In-Vitro Application of a Qatari Burkholderia cepacia strain (QBC03) in the Biocontrol of Mycotoxigenic Fungi and in the Reduction of Ochratoxin A biosynthesis by Aspergillus carbonarius




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Determination of the Spectrum of Burkholderia Cepacia QBC03 Antifungal Activity
2.2. Antifungal Activity of QBC03’s Culture Extract in Solid Media
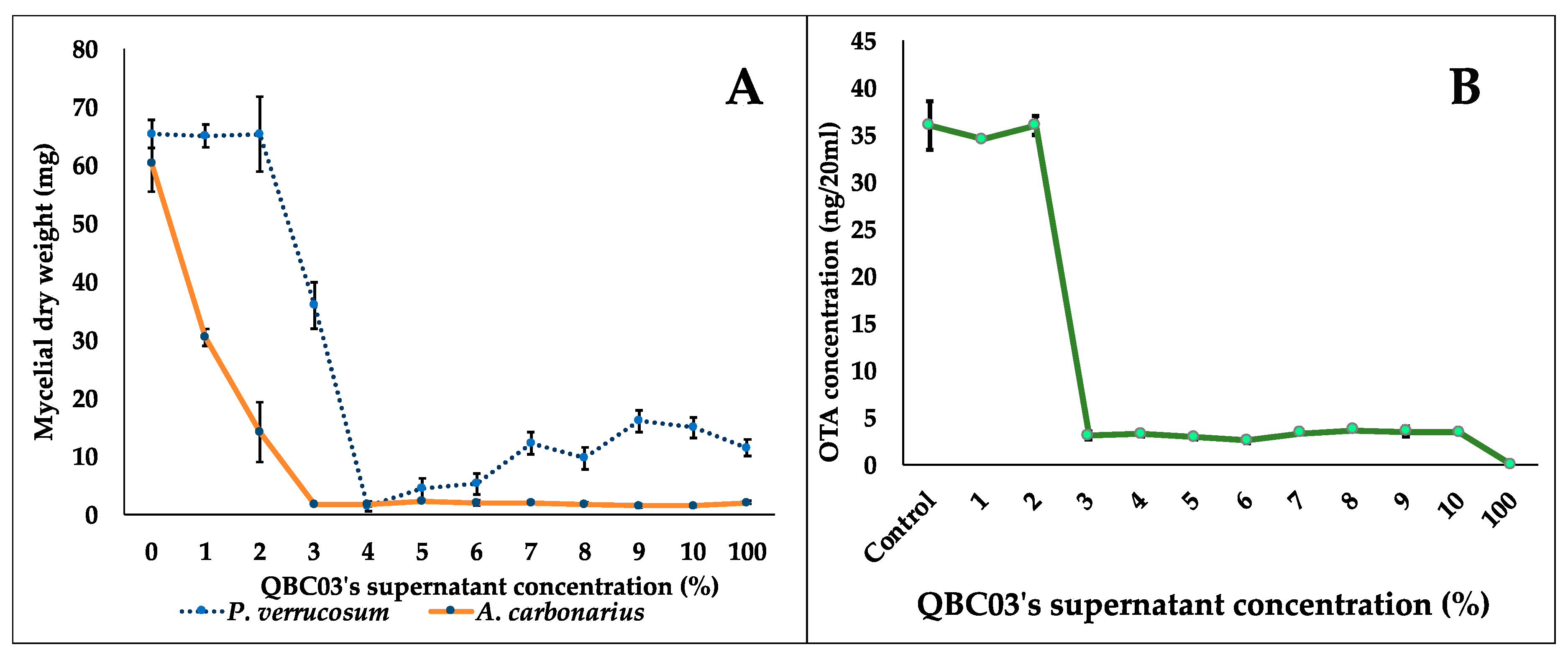
2.3. Effect of QBC03’s Metabolites on the Fungal Biomass
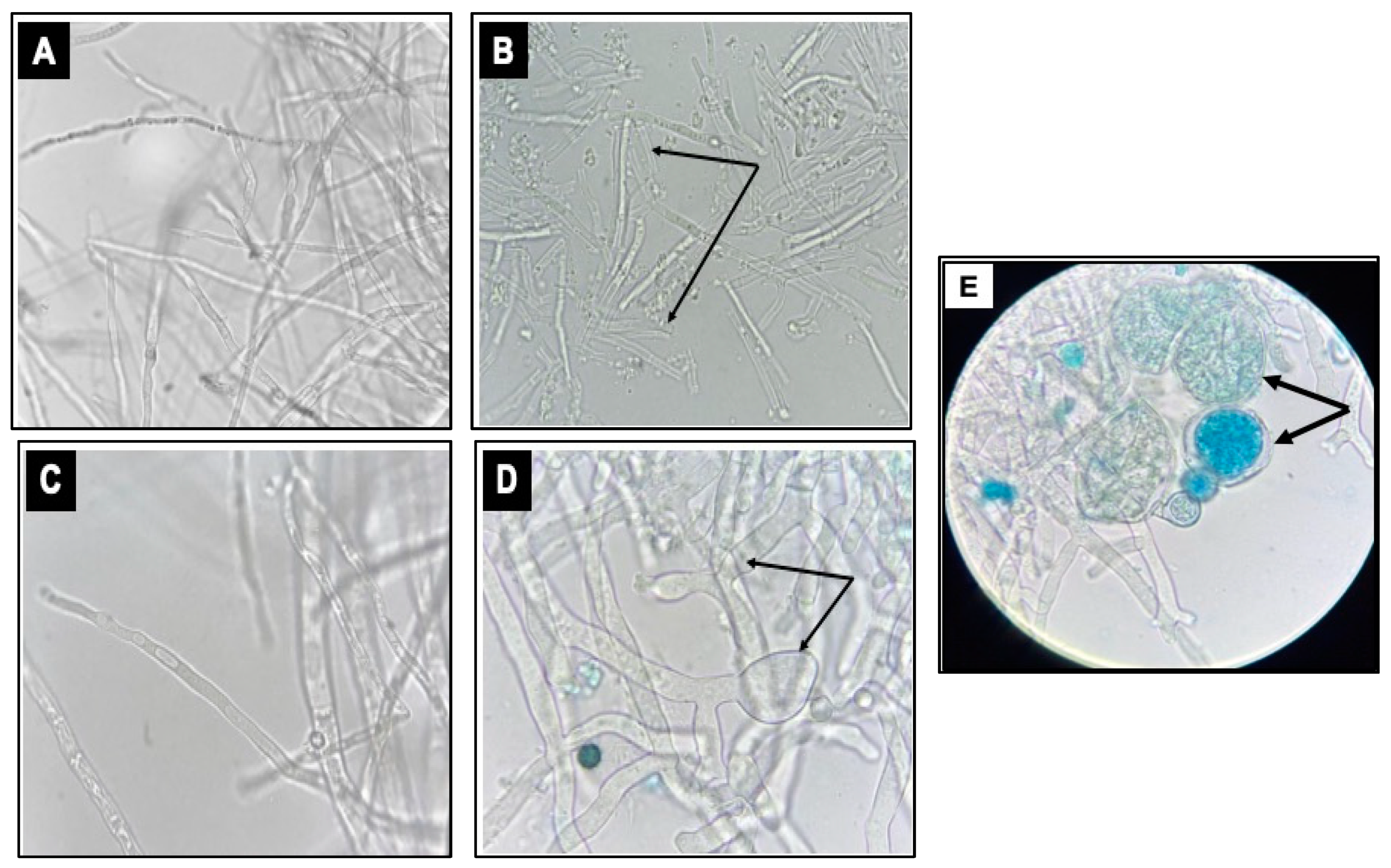
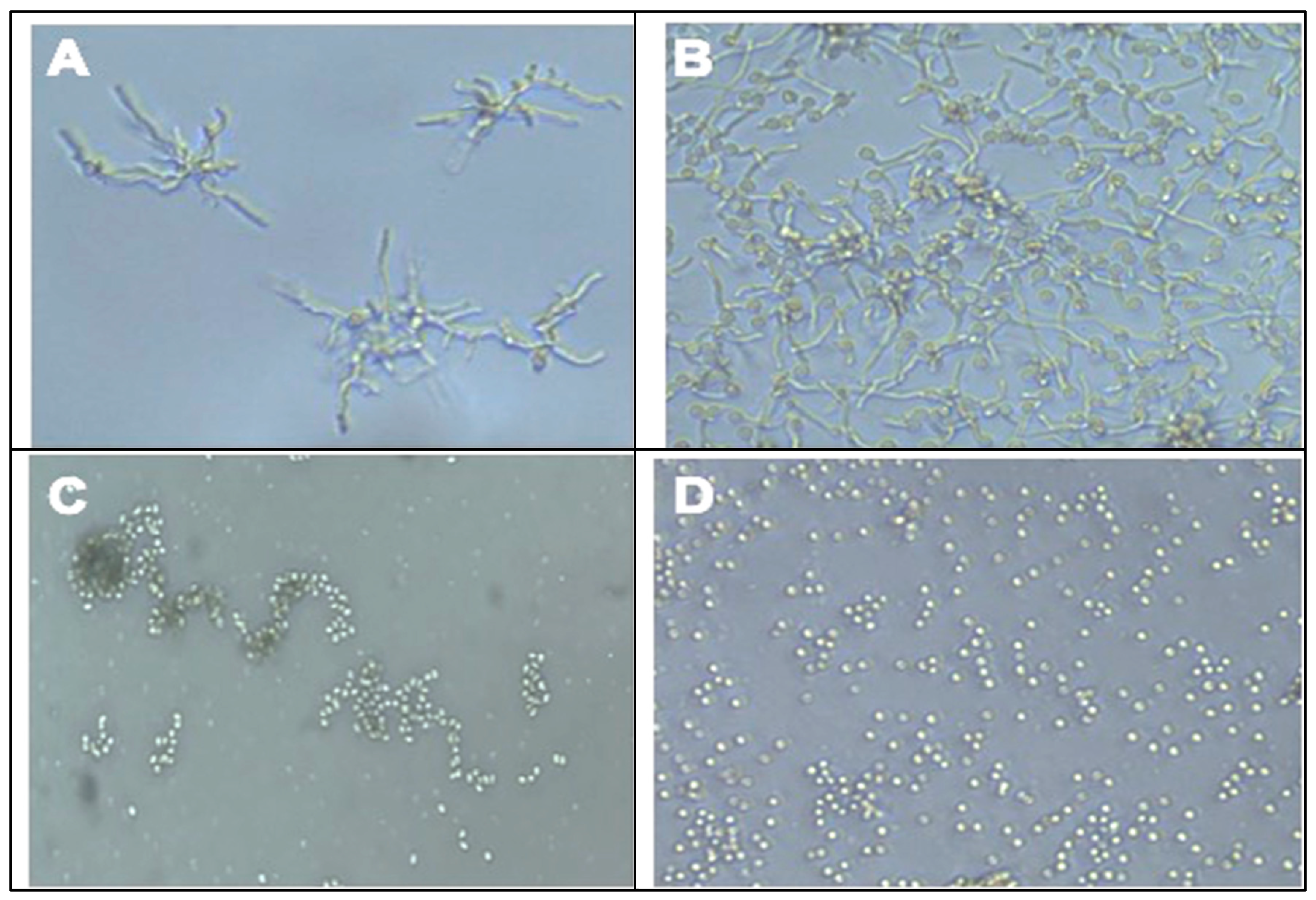
2.4. Effect of QBC03 Culture Extract on Fungal Spore Germination
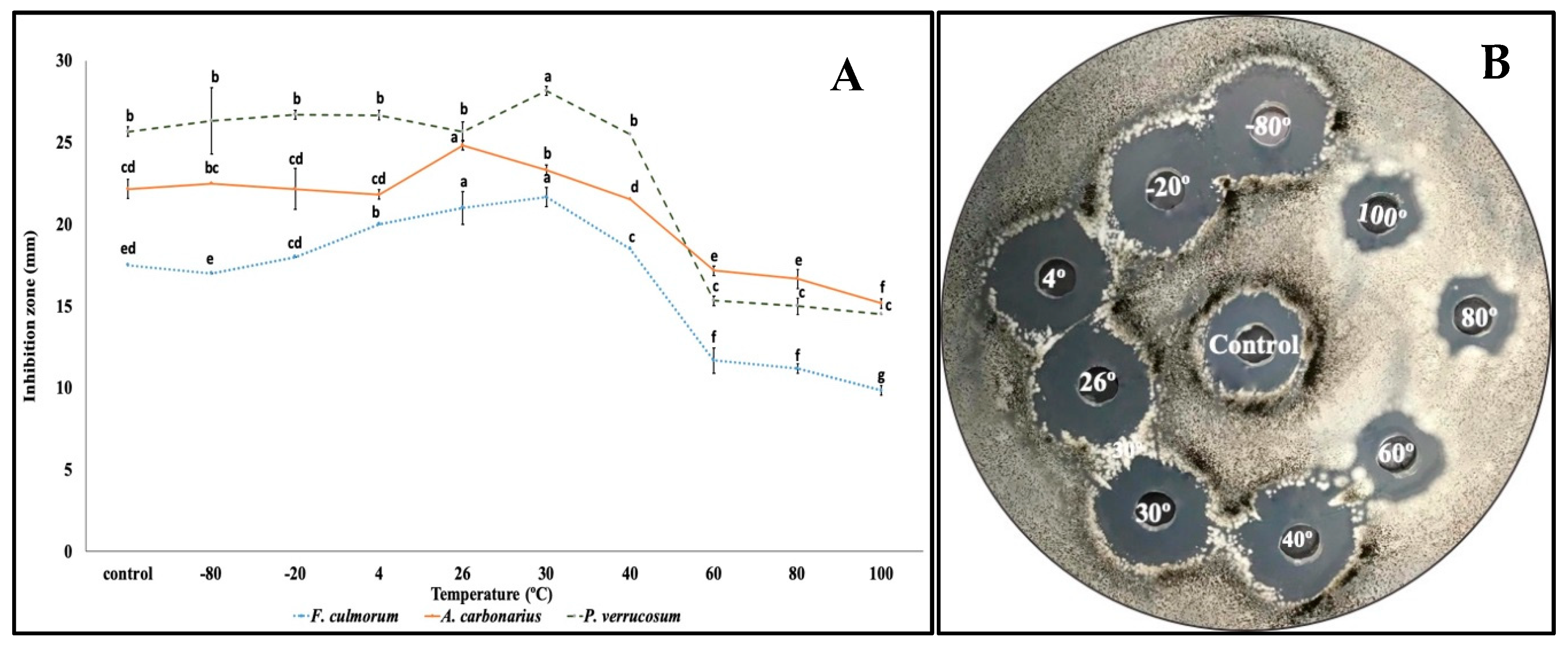
2.5. Study on the Thermostability of QBC03 Antifungal Compounds
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Microbial Strains
5.2. Screening the Antifungal Activity of QBC03
5.3. Investigation of QBC03 Culture Extract on the Fungal Growth in Solid Media
5.4. Investigation of the Effect of QBC03 Culture Extract on Fungal Biomass and Mycotoxin Synthesis
5.5. Evaluation of the Effect of QBC03’s Antifungal Compounds on Fungal Spore Germination
5.6. Influence of Temperature on the Stability of QBC03 Antifungal Compounds
5.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abbas, H.K. (Ed.) Aflatoxin and Food Safety; Taylor and Frances Group: Philadelphia, PA, USA, 2005. [Google Scholar]
- Rocha, M.E.B.; Freire, F.D.C.O.; Maia, F.E.F.; Guedes, M.I.F.; Rondina, D. Mycotoxins and their effects on human and animal health. Food Control 2014, 36, 159–165. [Google Scholar] [CrossRef]
- Anfossi, L.; Giovannoli, C.; Baggiani, C. Mycotoxin detection. Curr. Opin. Biotechnol. 2016, 37, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Zhao, Z.; Wang, J.; Bai, B.; Wu, A.; Yan, L.; Song, S. A simple sample pretreatment method for multi-mycotoxin determination in eggs by liquid chromatography tandem mass spectrometry. J. Chromatogr. 2015, 1417, 1–7. [Google Scholar] [CrossRef]
- Hameed, M.R.; Khan, M.Z.; Khan, A.; Javed, I. Ochratoxin induced pathological alterations in broiler chicks: Effect of dose and duration. Pak. Vet. J. 2013, 33, 145–149. [Google Scholar]
- Ul-Hassan, Z.; Khan, M.Z.; Saleemi, M.K.; Khan, A.; Javed, I.; Bhatti, S.A. Toxico-pathological effects of in-ovo inoculation of ochratoxin A (OTA) in chick embryos. Toxicol. Pathol. 2012, 40, 33–39. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Liu, F.; Wang, Q.; Selvaraj, J.N.; Xing, F.; Zhao, Y.; Liu, Y. Ochratoxin A producing fungi, biosynthetic pathway and regulatory mechanisms. Toxins 2016, 8, 83. [Google Scholar] [CrossRef]
- Hassan, Z.U.; Roda, F.A.; Migheli, Q.; Samir, J. Detection of toxigenic mycobiota and mycotoxins in cereal feed market. Food Control 2018, 84, 389–394. [Google Scholar] [CrossRef]
- Hassan, Z.U.; Roda, F.A.; Fathy, A.A.; Saeed, A.M.; Migheli, Q.; Samir, J. Co-occurrence of mycotoxins in commercial formula milk and cereal-based baby food. Food Addit. Contam. Part B 2018, 11, 191–197. [Google Scholar] [CrossRef]
- De Jesus, C.L.; Bartley, A.; Welch, A.Z.; Berry, J.P. High incidence and levels of ochratoxin A in wines sourced from the United States. Toxins 2017, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.H.; Bingle, L.E. Comparative ochratoxin toxicity: A review of the available data. Toxins 2015, 7, 4253–4582. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.L.; Cendoya, E.; Nichea, M.J.; Zachetti, V.G.L.; Chulze, S.N. Impact of toxigenic fungi and mycotoxins in chickpea: A review. Curr. Opin. Food Sci. 2018, 23, 32–37. [Google Scholar] [CrossRef]
- Milani, J.M. Ecological conditions affecting mycotoxin production in cereals: A review. Veterinarni Medicina 2013, 58, 405–411. [Google Scholar] [CrossRef]
- Mannaa, M.; Kim, K.D. Influence of temperature and water activity on deleterious fungi and mycotoxin producing during grain storage. Mycobiology 2017, 45, 240–254. [Google Scholar] [CrossRef] [PubMed]
- Kabak, B.; Dobson, A.D.W.; Var, I. Strategies to prevent mycotoxin contamination of food and animal feed: A review. Crit. Rev. Food Sci. Nutr. 2006, 46, 593–619. [Google Scholar] [CrossRef] [PubMed]
- Tola, M.; Kebede, B. Occurrence, importance and control of mycotoxins: A review. Cogent Food Agric. 2016, 2. [Google Scholar] [CrossRef]
- Mannaa, M.; Oh, J.Y.; Kim, K.D. Biocontrol activity of volatile-producing Bacillus megaterium and Pseudomonas protegens against Aspergillus flavus and aflatoxin production on stored rice grains. Mycobiology 2017, 45, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Mannaa, M.; Oh, J.Y.; Kim, K.D. Microbe-mediated control of Aspergillus flavus in stored rice grains with a focus on aflatoxin inhibition and biodegradation. Ann. Appl. Biol. 2017, 171, 376–392. [Google Scholar] [CrossRef]
- Hassan, Z.U.; Al-Thani, R.; Alnaimi, H.; Migheli, Q.; Jaoua, S. Investigation and application of Bacillus licheniformis volatile compounds for the biological control of toxigenic Aspergillus and Penicillium spp. ACS Omega 2019, 4, 17186–17193. [Google Scholar] [CrossRef]
- Zeidan, R.; Ul-Hassan, Z.; Al-Thani, R.; Balmas, V.; Jaoua, S. Application of low-fermenting yeast Lachancea thermotolerans for the control of toxigenic fungi Aspergillus parasiticus, Penicillium verrucosum and Fusarium graminearum and their mycotoxins. Toxins 2018, 10, 242. [Google Scholar] [CrossRef]
- Abdallah, M.F.; Ameye, M.; De Saeger, S.; Audenaert, K.; Haesaert, G. Biological control of mycotoxigenic fungi and their toxins: An update for the pre-harvest approach. In Fungi and Mycotoxins: Their Occurrence, Impact on Health and the Economy as Well as Pre- and Postharvest Management Strategies; Berka Njobeh, P., Ed.; IntechOpen Press: London, UK, 2018; pp. 1–31. [Google Scholar] [CrossRef]
- Nagórska, K.; Bikowski, M.; Obuchowski, M. Multicellular behaviour and production of a wide variety of toxic substances support usage of Bacillus subtilis as a powerful biocontrol agent. Acta Biochim. Polon. 2007, 54, 495–508. [Google Scholar] [CrossRef]
- Mannaa, M.; Kim, K.D. Biocontrol activity of volatile-producing Bacillus megaterium and Pseudomonas protegens against Aspergillus and Penicillium spp. Predominant in stored rice grains: Study II. Mycobiology 2018, 46, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Kilani-Feki, O.; Culioli, G.; Ortalo-Magné, A.; Zouari, N.; Blache, Y.; Jaoua, S. Environmental Burkholderia cepacia strain Cs5 acting by two analogous alkyl-quinolones and a didecyl-phthalate against a broad spectrum of phytopathogens fungi. Curr. Microbiol. 2011, 62, 1490–1495. [Google Scholar] [CrossRef] [PubMed]
- Petatán-Sagahón, I.; Anducho-Reyes, M.A.; Silva-Rojas, H.V.; Arana-Cuenca, A.; Tellez-Jurado, A.; Cárdenas-Álvarez, I.O.; Mercado-Flores, Y. Isolation of bacteria with antifungal activity against the phytopathogenic fungi Stenocarpella maydis and Stenocarpella macrospora. Int. J. Mol. Sci. 2011, 12, 5522–5537. [Google Scholar] [CrossRef] [PubMed]
- Kilani-Feki, O.; Jaoua, S. Biological control of Botrytis cinerea using the antagonistic and endophytic Burkholderia cepacia Cs5 for vine plantlet protection. Can. J. Microbiol. 2011, 57, 896–901. [Google Scholar] [CrossRef] [PubMed]
- Farbo, M.G.; Urgeghe, P.P.; Fiori, S.; Marcello, A.; Oggiano, S.; Balmas, V.; Ul Hassan, Z.; Jaoua, S.; Migheli, Q. Effect of yeast volatile organic compounds on ochratoxin A-producing Aspergillus carbonarius and A. ochraceus. Int. J. Food Microbiol. 2018, 284, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Quan, C.S.; Fan, S.D. Antifungal activity of a novel compound from Burkholderia cepacia against plant pathogenic fungi. Lett. Appl. Microbiol. 2007, 45, 508–514. [Google Scholar] [CrossRef]
- Oliveira, R.R.; Aguiar, B.D.M.; Tessmann, D.J.; Pujade-Renaud, V.; Vida, J.B. Chlamydospore formation by Corynespora cassiicola. Trop. Plant Pathol. 2012, 37, 415–418. [Google Scholar] [CrossRef]
- Chen, N.; Jin, M.; Qu, H.M.; Chen, Z.Q.; Chen, Z.L.; Qiu, Z.G.; Wang, X.W.; Li, J.W. Isolation and characterization of Bacillus sp. producing broad-spectrum antibiotics against human and plant pathogenic fungi. J. Microbiol. Biotechnol. 2012, 22, 256–563. [Google Scholar] [CrossRef]
- Joo, H.J.; Kim, H.Y.; Kim, L.H.; Lee, S.; Ryu, J.G.; Lee, T. A Brevibacillus sp. antagonistic to mycotoxigenic Fusarium spp. Biol. Control 2015, 87, 64–70. [Google Scholar] [CrossRef]
- Kilani-Feki, O.; Zouari, I.; Culioli, G.; Ortalo-Magné, A.; Zouari, N.; Blache, Y.; Jaoua, S. Correlation between synthesis variation of 2-alkylquinolones and the antifungal activity of a Burkholderia cepacia strain collection. World J. Microbiol. Biotechnol. 2012, 28, 275–281. [Google Scholar] [CrossRef]
- Cheba, B.A.; Zaghloul, T.I.; EL-Mahdy, A.R.; EL-Massry, M.H. Effect of pH and temperature on Bacillus sp. R2 chitinase activity and stability. Procedia Technol. 2016, 22, 471–477. [Google Scholar] [CrossRef]
- Muhialdin, B.J.; Hassan, Z.; Saari, N. In vitro antifungal activity of lactic acid bacteria low molecular peptides against spoilage fungi of bakery products. Ann. Microbiol. 2018, 68, 557–567. [Google Scholar] [CrossRef]
- Gomaa, E.Z.; Abdelall, M.F.; El-Mahdy, O.M. Detoxification of aflatoxin B1 by antifungal compounds from Lactobacillus brevis and Lactobacillus paracasei, isolated from dairy products. Probiotics Antimicro. 2018, 10, 201–209. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeidan, R.; Ul-Hassan, Z.; Al-Thani, R.; Migheli, Q.; Jaoua, S. In-Vitro Application of a Qatari Burkholderia cepacia strain (QBC03) in the Biocontrol of Mycotoxigenic Fungi and in the Reduction of Ochratoxin A biosynthesis by Aspergillus carbonarius. Toxins 2019, 11, 700. https://doi.org/10.3390/toxins11120700
Zeidan R, Ul-Hassan Z, Al-Thani R, Migheli Q, Jaoua S. In-Vitro Application of a Qatari Burkholderia cepacia strain (QBC03) in the Biocontrol of Mycotoxigenic Fungi and in the Reduction of Ochratoxin A biosynthesis by Aspergillus carbonarius. Toxins. 2019; 11(12):700. https://doi.org/10.3390/toxins11120700
Chicago/Turabian StyleZeidan, Randa, Zahoor Ul-Hassan, Roda Al-Thani, Quirico Migheli, and Samir Jaoua. 2019. "In-Vitro Application of a Qatari Burkholderia cepacia strain (QBC03) in the Biocontrol of Mycotoxigenic Fungi and in the Reduction of Ochratoxin A biosynthesis by Aspergillus carbonarius" Toxins 11, no. 12: 700. https://doi.org/10.3390/toxins11120700
APA StyleZeidan, R., Ul-Hassan, Z., Al-Thani, R., Migheli, Q., & Jaoua, S. (2019). In-Vitro Application of a Qatari Burkholderia cepacia strain (QBC03) in the Biocontrol of Mycotoxigenic Fungi and in the Reduction of Ochratoxin A biosynthesis by Aspergillus carbonarius. Toxins, 11(12), 700. https://doi.org/10.3390/toxins11120700