Iridoid Glycosides Isolated from Bellardia trixago Identified as Inhibitors of Orobanche cumana Radicle Growth


 ,
,  , ,
, ,  , ,
, ,  and
and 
Abstract
:1. Introduction
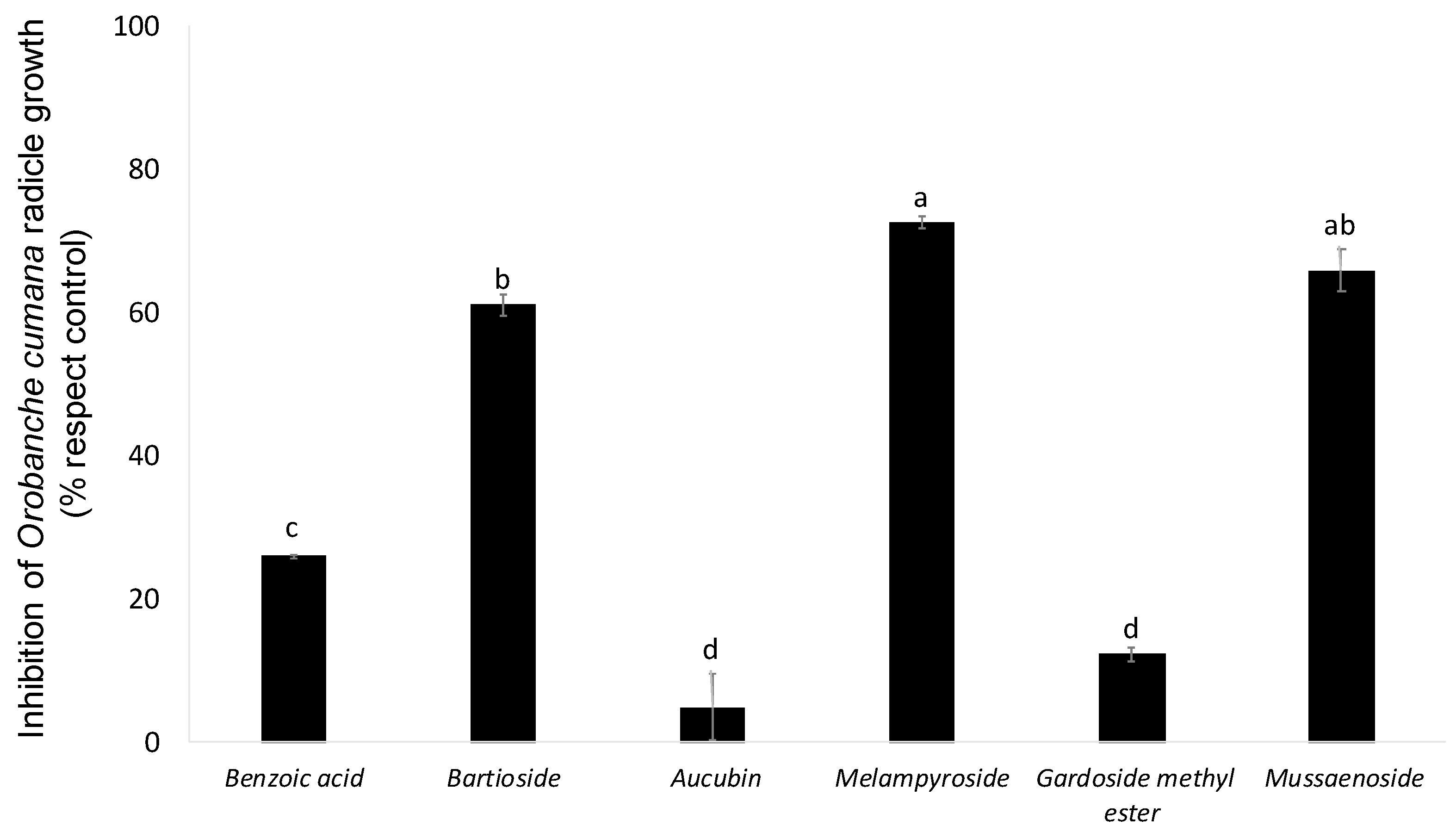
2. Results and Discussion
3. Conclusions
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Extractions of Bellardia trixago Organs
4.4. Isolation and Identification of Metabolites from Bellardia trixago Green Organs of the White-Flowered Population
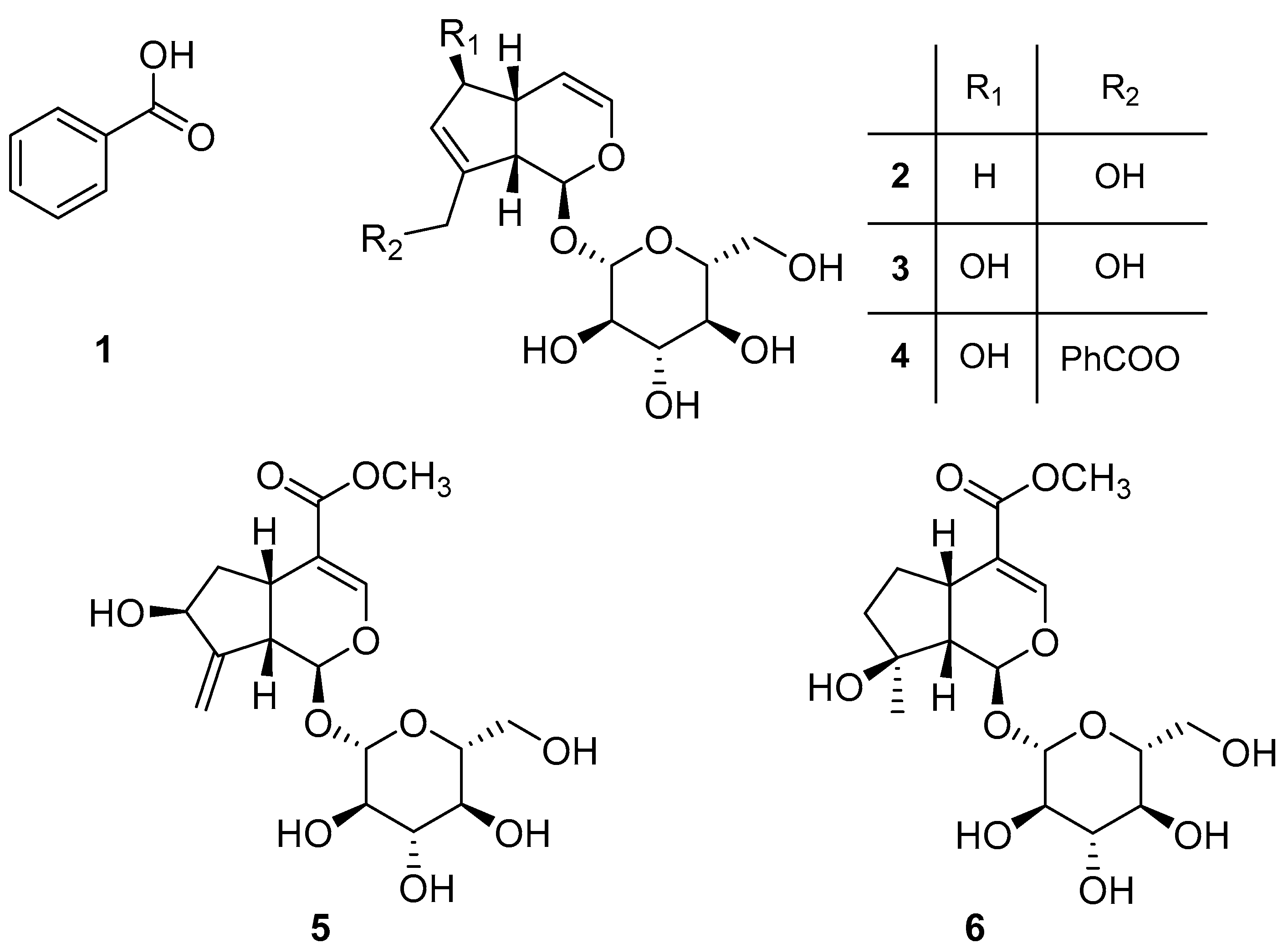
- Melampyroside (4): [α]D22-69.6 (c 0.79, MeOH) [lit. [27]: [α]D26-52.9 (c 0.31, MeOH)]. 1H and 13C NMR spectra (Figures S4 and S5) were in agreement with data previously reported [11,49], while its NOESY spectrum is reported in Figure S6. ESI MS (+) m/z: 451 [M + H]+.
4.5. Bioactivity on Parasitic Weed Seeds
4.6. Molecular Modelling
4.7. Ecotoxicity Analysis on Melampyroside
4.8. Data Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parker, C.; Riches, C.R. Parasitic Weeds of the World: Biology and Control; CAB International: Wallingford, UK, 1993; ISBN 9780851988733. [Google Scholar]
- Westwood, J.H.; Yoder, J.I.; Timko, M.P.; dePamphilis, C.W. The evolution of parasitism in plants. Trends Plant Sci. 2010, 15, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Press, M.C.; Parsons, A.N.; Mackay, A.W.; Vincent, C.A.; Cochrane, V.; Seel, W.E. Gas exchange characteristics and nitrogen relations of two Mediterranean root hemiparasites: Bartsia trixago and Parentucellia viscosa. Oecologia 1993, 95, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Uribe-Convers, S.; Tank, D.C. Phylogenetic revision of the genus Bartsia (Orobanchaceae): Disjunct distributions correlate to independent lineages. Syst. Bot. 2016, 41, 672–684. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Sillero, J.C.; Rubiales, D. Resistance to broomrape species (Orobanche spp.) in common vetch (Vicia sativa L.). Crop. Prot. 2009, 28, 7–12. [Google Scholar] [CrossRef]
- Molinero-Ruiz, L.; Delavault, P.; Pérez-Vich, B.; Pacureanu-Joita, M.; Bulos, M.; Altieri, E.; Domínguez, J. History of the race structure of Orobanche cumana and the breeding of sunflower for resistance to this parasitic weed: A review. Span. J. Agric. Res. 2015, 13, e10R01. [Google Scholar] [CrossRef]
- Parker, C. Observations on the current status of Orobanche and Striga problems worldwide. Pest Manag. Sci. 2009, 65, 453–459. [Google Scholar] [CrossRef]
- Eizenberg, H.; Hershenhorn, J.; Ephrath, J.E. Factors affecting the efficacy of Orobanche cumana chemical control in sunflower. Weed Res. 2008, 49, 308–315. [Google Scholar] [CrossRef]
- Joel, D.M.; Hershenhorn, J.; Eizenberg, H.; Aly, R.; Ejeta, G.; Rich, P.J.; Ransom, J.K.; Sauerborn, J.; Rubiales, D. Biology and management of weedy root parasites. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons: New York, NY, USA, 2007; Volume 33, pp. 267–350. [Google Scholar]
- Labrousse, P.; Arnaud, M.C.; Griveau, Y.; Fer, A.; Thalouarn, P. Analysis of resistance criteria of sunflower recombined inbred lines against Orobanche cumana Wallr. Crop Prot. 2004, 23, 407–413. [Google Scholar] [CrossRef]
- Aly, R.; Goldwasser, Y.; Eizenber, H.; Hershenhorn, J.; Golan, S.; Kleifeld, Y. Broomrape (Orobanche cumana) control in sunflower (Helianthus annuus) with imazapic. Weed Technol. 2009, 15, 306–3009. [Google Scholar] [CrossRef]
- Lerner, F.; Pfenning, M.; Picard, L.; Lerchl, J.; Hollenbach, E. Prohesadione calcium is herbicidal to the sunflower root parasite Orobanche cumana. Pest Manag. Sci. 2021, 77, 1893–1902. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Delavault, P.; Timko, M.P. Management of infection by parasitic weeds: A Review. Plants 2020, 9, 1184. [Google Scholar] [CrossRef] [PubMed]
- Tomas-Barberan, F.A.; Cole, M.D.; Garcia-Viguera, C.; Tomas-Lorente, F.; Guirado, A. Epicuticular flavonoids from Bellardia trixago and their antifungal fully methylated derivatives. Int. J. Crude Drug Res. 1990, 28, 57–60. [Google Scholar] [CrossRef]
- Bianco, A.; Guiso, M.; Iavarone, C.; Trogolo, C. Iridoids. XX. Bartsioside, structure and configuration. Gazz. Chim. Ital. 1976, 106, 725–732. [Google Scholar]
- Barrero, A.F.; Sánchez, J.F.; Cuenca, F.G. Dramatic variation in diterpenoids of different populations of Bellardia trixago. Phytochemistry 1988, 27, 3676–3678. [Google Scholar] [CrossRef]
- Ersöz, T.; Yalçin, F.N.; Taşdemir, D.; Sticher, O.; Çaliş, İ. Iridoid and lignan glucosides from Bellardia trixago (L.) All. Turk. J. Med. Sci. 1998, 28, 397–400. [Google Scholar]
- Pascual-Villalobos, M.J.; Robledo, A. Screening for anti-insect activity in Mediterranean plants. Ind. Crops Prod. 1998, 8, 183–194. [Google Scholar] [CrossRef]
- Formisano, C.; Rigano, D.; Senatore, F.; Simmonds, M.S.J.; Bisio, A.; Bruno, M.; Rosselli, S. Essential oil composition and antifeedant properties of Bellardia trixago (L.) All. (sin. Bartsia trixago L.) (Scrophulariaceae). Biochem. Syst. Ecol. 2008, 36, 454–457. [Google Scholar] [CrossRef]
- Weinges, K.; Ziegler, H.J. Chemie und Stereochemie der Iridoide, XIV. Aucubin und Scandosid aus Catalpol. Liebigs Ann. Chem. 1990, 1990, 715–717. [Google Scholar] [CrossRef]
- Tietze, L.F.; Niemeyer, U.; Marx, P.; Glüsenkamp, K.H.; Schwenen, L. Iridoide—XIII: Bestimmung der absoluten konfiguration und konformation isomerer iridoidglycoside mit hilfe chiroptischer methoden. Tetrahedron 1980, 36, 735–739. [Google Scholar] [CrossRef]
- Venditti, A.; Ballero, M.; Serafini, M.; Bianco, A. Polar compounds from Parentucellia viscosa (L.) Caruel from Sardinia. Nat. Prod. Res. 2015, 29, 602–606. [Google Scholar] [CrossRef]
- Cuendet, M.; Potterat, O.; Hostettmann, K. Iridoid glucosides, phenylpropanoid derivatives and flavanoids from Bartsia alpina. Pharm. Biol. 1999, 37, 318–320. [Google Scholar] [CrossRef]
- Kirmizibekmez, H.; Atay, I.; Kaiser, M.; Brun, R.; Cartagena, M.M.; Carballeira, N.M.; Yesilada, E.; Tasdemir, D. Antiprotozoal activity of Melampyrum arvense and its metabolites. Phytother. Res. 2011, 25, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Takeda, Y.; Tamura, K.; Matsumoto, T.; Terao, H.; Tabata, M.; Fujita, T.; Honda, G.; Sezik, E.; Yesiladat, E. 6′-O-benzoylshanzhiside methyl ester from Rhinanthus angustifolius subsp. grandiflorus. Phytochemistry 1993, 33, 623–625. [Google Scholar] [CrossRef]
- Bianco, A.; Bolli, D.; Passacantilli, P. 6-O-β-Glucopyranosylaucubin, a new irodoid from Odontites verna. Planta Med. 1982, 44, 97–99. [Google Scholar] [CrossRef]
- Takeda, Y.; Fujita, T. Iridoid glucosides of Melampyrum laxum. Planta Med. 1981, 41, 192–194. [Google Scholar] [CrossRef] [PubMed]
- Damtoft, S.; Hansen, S.B.; Jacobsen, B.; Jensen, S.R.; Nielsen, B.J. Iridoid glucosides from Melampyrum. Phytochemistry 1984, 23, 2387–2389. [Google Scholar] [CrossRef]
- Bianco, A.; Passacantilli, P.; Righi, G.; Nicoletti, M. Iridoid glucosides from Parentucellia viscosa. Phytochemistry 1985, 24, 1843–1845. [Google Scholar] [CrossRef]
- Petitto, V.; Serafini, M.; Ballero, M.; Foddai, S.; Stanzione, A.; Nicoletti, M. Iridoids from Euphrasia genargentea, a rare Sardinian endemism. Nat. Prod. Res. 2009, 23, 431–435. [Google Scholar] [CrossRef]
- Khanh, T.D.; Anh, L.H.; Nghia, L.T.; Trung, K.H.; Hien, P.B.; Trung, D.M.; Xuan, T.D. Allelopathic responses of rice seedlings under some different stresses. Plants 2018, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Aparicio, M.; Cimmino, A.; Evidente, A.; Rubiales, D. Inhibition of Orobanche crenata seed germination and radicle growth by allelochemicals identified in cereals. J. Agric. Food Chem. 2013, 61, 9797–9803. [Google Scholar] [CrossRef]
- Zhang, W.; Lu, L.-Y.; Hu, L.-Y.; Cao, W.; Sun, K.; Sun, Q.-B.; Siddikee, A.; Shi, R.-H.; Dai, C.-C. Evidence for the involvement of auxin, ethylene and ROS signaling during primary root inhibition of Arabidopsis by the allelochemical benzoic acid. Plant Cell Physiol. 2018, 59, 1889–1904. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Chen, Y.; Chen, J.; Zhang, Y.; Zhang, T.; He, H. Comparison of allelopathic effects of two typical invasive plants: Mikania micrantha and Ipomoea cairica in Hainan island. Sci. Rep. 2020, 10, 11332. [Google Scholar] [CrossRef] [PubMed]
- Baziramakenga, R.; Leroux, G.D.; Simard, R.R. Effects of benzoic and cinnamic acids on membrane permeability of soybean roots. J. Chem. Ecol. 1995, 21, 1271–1285. [Google Scholar] [CrossRef]
- Earl, D.F. Method of Treating Plants. U.S. Patent US2723909A, 21 August 1952. [Google Scholar]
- Pang, J. One Kind Greening Momordica Grosvenori Herbicide and Preparation Method Thereof. Chinese Patent CN107372626A, 14 August 2017. [Google Scholar]
- Jin, X.; Lin, Y.; Li, Y.; Lin, D.; Lin, J.; Lin, Z. Organic Crop Herbicide Containing Organic Acid and Mineral Oil and Preparation Method Thereof. Chinese Patent CN113925063A, 14 January 2022. [Google Scholar]
- Ahn, B.Z.; Pachaly, P. Melampyrosid, ein neus iridoid aus Melampyrum silvaticum L. Tetrahedron 1974, 30, 4049–4054. [Google Scholar] [CrossRef]
- Karrer, P.; Schmid, H. Über die Konstitution des Aucubins. Helv. Chim. Acta 1946, 29, 525–552. [Google Scholar] [CrossRef]
- Takeda, Y.; Nishimura, H.; Inouye, H. Two new iridoid glucosides from Mussaenda parviflora and Mussaenda shikokiana. Phytochemistry 1977, 16, 1401–1404. [Google Scholar] [CrossRef]
- Ji, M.; Wang, C.; Yang, T.; Meng, X.; Wang, X.; Li, M. Integrated phytochemical analysis based on UPLC–MS/MS and network pharmacology approaches to explore the effect of Odontites vulgaris Moench on rheumatoid arthritis. Front. Pharmacol. 2021, 12, 707687. [Google Scholar] [CrossRef]
- Pennacchio, M.; Syah, Y.M.; Ghisalberti, E.L.; Alexander, E. Cardioactive iridoid glycosides from Eremophila species. Phytomedicine 1997, 4, 325–330. [Google Scholar] [CrossRef]
- Çaliş, I.; Kirmizibekmez, H.; Taşdemir, D.; Ireland, C.M. Iridoid glycosides from Globularia davisiana. Chem. Pharm. Bull. 2002, 50, 678–680. [Google Scholar] [CrossRef]
- Liu, X.; Testa, B.; Fahr, A. Lipophilicity and its relationship with passive drug permeation. Pharm. Res. 2011, 28, 962–977. [Google Scholar] [CrossRef]
- Zhang, R.; Qin, X.; Kong, F.; Chen, P.; Pan, G. Improving cellular uptake of therapeutic entities through interaction with components of cell membrane. Drug Deliv. 2019, 26, 328–342. [Google Scholar] [CrossRef] [PubMed]
- Afolayan, A.J.; Ohikhena, F.U.; Wintola, O.A. Toxicity assessment of different solvent extracts of the medicinal plant, Phragmanthera capitata (Sprengel) Balle on brine shrimp (Artemia salina). Int. J. Pharmacol. 2016, 12, 701–710. [Google Scholar]
- Page, S.W. Antiparasitic drugs. In Small Animal Clinical Pharmacology; Maddison, J.E., Page, S.W., Church, D.B., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 198–260. [Google Scholar]
- European Commission (EC). Technical Guidance Document in Support of the Commission Directive 93/67/EEC on Risk Assessment for New Notified Substances and Commission Regulation (EC) No 1488/94 on Risk Assessment for Existing Substances; Parts 1–4; Office for Official Publications of the EC: Luxembourg, 1996. [Google Scholar]
- Koko, W.S.; Jentzsch, J.; Kalie, H.; Schobert, R.; Ersfeld, K.; Al Nasr, I.S.; Biersack, B. Evaluation of the antiparasitic activities of imidazol-2-ylidene-gold (I) complexes. Arch. Pharm. 2020, 353, e1900363. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.W.; Phillipson, J.D. Natural products and the development of selective antiprotozoal drugs. Phytother. Res. 1990, 4, 127–139. [Google Scholar] [CrossRef]
- Barzman, M.; Bàrberi, P.; Birch, A.N.E.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Sattin, M. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Cimmino, A.; Roscetto, E.; Masi, M.; Tuzi, A.; Radjai, I.; Gahdab, C.; Paolillo, R.; Guarino, A.; Catania, M.R.; Evidente, A. Sesquiterpene lactones from Cotula cinerea with antibiotic activity against clinical isolates of Enterococcus faecalis. Antibiotics 2021, 10, 819. [Google Scholar] [CrossRef]
- Soriano, G.; Petrillo, C.; Masi, M.; Bouafiane, M.; Khelil, A.; Tuzi, A.; Isticato, R.; Fernández-Aparicio, M.; Cimmino, A. Specialized metabolites from the allelopathic plant Retama raetam as potential biopesticides. Toxins 2022, 14, 311. [Google Scholar] [CrossRef]
- Cui, L.-Q.; Liu, K.; Zhang, C. Effective oxidation of benzylic and alkane C–H bonds catalyzed by sodium o-iodobenzenesulfonate with Oxone as a terminal oxidant under phase-transfer conditions. Org. Biomol. Chem. 2011, 9, 2258–2265. [Google Scholar] [CrossRef]
- Delicato, A.; Masi, M.; de Lara, F.; Rubiales, D.; Paolillo, I.; Lucci, V.; Falco, G.; Calabro, V.; Evidente, A. In vitro characterization of iridoid and phenylethanoid glycosides from Cistanche phelypaea for nutraceutical and pharmacological applications. Phytother. Res. 2022, 1–12. [Google Scholar] [CrossRef]
- Andrzejewska-Golec, E.; Ofterdinger-Daegel, S.; Calis, I.; Światek, L. Chemotaxonomic aspects of iridoids occurring in Plantago subg. Psyllium (Plantaginaceae). Plant Syst. Evol. 1993, 185, 85–89. [Google Scholar] [CrossRef]
- Chaudhuri, R.K.; Afifi-Yazar, F.Ü.; Sticher, O.; Winkler, T. 13C NMR spectroscopy of naturally occurring iridoid glucosides and their acylated derivatives. Tetrahedron 1980, 36, 2317–2326. [Google Scholar] [CrossRef]
- Chu, H.-B.; Tan, N.-H.; Zhang, Y.-M. Chemical constituents from Pedicularis rex C. B. Clarke. Z. Naturforsch. B 2007, 62, 1465–1470. [Google Scholar] [CrossRef]
- Otsuka, H.; Watanabe, E.; Yuasa, K.; Ogimi, C.; Takushi, A.; Takeda, Y. A verbascoside iridoid glucoside conjugate from Premna corymbosa var. abtusifolia. Phytochemistry 1993, 32, 983–986. [Google Scholar] [CrossRef]
- Gardner, D.R.; Narum, J.; Zook, D.; Stermitz, F.R. New iridoid glucosides from Castilleja and Besseya: 6-Hydroxyadoxoside and 6-isovanillylcatapol. J. Nat. Prod. 1987, 50, 485–489. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Moral, A.; Kharrat, M.; Rubiales, D. Resistance against broomrapes (Orobanche and Phelipanche spp.) in faba bean (Vicia faba) based in low induction of broomrape seed germination. Euphytica 2012, 186, 897–905. [Google Scholar] [CrossRef]
- Westwood, J.H.; Foy, C.L. Influence of nitrogen on germination and early development of broomrape (Orobanche spp.). Weed Sci. 1999, 47, 2–7. [Google Scholar] [CrossRef]
- Cala, A.; Zorrilla, J.G.; Rial, C.; Molinillo, J.M.G.; Varela, R.M.; Macías, F.A. Easy access to alkoxy, amino, carbamoyl, hydroxy, and thiol derivatives of sesquiterpene lactones and evaluation of their bioactivity on parasitic weeds. J. Agric. Food Chem. 2019, 67, 10764–10773. [Google Scholar] [CrossRef]
- ISO 8692:2012; Water Quality—Fresh Water Algal Growth Inhibition Test with Unicellular Green Algae. ISO: Geneva, Switzerland, 2012.
- ISO 11269-1:2012; Soil Quality—Determination of the Effects of Pollutants on Soil Flora—Part 1: Method for the Measurement of Inhibition of Root Growth. ISO: Geneva, Switzerland, 2012.
- UNI EN ISO 6341:2013; Water Quality—Determination of the Inhibition of the Mobility of Daphnia magna Straus (Cladoc-Era, Crustacea)—Acute Toxicity Test. Ente Nazionale Italiano di Unificazione (UNI): Milan, Italy, 2013.
- ISO 11348-3:2007; Water Quality—Determination of the Inhibitory Effect of Water Samples on the Light Emission of Vibrio fischeri (Luminescent Bacteria Test)—Part 3: Method Using Freeze-Dried Bacteria. ISO: Geneva, Switzerland, 2007.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 2 | 3 | 4 | 5 | 6 | |
|---|---|---|---|---|---|---|
| CLogP | 1.885 | −1.941 | −4.028 | −1.153 | −2.133 | −1.849 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soriano, G.; Siciliano, A.; Fernández-Aparicio, M.; Cala Peralta, A.; Masi, M.; Moreno-Robles, A.; Guida, M.; Cimmino, A. Iridoid Glycosides Isolated from Bellardia trixago Identified as Inhibitors of Orobanche cumana Radicle Growth. Toxins 2022, 14, 559. https://doi.org/10.3390/toxins14080559
Soriano G, Siciliano A, Fernández-Aparicio M, Cala Peralta A, Masi M, Moreno-Robles A, Guida M, Cimmino A. Iridoid Glycosides Isolated from Bellardia trixago Identified as Inhibitors of Orobanche cumana Radicle Growth. Toxins. 2022; 14(8):559. https://doi.org/10.3390/toxins14080559
Chicago/Turabian StyleSoriano, Gabriele, Antonietta Siciliano, Mónica Fernández-Aparicio, Antonio Cala Peralta, Marco Masi, Antonio Moreno-Robles, Marco Guida, and Alessio Cimmino. 2022. "Iridoid Glycosides Isolated from Bellardia trixago Identified as Inhibitors of Orobanche cumana Radicle Growth" Toxins 14, no. 8: 559. https://doi.org/10.3390/toxins14080559
APA StyleSoriano, G., Siciliano, A., Fernández-Aparicio, M., Cala Peralta, A., Masi, M., Moreno-Robles, A., Guida, M., & Cimmino, A. (2022). Iridoid Glycosides Isolated from Bellardia trixago Identified as Inhibitors of Orobanche cumana Radicle Growth. Toxins, 14(8), 559. https://doi.org/10.3390/toxins14080559