E. Aspergillus carbonarius-derived ochratoxins are inhibited by Amazonian Bacillus spp. used as a biocontrol agent in grapes. Food Addit. Contam. Part A 2022, 39, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Kalagatur, N.K.; Gurunathan, S.; Kamasani, J.R.; Gunti, L.; Kadirvelu, K.; Mohan, C.D.; Rangappa, S.; Prasad, R.; Almeida, F.; Mudili, V.; et al. Inhibitory effect of C. zeylanicum, C. longa, O. basilicum, Z. officinale, and C. martini essential oils on growth and ochratoxin A content of A. ochraceous and P. verrucosum in maize grains. Biotechnol. Rep. 2020, 27, e00490. [Google Scholar] [CrossRef] [PubMed]
- Mirza Alizadeh, A.; Golzan, S.A.; Mahdavi, A.; Dakhili, S.; Torki, Z.; Hosseini, H. Recent advances on the efficacy of essential oils on mycotoxin secretion and their mode of action. Crit. Rev. Food Sci. Nutr. 2022, 62, 4726–4751. [Google Scholar] [CrossRef]
- Zjalic, S.; Markov, K.; Loncar, J.; Jakopovic, Z.; Beccaccioli, M.; Reverberi, M. Biocontrol of Occurrence Ochratoxin A in Wine: A Review. Toxins 2024, 16, 277. [Google Scholar] [CrossRef]
- Tsolakis, H.; Corona, O.; Pulizzi, A.S.; Grippi, F.; Mondello, V. Incidence of grapevine moth Lobesia botrana (Den. & Schiff.) on occurrence of ochratoxin A in grapes. IOBC/WPRS Bull. 2008, 36, 363–368. [Google Scholar]
- Cozzi, G.; Somma, S.; Haidukowski, M.; Logrieco, A.F. Ochratoxin A Management in Vineyards by Lobesia botrana Biocontrol. Toxins 2013, 5, 49–59. [Google Scholar] [CrossRef]
- Dimakopoulou, M.; Tjamos, S.E.; Antoniou, P.P.; Pietri, A.; Battilani, P.; Avramidis, N.; Markakis, E.A.; Tjamos, E.C. Phyllosphere grapevine yeast Aureobasidium pullulans reduces Aspergillus carbonarius (sour rot) incidence in wine-producing vineyards in Greece. Biol. Control 2008, 46, 158–165. [Google Scholar] [CrossRef]
- Pfliegler, W.P.; Pusztahelyi, T.; Pócsi, I. Mycotoxins—Prevention and decontamination by yeasts. J. Basic. Microbiol. 2015, 55, 805–818. [Google Scholar] [CrossRef]
- Petruzzi, L.; Sinigaglia, M.; Corbo, M.R.; Campaniello, D.; Speranza, B.; Bevilacqua, A. Decontamination of ochratoxin A by yeasts: Possible approaches and factors leading to toxin removal in wine. Appl. Microbiol. Biotechnol. 2014, 98, 6555–6567. [Google Scholar] [CrossRef]
- Xu, X.; Pang, M.; Liu, J.; Wang, Y.; Wu, X.; Huang, K.; Liang, Z. Genome mining reveals the genes of carboxypeptidase for OTA-detoxification in Bacillus subtilis CW14. Int. J. Biol. Macromol. 2021, 186, 800–810. [Google Scholar] [CrossRef] [PubMed]
- Mwabulili, F.; Xie, Y.; Li, Q.; Sun, S.; Yang, Y.; Ma, W. Research progress of ochratoxin a bio-detoxification. Toxicon 2023, 222, 107005. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Wu, Z.; Wu, S.; Dai, Y.; Sun, C. Degradation of ochratoxin A by Bacillus amyloliquefaciens ASAG1. Food Addit. Contam. Part A 2015, 32, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Valero, A.; Marín, S.; Ramos, A.J.; Sanchis, V. Effect of preharvest fungicides and interacting fungi on Aspergillus carbonarius growth and ochratoxin A synthesis in dehydrating grapes. Lett. Appl. Microbiol. 2007, 45, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Thomidis, T.; Farmakis, A.; Prodromou, I.; Paresidou, M. Pre- and post-storage applications of rhamnolipids and cyprodinil + fludioxonil against Aspergillus section Nigri population and ochratoxin A contamination of grapes. Can. J. Plant Pathol. 2024, 46, 456–464. [Google Scholar] [CrossRef]
- Bellí, N.; Marín, S.; Sanchis, V.; Ramos, A.J. Impact of fungicides on Aspergillus carbonarius growth and ochratoxin A production on synthetic grape-like medium and on grapes. Food Addit. Contam. 2006, 23, 1021–1029. [Google Scholar] [CrossRef]
- Curto, R.L.; Pellicano, T.; Vilasi, F.; Munafo, P.; Dugo, G. Ochratoxin A occurrence in experimental wines in relationship with different pesticide treatments on grapes. Food Chem. 2004, 84, 71–75. [Google Scholar] [CrossRef]
- Malandrakis, A.A.; Vattis, K.N.; Markoglou, A.N.; Karaoglanidis, G.S. Characterization of boscalid-resistance conferring mutations in the SdhB subunit of respiratory complex II and impact on fitness and mycotoxin production in Penicillium expansum laboratory strains. Pestic. Biochem. Physiol. 2017, 138, 97–103. [Google Scholar] [CrossRef]
- Masiello, M.; Somma, S.; Haidukowski, M.; Logrieco, A.F.; Moretti, A. Genetic polymorphisms associated to SDHI fungicides resistance in selected Aspergillus flavus strains and relation with aflatoxin production. Int. J. Food Microbiol. 2020, 334, 108799. [Google Scholar] [CrossRef]
- Testempasis, S.I.; Karaoglanidis, G.S. Resistance of Black Aspergilli Species from Grape Vineyards to SDHI, QoI, DMI, and Phenylpyrrole Fungicides. J. Fungi 2023, 9, 221. [Google Scholar] [CrossRef]
- Zouhair, S.; Qjidaa, S.; Selouane, A.; Bouya, D.; Decock, C.; Bouseta, A. Effect of five fungicides on growth and ochratoxin A production by two Aspergillus carbonarius and Aspergillus niger isolated from Moroccan grapes. South Asian J. Exp. Biol. 2014, 4, 118–126. [Google Scholar]
- Terra, M.F.; Lira, N.d.A.; Passamani, F.R.F.; Santiago, W.D.; das Graças Cardoso, M.; Batista, L.R. Effect of Fungicides on Growth and Ochratoxin A Production by Aspergillus carbonarius from Brazilian Wine Grapes. J. Food Prot. 2016, 79, 1508–1516. [Google Scholar] [CrossRef] [PubMed]
- de Felice, D.V.; Solfrizzo, M.; De Curtis, F.; Lima, G.; Visconti, A.; Castoria, R. Strains of Aureobasidium pullulans Can Lower Ochratoxin A Contamination in Wine Grapes. Phytopathology 2008, 98, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Bozoudi, D.; Tsaltas, D. The Multiple and Versatile Roles of Aureobasidium pullulans in the Vitivinicultural Sector. Fermentation 2018, 4, 85. [Google Scholar] [CrossRef]
- Asad, S.A. Mechanisms of action and biocontrol potential of Trichoderma against fungal plant diseases—A review. Ecol. Complex. 2022, 49, 100978. [Google Scholar] [CrossRef]
- Mukhopadhyay, R.; Kumar, D. Trichoderma: A beneficial antifungal agent and insights into its mechanism of biocontrol potential. Egypt. J. Biol. Pest. Control 2020, 30, 133. [Google Scholar] [CrossRef]
- Barberis, C.L.; Pena, G.; Carranza, C.; Magnoli, C.E. Effect of indigenous mycobiota on ochratoxin A production by Aspergillus carbonarius isolated from soil. Mycotoxin Res. 2014, 30, 1–8. [Google Scholar] [CrossRef]
- Venkatesh, N.; Keller, N.P. Mycotoxins in Conversation With Bacteria and Fungi. Front. Microbiol. 2019, 10, 403. [Google Scholar]
- Sharma, R.R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control 2009, 50, 205–221. [Google Scholar]
- El Khoury, R.; Atoui, A.; Verheecke, C.; Maroun, R.; El Khoury, A.; Mathieu, F. Essential Oils Modulate Gene Expression and Ochratoxin A Production in Aspergillus carbonarius. Toxins 2016, 8, 242. [Google Scholar] [CrossRef]
- Hua, H.; Xing, F.; Selvaraj, J.N.; Wang, Y.; Zhao, Y.; Zhou, L.; Liu, X.; Liu, Y. Inhibitory Effect of Essential Oils on Aspergillus ochraceus Growth and Ochratoxin A Production. PLoS ONE 2014, 9, e108285. [Google Scholar]
- Minardi, P.; Pizzamiglio, V.; Mucini, S.; Piva, A. Effects of Thymol, Carvacrol and Some Weak Organic Acids on Growth and Ochratoxin A Production by the Food Spoilage Aspergillus ochraceus. Ital. J. Agron. 2008, 3, 457–458. [Google Scholar]
- Minardi, P.; Pizzamiglio, V.; Mucini, S.; Piva, A. Aspergillus ochraceus growth and ocratoxin A production inhibited by natural phenolic compounds and weak organic acids. In Proceedings of the ISM Conference on Worldwide Mycotoxin Reduction in Food and Feed Chains, Tulin, Austria, 9–11 September 2009; Department for Agrobiotechnology, IFA: Tulln, Austria, 2009; p. 86. [Google Scholar]
- De Curtis, F.; de Felice, D.V.; Ianiri, G.; De Cicco, V.; Castoria, R. Environmental factors affect the activity of biocontrol agents against ochratoxigenic Aspergillus carbonarius on wine grape. Int. J. Food Microbiol. 2012, 159, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Kizis, D.; Natskoulis, P.; Nychas, G.J.E.; Panagou, E.Z. Biodiversity and ITS-RFLP Characterisation of Aspergillus Section Nigri Isolates in Grapes from Four. PLoS ONE 2014, 9, e93923. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Boscalid | Azoxystrobin | Fludioxonil | Cyprodinil | Pyrimethanil | Tebuconazole | Chlorothalonil |
|---|---|---|---|---|---|---|---|
| AASN54 | 0.1934 | 0.1111 | <0.1 | 0.0128 | 0.0433 | 0.8751 | 0.1937 |
| BASN31 | 0.1237 | <0.025 | <0.1 | 0.0111 | 0.0430 | 0.2999 | 1.0189 |
| CASN16 | 0.2073 | 0.0394 | <0.1 | <0.01 | 0.0457 | 0.3260 | 0.1398 |
| DASN43 | 0.1487 | 0.0453 | <0.1 | 0.0107 | 0.0845 | 0.4344 | 0.6098 |
| EASN14 | >5 | 0.0740 | <0.1 | <0.01 | 0.1862 | 0.2619 | 0.6339 |
| AC33 | 3.3275 | 0.0936 | <0.1 | 0.0102 | 0.0414 | 0.2718 | 0.0833 |
| AC29 | <0.1 | <0.025 | <0.1 | <0.01 | 0.0426 | 0.1308 | 0.1608 |
| AC55 | 2.7692 | <0.025 | <0.1 | <0.01 | 0.0398 | 0.0862 | 0.0283 |
| AC60 | 0.1580 | 0.0298 | <0.1 | <0.01 | 0.0475 | 0.3858 | 0.0412 |
| Chemical PPPs | Active Substances | Maximum Certified Dose | Supplier |
|---|---|---|---|
| Cantus® | Boscalid | 12 gr L−1 | BASF Hellas S.A., Athens, Greece |
| Chorus® | Cyprodinil | 5 gr L−1 | Syngenta Hellas S.A., Athens, Greece |
| Geoxe® | Fludioxonil | 1 gr L−1 | Syngenta Hellas S.A., Athens, Greece |
| Quadris® | Azoxystrobin | 1 mL L−1 | Syngenta Hellas S.A., Athens, Greece |
| Scala® | Pyrimethanil | 2 mL L−1 | Bayer Hellas A.G., Athens, Greece |
| Switch® | Fludioxonil + cyprodinil | 1 gr L−1 | Syngenta Hellas S.A., Athens, Greece |
| Biological PPPs/ Biocontrol Agents | Active Substances | Maximum Certified Dose | Supplier |
|---|---|---|---|
| Botector® | Aureobasidium pullulans DSM 14940/DSM 14941 | 1 gr L−1 | Elanco Hellas S.A., Athens, Greece |
| EM® | Effective microorganisms: mixture of beneficial microorganisms | 0.2 mL L−1 | EMRO, Japan, Okinawa, Japan |
| Mevalone® | Eugenol, geraniol and thymol | 4 mL L−1 | K & N Efthymiadis S.A., Athens, Greece |
| Mycostop® | Streptomyces griseoviridis strain K61 | 1 gr L−1 | Verdera Oy Finland, Espoo, Finland |
| Remedier® | Trichoderma asperellum ICC012 Trichoderma gamsii ICC080 | 2.5 gr L−1 | Agrology S.A., Thessaloniki, Greece |
| Serenade Max® | Bacillus amyloliquefaciens strain QST 713 | 4 gr L−1 | Bayer Hellas A.G., Athens, Greece |
| Trianum® | Trichoderma harzianum T22 | 0.3 g L−1 | Koppert B.V. Hellas, Athens, Greece |
| Tusal® | Trichoderma asperellum T25 Trichoderma atroviride T11 | 1 gr L−1 | K & N Efthymiadis S.A., Athens, Greece |
| Vacciplant® | Laminarin | 2 mL L−1 | Alfa Agricultural Supplies S.A., Athens, Greece |
| Y1 | Aureobasidium pullulans | 107 cfu mL−1 | Plant Pathology—AUA, Athens, Greece |
| Mix Yeasts | Candida railenensis Z8 Aureobasidium pullulans Z31 Aureobasidium pullulans Y1 Rhodotorula mucilaginosa SR8 Debaryomyces hansenii VOL3 | 107 cfu mL−1 | Plant Pathology—AUA, Athens, Greece |
| Active Substances | Applied Doses (μg mL−1) | |||||
|---|---|---|---|---|---|---|
| Azoxystrobin | 0.025 | 0.05 | 0.1 | 0.5 | 1 | 5 |
| Boscalid | 0.1 | 0.5 | 1 | 5 | ||
| Chlorothalonil | 0.01 | 0.1 | 0.5 | 1 | 5 | |
| Cyprodinil | 0.01 | 0.1 | 1 | 10 | 50 | |
| Fludioxonil | 0.1 | 1 | 10 | 50 | ||
| Pyrimethanil | 0.01 | 0.025 | 0.05 | 0.1 | 0.5 | 1 |
| Tebuconazole | 0.05 | 0.1 | 0.5 | 1 | 5 | |
| Year of Isolation | Strain Code | Origin | Reference * |
|---|---|---|---|
| 2015 | AASN54 | Stimagka | PP AUA |
| 2016 | BASN31 | Stimagka | PP AUA |
| 2017 | CASN16 | Spata | PP AUA |
| 2018 | DASN43 | Spata | PP AUA |
| 2019 | EASN14 | Spata | PP AUA |
| 2014 | AC29 | Crete | [57] |
| 2014 | AC33 | Attica | [57] |
| 2014 | AC55 | Peloponnese | [57] |
| 2014 | AC60 | Macedonia | [57] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iliadi, M.K.; Varveri, M.; Tsitsigiannis, D.I. Biological and Chemical Management of Aspergillus carbonarius and Ochratoxin A in Vineyards. Toxins 2024, 16, 527. https://doi.org/10.3390/toxins16120527
Iliadi MK, Varveri M, Tsitsigiannis DI. Biological and Chemical Management of Aspergillus carbonarius and Ochratoxin A in Vineyards. Toxins. 2024; 16(12):527. https://doi.org/10.3390/toxins16120527
Chicago/Turabian StyleIliadi, Maria K., Maria Varveri, and Dimitrios I. Tsitsigiannis. 2024. "Biological and Chemical Management of Aspergillus carbonarius and Ochratoxin A in Vineyards" Toxins 16, no. 12: 527. https://doi.org/10.3390/toxins16120527
APA StyleIliadi, M. K., Varveri, M., & Tsitsigiannis, D. I. (2024). Biological and Chemical Management of Aspergillus carbonarius and Ochratoxin A in Vineyards. Toxins, 16(12), 527. https://doi.org/10.3390/toxins16120527